
The Antidepressant Sertraline Induces the Formation of Supersized Lipid Droplets in the Human Pathogen Cryptococcus neoformans

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Media, and Drug Treatments
2.2. LD Staining
2.3. Imaging and Post-Acquisition Processing
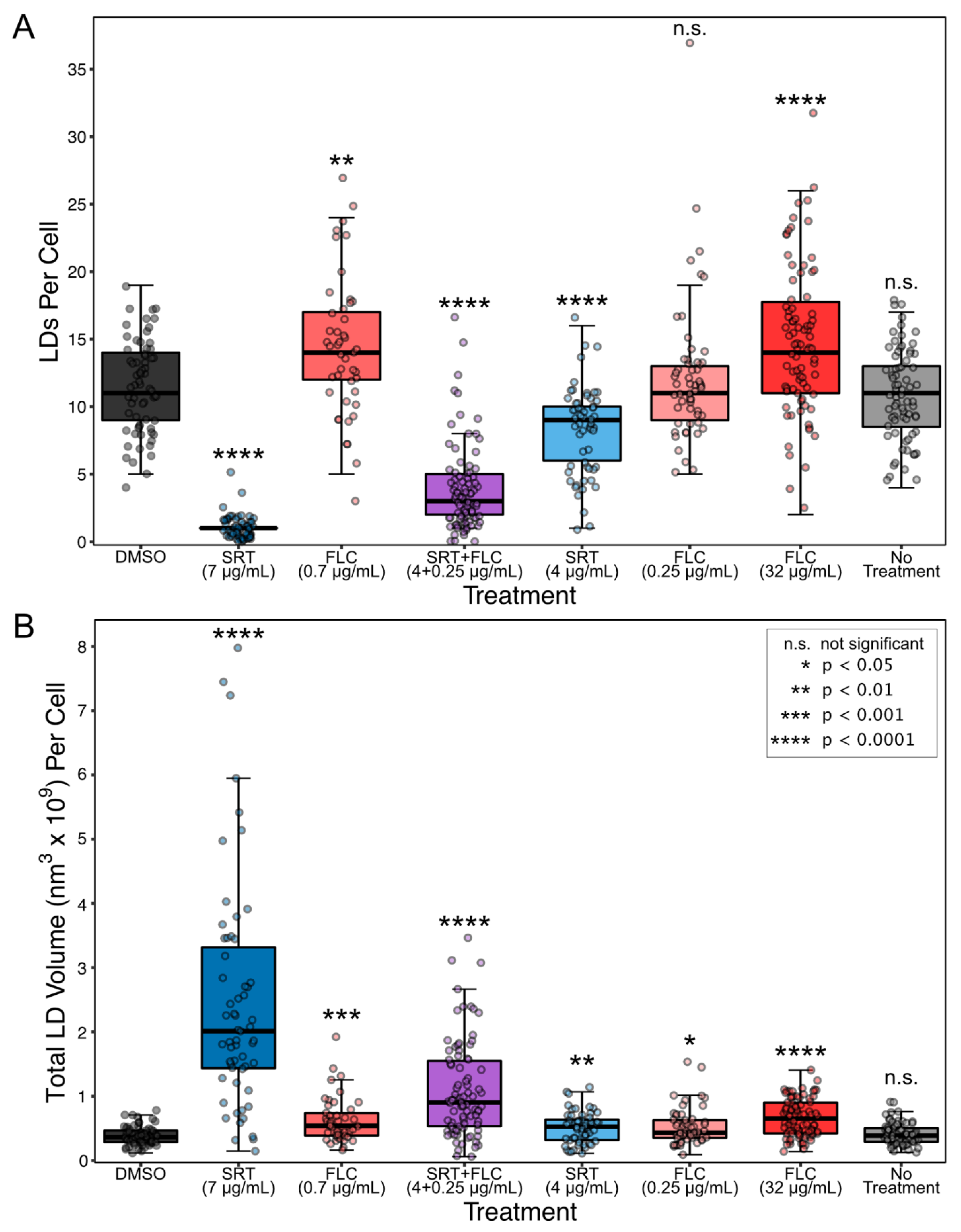
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bongomin, F.; Gago, S.; Oladele, R.O.; Denning, D.W. Global and multi-national prevalence of fungal diseases—estimate precision. J. Fungi 2017, 3, 57. [Google Scholar] [CrossRef]
- Kainz, K.; Bauer, M.A.; Madeo, F.; Carmona-Gutierrez, D. Fungal infections in humans: The silent crisis. Microb. Cell 2020, 7, 143–145. [Google Scholar] [CrossRef]
- Maziarz, E.K.; Perfect, J.R. Cryptococcosis. Infect. Dis. Clin. N. Am. 2016, 30, 179–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Idnurm, A.; Bahn, Y.-S.; Nielsen, K.V.; Lin, X.; Fraser, J.A.; Heitman, J. Deciphering the model pathogenic fungus Cryptococcus neoformans. Nat. Rev. Microbiol. 2005, 3, 753–764. [Google Scholar] [CrossRef] [PubMed]
- Rajasingham, R.; Smith, R.M.; Park, B.J.; Jarvis, J.N.; Govender, N.P.; Chiller, T.M.; Denning, D.W.; Loyse, A.; Boulware, D.R. Global burden of disease of HIV-associated cryptococcal meningitis: An updated analysis. Lancet Infect. Dis. 2017, 17, 873–881. [Google Scholar] [CrossRef] [Green Version]
- Coelho, C.; Casadevall, A. Cryptococcal therapies and drug targets: The old, the new and the promising. Cell. Microbiol. 2016, 18, 792–799. [Google Scholar] [CrossRef] [Green Version]
- Spadari, C.D.C.; Wirth, F.; Lopes, L.B.; Ishida, K. New approaches for Cryptococcosis treatment. Microorganisms 2020, 8, 613. [Google Scholar] [CrossRef]
- Wall, G.; Lopez-Ribot, J.L. Screening repurposing libraries for identification of drugs with novel antifungal activity. Antimicrob. Agents Chemother. 2020, 64, e00924-20. [Google Scholar] [CrossRef] [PubMed]
- Truong, M.; Monahan, L.G.; Carter, D.A.; Charles, I.G. Repurposing drugs to fast-track therapeutic agents for the treatment of cryptococcosis. PeerJ 2018, 6, e4761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McRae, A.L.; Brady, K.T. Review of sertraline and its clinical applications in psychiatric disorders. Expert Opin. Pharmacother. 2001, 2, 883–892. [Google Scholar] [CrossRef]
- Zhai, B.; Wu, C.; Wang, L.; Sachs, M.S.; Lin, X. The antidepressant sertraline provides a promising therapeutic option for neurotropic cryptococcal infections. Antimicrob. Agents Chemother. 2012, 56, 3758–3766. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, S.; Hackett, E.; DuBoff, E.; Greist, J.; Halaris, A.; Koran, L.M.; Liebowitz, M.; Lydiard, R.B.; McElroy, S.; Mendels, J.; et al. A 2-year study of sertraline in the treatment of obsessive-compulsive disorder. Int. Clin. Psychopharmacol. 1997, 12, 309–316. [Google Scholar] [CrossRef]
- Cook, E.H.; Wagner, K.D.; March, J.S.; Biederman, J.; Landau, P.; Wolkow, R.; Messig, M. Long-term sertraline treatment of children and adolescents with obsessive-compulsive disorder. J. Am. Acad. Child Adolesc. Psychiatry 2001, 40, 1175–1181. [Google Scholar] [CrossRef] [PubMed]
- Rapaport, M.H.; Wolkow, R.; Rubin, A.; Hackett, E.; Pollack, M.; Ota, K.Y. Sertraline treatment of panic disorder: Results of a long-term study. Acta Psychiatr. Scand. 2001, 104, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Treviño-Rangel, R.D.J.; Villanueva-Lozano, H.; Hernández-Rodríguez, P.; Martínez-Reséndez, M.F.; García-Juárez, J.; Rodríguez-Rocha, H.; González, G.M. Activity of sertraline against Cryptococcus neoformans: In vitro and in vivo assays. Med. Mycol. 2016, 54, 280–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tremaine, L.M.; Welch, W.M.; Ronfeld, R.A. Metabolism and disposition of the 5-hydroxytryptamine uptake blocker sertraline in the rat and dog. Drug Metab. Dispos. 1989, 17, 542–550. [Google Scholar] [PubMed]
- Nayak, R.; Xu, J. Effects of sertraline hydrochloride and fluconazole combinations on Cryptococcus neoformans and Cryptococcus gattii. Mycology 2010, 1, 99–105. [Google Scholar] [CrossRef]
- Rossato, L.; Loreto, E.S.; Zanette, R.A.; Chassot, F.; Santurio, J.M.; Alves, S.H. In vitro synergistic effects of chlorpromazine and sertraline in combination with amphotericin B against Cryptococcus neoformans var. grubii. Folia Microbiol. 2016, 61, 399–403. [Google Scholar] [CrossRef]
- Brown, J.C.S.; Nelson, J.; VanderSluis, B.; Deshpande, R.; Butts, A.; Kagan, S.; Polacheck, I.; Krysan, D.J.; Myers, C.L.; Madhani, H.D. Unraveling the biology of a fungal meningitis pathogen using chemical genetics. Cell 2014, 159, 1168–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuller, R.W.; Hemrick-Luecke, S.K.; Littlefield, E.S.; Audia, J.E. Comparison of desmethylsertraline with sertraline as a monoamine uptake inhibitor in vivo. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 1995, 19, 135–149. [Google Scholar] [CrossRef]
- Nedahl, M.; Johansen, S.S.; Linnet, K. Reference brain/blood concentrations of citalopram, duloxetine, mirtazapine and sertraline. J. Anal. Toxicol. 2018, 42, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Wiener, H.L.; Kramer, H.K.; Reith, M.E. Separation and determination of sertraline and its metabolite, desmethylsertraline, in mouse cerebral cortex by reversed-phase high-performance liquid chromatography. J. Chromatogr. 1990, 527, 467–472. [Google Scholar] [CrossRef]
- Boulware, D.R.; Nalintya, E.; Rajasingham, R.; Kirumira, P.; Naluyima, R.; Turya, F.; Namanda, S.; Rutakingirwa, M.K.; Skipper, C.P.; Nikweri, Y.; et al. Adjunctive sertraline for asymptomatic cryptococcal antigenemia: A randomized clinical trial. Med. Mycol. 2020, 58, 1037–1043. [Google Scholar] [CrossRef]
- Rhein, J.; Morawski, B.M.; Hullsiek, K.H.; Nabeta, H.W.; Kiggundu, R.; Tugume, L.; Musubire, A.; Akampurira, A.; Smith, K.D.; Alhadab, A.; et al. Efficacy of adjunctive sertraline for the treatment of HIV-associated cryptococcal meningitis: An open-label dose-ranging study. Lancet Infect. Dis. 2016, 16, 809–818. [Google Scholar] [CrossRef] [Green Version]
- Alhadab, A.A.; ASTRO-CM Study Team; Rhein, J.; Tugume, L.; Musubire, A.; Williams, D.A.; Abassi, M.; Nicol, M.R.; Meya, D.B.; Boulware, D.R.; et al. Pharmacokinetics-pharmacodynamics of sertraline as an antifungal in HIV-infected Ugandans with cryptococcal meningitis. J. Pharmacokinet. Pharmacodyn. 2019, 46, 565–576. [Google Scholar] [CrossRef]
- Rainey, M.M.; Korostyshevsky, D.; Lee, S.; Perlstein, E.O. The antidepressant sertraline targets intracellular vesiculogenic membranes in yeast. Genetics 2010, 185, 1221–1233. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Korostyshevsky, D.; Lee, S.; Perlstein, E.O. Accumulation of an antidepressant in vesiculogenic membranes of yeast cells triggers autophagy. PLoS ONE 2012, 7, e34024. [Google Scholar] [CrossRef] [Green Version]
- Olzmann, J.A.; Carvalho, P. Dynamics and functions of lipid droplets. Nat. Rev. Mol. Cell Biol. 2019, 20, 137–155. [Google Scholar] [CrossRef]
- Henne, W.M.; Reese, M.L.; Goodman, J.M. The assembly of lipid droplets and their roles in challenged cells. EMBO J. 2018, 37, e98947. [Google Scholar] [CrossRef]
- Kuo, K.; Hoch, H. Germination of Phyllosticta ampelicida pycnidiospores: Prerequisite of adhesion to the substratum and the relationship of substratum wettability. Fungal Genet. Biol. 1996, 20, 18–29. [Google Scholar] [CrossRef]
- Gocze, P.M.; Freeman, D.A. Factors underlying the variability of lipid droplet fluorescence in MA-10 leydig tumor cells. Cytometry 1994, 17, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.-J.; Hsu, C.-L.; Yang, J.-Y.; Yang, W.Y. Monodansylpentane as a blue-fluorescent lipid-droplet marker for multi-color live-cell imaging. PLoS ONE 2012, 7, e32693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azadmanesh, J.; Gowen, A.M.; Creger, P.E.; Schafer, N.D.; Blankenship, J.R. Filamentation involves two overlapping, but distinct, programs of filamentation in the pathogenic fungus Candida albicans. G3 Genes Genomes Genet. 2017, 7, 3797–3808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fei, W.; Shui, G.; Zhang, Y.; Krahmer, N.; Ferguson, C.; Kapterian, T.S.; Lin, R.C.; Dawes, I.W.; Brown, A.J.; Li, P.; et al. A role for phosphatidic acid in the formation of “supersized” lipid droplets. PLoS Genet. 2011, 7, e1002201. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.-W. Lipid droplet dynamics in budding yeast. Cell. Mol. Life Sci. 2015, 72, 2677–2695. [Google Scholar] [CrossRef]
- Krahmer, N.; Guo, Y.; Wilfling, F.; Hilger, M.; Lingrell, S.; Heger, K.; Newman, H.W.; Schmidt-Supprian, M.; Vance, D.E.; Mann, M.; et al. Phosphatidylcholine synthesis for lipid droplet expansion is mediated by localized activation of CTP: Phosphocholine cytidylyltransferase. Cell Metab. 2011, 14, 504–515. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.; Zhang, M.; Zheng, S.; Li, Y.; Li, X.; Li, W.; Li, G.; Lin, Z.; Xie, Z.; Zhao, Z.; et al. Trapping toxins within lipid droplets is a resistance mechanism in fungi. Sci. Rep. 2015, 5, 15133. [Google Scholar] [CrossRef] [Green Version]
- Hanano, A.; Alkara, M.; Almousally, I.; Shaban, M.; Rahman, F.; Hassan, M.; Murphy, D.J. The peroxygenase activity of the Aspergillus flavus caleosin, AfPXG, modulates the biosynthesis of aflatoxins and their trafficking and extracellular secretion via lipid droplets. Front. Microbiol. 2018, 9, 158. [Google Scholar] [CrossRef]
- Deak, K.; Takacs-Novak, K.; Tihanyi, K.; Noszal, B. Physico-chemical profiling of antidepressive sertraline: Solubility, ionisation, lipophilicity. Med. Chem. 2006, 2, 385–389. [Google Scholar] [CrossRef]
- Li, W.; Yun, Z.; Ji, C.; Tu, J.; Yang, W.; Li, J.; Liu, N.; Sheng, C. Discovery of novel sertraline derivatives as potent anti-Cryptococcus agents. J. Med. Chem. 2022, 65, 6541–6554. [Google Scholar] [CrossRef]
- Fei, W.; Shui, G.; Gaeta, B.; Du, X.; Kuerschner, L.; Li, P.; Brown, A.J.; Wenk, M.R.; Parton, R.G.; Yang, H. Fld1p, a functional homologue of human seipin, regulates the size of lipid droplets in yeast. J. Cell Biol. 2008, 180, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Nolan, S.J.; Fu, M.S.; Coppens, I.; Casadevall, A. Lipids affect the Cryptococcus neoformans-macrophage interaction and promote nonlytic exocytosis. Infect. Immun. 2017, 85, e00564-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Breuer, M.R.; Dasgupta, A.; Vasselli, J.G.; Lin, X.; Shaw, B.D.; Sachs, M.S. The Antidepressant Sertraline Induces the Formation of Supersized Lipid Droplets in the Human Pathogen Cryptococcus neoformans. J. Fungi 2022, 8, 642. https://doi.org/10.3390/jof8060642
Breuer MR, Dasgupta A, Vasselli JG, Lin X, Shaw BD, Sachs MS. The Antidepressant Sertraline Induces the Formation of Supersized Lipid Droplets in the Human Pathogen Cryptococcus neoformans. Journal of Fungi. 2022; 8(6):642. https://doi.org/10.3390/jof8060642
Chicago/Turabian StyleBreuer, Matthew R., Ananya Dasgupta, Joseph G. Vasselli, Xiaorong Lin, Brian D. Shaw, and Matthew S. Sachs. 2022. "The Antidepressant Sertraline Induces the Formation of Supersized Lipid Droplets in the Human Pathogen Cryptococcus neoformans" Journal of Fungi 8, no. 6: 642. https://doi.org/10.3390/jof8060642