
Lack of Association between Fluconazole Susceptibility and ERG11 Nucleotide Polymorphisms in Cryptococcus neoformans Clinical Isolates from Uganda

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fluconazole Minimum Inhibitory Concentration (MIC) Broth Microdilution Assays
2.2. DNA Extraction, Amplification, and Sequencing
2.3. Analysis
3. Results
3.1. In Vitro Fluconazole Susceptibility
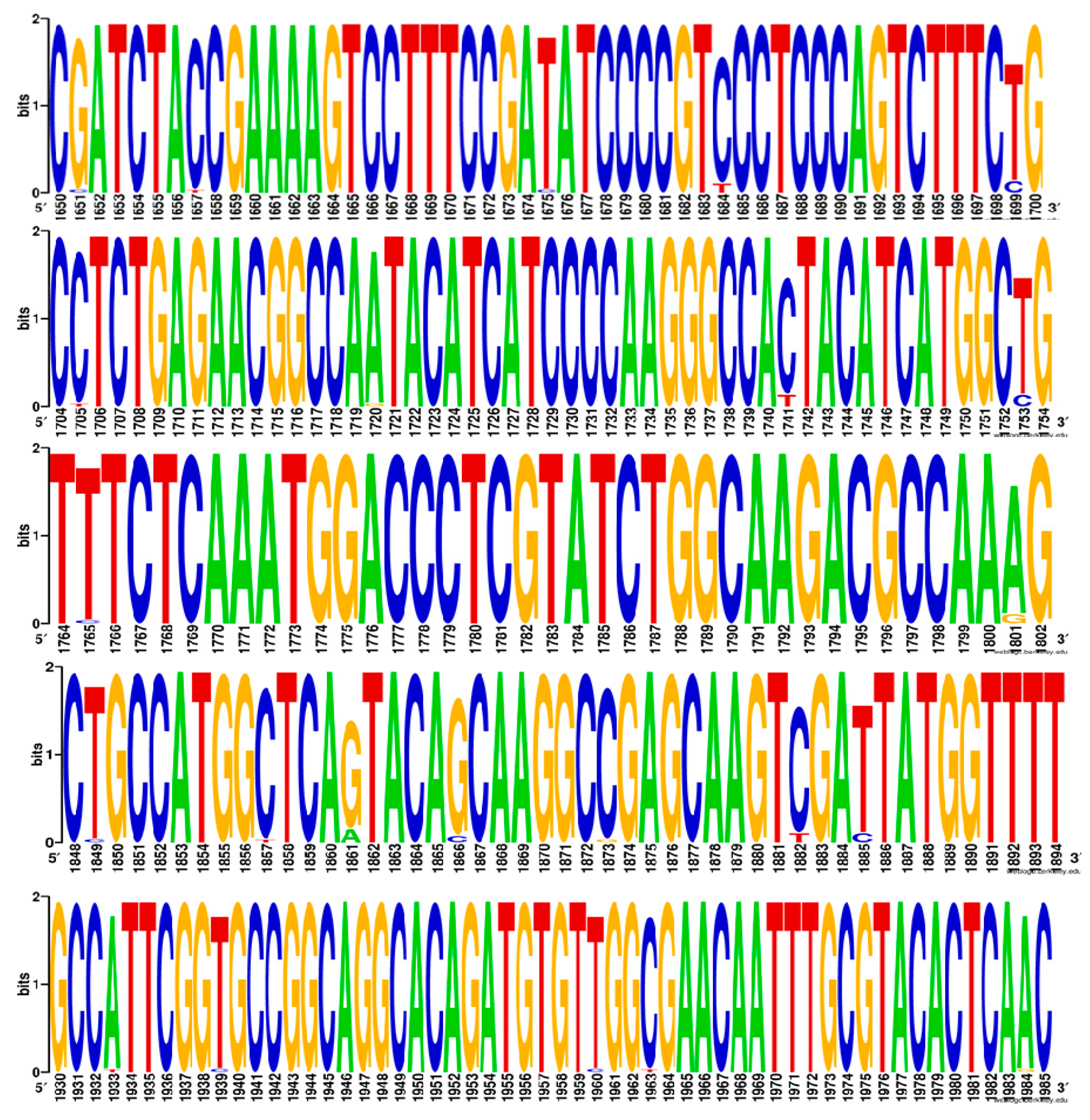
3.2. SNP Analysis of the ERG11 Gene
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parkes-Ratanshi, R.; Wakeham, K.; Levin, J.; Namusoke, D.; Whitworth, J.; Coutinho, A.; Mugisha, N.K.; Grosskurth, H.; Kamali, A.; Lalloo, D.G. Primary prophylaxis of cryptococcal disease with fluconazole in HIV-positive Ugandan adults: A double-blind, randomised, placebo-controlled trial. Lancet Infect. Dis. 2011, 11, 933–941. [Google Scholar] [CrossRef] [Green Version]
- Perfect, J.R.; Dismukes, W.E.; Dromer, F.; Goldman, D.L.; Graybill, J.R.; Hamill, R.J.; Harrison, T.S.; Larsen, R.A.; Lortholary, O.; Nguyen, M.-H.; et al. Clinical practice guidelines for the management of cryptococcal disease: 2010 update by the infectious diseases society of america. Clin. Infect. Dis. 2010, 50, 291–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loyse, A.; Burry, J.; Cohn, J.; Ford, N.; Chiller, T.; Ribeiro, I.; Koulla-Shiro, S.; Mghamba, J.; Ramadhani, A.; Nyirenda, R.; et al. Leave no one behind: Response to new evidence and guidelines for the management of cryptococcal meningitis in low-income and middle-income countries. Lancet Infect. Dis. 2019, 19, 143–147. [Google Scholar] [CrossRef]
- Hope, W.; Stone, N.R.H.; Johnson, A.; McEntee, L.; Farrington, N.; Santoro-Castelazo, A.; Liu, X.; Lucaci, A.; Hughes, M.; Oliver, J.D.; et al. Fluconazole monotherapy is a suboptimal option for initial treatment of cryptococcal meningitis because of emergence of resistance. MBio 2019, 10, e02575-19. Available online: https://www.meta.org/papers/fluconazole-monotherapy-is-a-suboptimal-option/31796539 (accessed on 8 February 2021). [CrossRef] [PubMed] [Green Version]
- Parkes-Ratanshi, R.; Achan, B.; Kwizera, R.; Kambugu, A.; Meya, D.; Denning, D.W. Cryptococcal disease and the burden of other fungal diseases in Uganda; Where are the knowledge gaps and how can we fill them? Mycoses 2015, 58, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Abassi, M.; Rhein, J.; Meya, D.B.; Boulware, D.R. Cryptococcal disease in the era of “test and treat”: Is there cause for concern? Open Forum Infect. Dis. 2018, 5, ofx274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williamson, P.R. The relentless march of cryptococcal meningitis. Lancet Infect. Dis. 2017, 8, 790–791. [Google Scholar] [CrossRef]
- Smith, K.D.; Achan, B.; Hullsiek, K.H.; McDonald, T.R.; Okagaki, L.H.; Alhadab, A.A.; Akampurira, A.; Rhein, J.R.; Meya, D.B.; Boulware, D.R.; et al. Increased antifungal drug resistance in clinical isolates of Cryptococcus neoformans in Uganda. Antimicrob. Agents Chemother. 2015, 59, 7197–7204. [Google Scholar] [CrossRef] [Green Version]
- Naicker, S.D.; Mpembe, R.S.; Maphanga, T.G.; Zulu, T.G.; Desanto, D.; Wadula, J.; Mvelase, N.; Maluleka, C.; Reddy, K.; Dawood, H.; et al. Decreasing fluconazole susceptibility of clinical south african Cryptococcus neoformans isolates over a decade. PLoS Negl. Trop. Dis. 2020, 14, e0008137. [Google Scholar] [CrossRef] [Green Version]
- Cheong, J.W.S.; Mccormack, J. Fluconazole resistance in cryptococcal disease: Emerging or intrinsic? Med. Mycol. 2013, 51, 261–269. [Google Scholar] [CrossRef] [Green Version]
- Gerlach, E.S.; Altamirano, S.; Yoder, J.M.; Luggya, T.S.; Akampurira, A.; Meya, D.B.; Boulware, D.R.; Rhein, J.; Nielsen, K. ATI-2307 exhibits equivalent antifungal activity in Cryptococcus neoformans clinical isolates with High and low Fluconazole IC50. Front. Cell. Infect. Microbiol. 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Goa, K.L.; Barradell, L.B. Fluconazole: An update of its pharmacodynamic and pharmacokinetic properties and therapeutic use in major superficial and systemic mycoses in immunocompromised patients. Drugs 1995, 50, 658–690. [Google Scholar] [CrossRef] [PubMed]
- Sykes, J.E.; Hodge, G.; Singapuri, A.; Yang, M.L.; Gelli, A.; Thompson, G.R. In vivo development of fluconazole resistance in serial Cryptococcus gattii isolates from a cat. Med. Mycol. 2017, 55, 396–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sionov, E.; Lee, H.; Chang, Y.C.; Kwon-Chung, K.J. Cryptococcus neoformans overcomes stress of azole drugs by formation of disomy in specific multiple chromosomes. PLoS Pathog. 2010, 6, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sionov, E.; Chang, Y.C.; Garraffo, H.M.; Kwon-Chung, K.J. Heteroresistance to fluconazole in Cryptococcus neoformans is intrinsic and associated with virulence. Antimicrob. Agents Chemother. 2009, 53, 2804–2815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basso, L.R.; Gast, C.E.; Bruzual, I.; Wong, B. Identification and properties of plasma membrane azole efflux pumps from the pathogenic fungi Cryptococcus gattii and Cryptococcus neoformans. J. Antimicrob. Chemother. 2015, 70, 1396–1407. [Google Scholar] [CrossRef] [Green Version]
- Kano, R.; Okubo, M.; Hasegawa, A.; Kamata, H. Multi-azole-resistant strains of Cryptococcus neoformans var. grubii isolated from a FLZ-resistant strain by culturing in medium containing voriconazole. Med. Mycol. 2017, 55, 877–882. [Google Scholar] [CrossRef]
- Sionov, E.; Chang, Y.C.; Garraffo, H.M.; Dolan, M.A.; Ghannoum, M.A.; Kwon-Chung, K.J. Identification of a Cryptococcus neoformans Cytochrome P450 Lanosterol 14 -Demethylase (Erg11) residue critical for differential susceptibility between Fluconazole/Voriconazole and Itraconazole/Posaconazole. Antimicrob. Agents Chemother. 2011, 56, 1162–1169. [Google Scholar] [CrossRef] [Green Version]
- Selb, R.; Fuchs, V.; Graf, B.; Hamprecht, A.; Hogardt, M.; Sedlacek, L.; Schwarz, R.; Idelevich, E.A.; Becker, S.L.; Held, J.; et al. Molecular typing and in vitro resistance of Cryptococcus neoformans clinical isolates obtained in Germany between 2011 and 2017. Int. J. Med. Microbiol. 2019, 309, 151336. [Google Scholar] [CrossRef]
- Rodero, L.; Mellado, E.; Rodriguez, A.C.; Salve, A.; Guelfand, L.; Cahn, P.; Cuenca-Estrella, M.; Davel, G.; Rodriguez-Tudela, J.L. G484S amino acid substitution in lanosterol 14-alpha demethylase (ERG11) is related to fluconazole resistance in a recurrent Cryptococcus neoformans clinical isolate. Antimicrob. Agents Chemother. 2003, 47, 3653–3656. [Google Scholar] [CrossRef] [Green Version]
- Carneiro, H.C.S.; Bastos, R.W.; Ribeiro, N.Q.; Gouveia-Eufrasio, L.; Costa, M.C.; Magalhães, T.F.F.; Oliveira, L.V.N.; Paixão, T.A.; Joffe, L.S.; Rodrigues, M.L.; et al. Hypervirulence and cross-resistance to a clinical antifungal are induced by an environmental fungicide in Cryptococcus gattii. Sci. Total Environ. 2020, 740, 140135. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.; Bangdiwala, A.S.; Cresswell, F.V.; Rhein, J.; Nuwagira, E.; Ssebambulidde, K.; Tugume, L.; Rajasingham, R.; Bridge, S.C.; Muzoora, C.; et al. The changing epidemiology of HIV-Associated Adult Meningitis, Uganda 2015–2017. Open Forum Infect. Dis. 2019, 6, ofz419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhein, J.; Morawski, B.M.; Hullsiek, K.H.; Nabeta, H.W.; Kiggundu, R.; Tugume, L.; Musubire, A.; Akampurira, A.; Smith, K.D.; Alhadab, A.; et al. Efficacy of adjunctive sertraline for the treatment of HIV-associated cryptococcal meningitis: An open-label dose-ranging study. Lancet. Infect. Dis. 2016, 16, 809–818. [Google Scholar] [CrossRef] [Green Version]
- Rhein, J.; Hullsiek, K.H.; Evans, E.E.; Tugume, L.; Nuwagira, E.; Ssebambulidde, K.; Kiggundu, R.; Mpoza, E.; Musubire, A.K.; Bangdiwala, A.S.; et al. Detrimental outcomes of unmasking Cryptococcal Meningitis with recent ART initiation. Open Forum Infect. Dis. 2018, 5, ofy122. [Google Scholar] [CrossRef] [PubMed]
- Arendrup, M.C.; Meletiadis, J.; Mouton, J.W.; Lagrou, K.; Hamal, P.; Guinea, J. Method for the determination of broth dilution minimum inhibitory concentrations of antifungal agents for yeasts. EUCAST definitive document E. Def 2017, 7, E246–E247. [Google Scholar]
- Nielsen, K.; Cox, G.M.; Wang, P.; Toffaletti, D.L.; Perfect, J.R.; Heitman, J. Sexual cycle of Cryptococcus neoformans var. grubii and Virulence of congenic a and α isolates. Infect. Immun. 2003, 71, 4831–4841. [Google Scholar] [CrossRef] [Green Version]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coordinators, N.R. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2016, 44, D7–D19. [Google Scholar] [CrossRef] [Green Version]
- Crooks, G.E.; Hon, G.; Chandonia, J.-M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [Green Version]
- Stanke, M.; Morgenstern, B. Augustus: A web server for gene prediction in eukaryotes that allows user-defined constraints. Nucleic Acids Res. 2005, 33, W465–W467. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Muscle: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bongomin, F.; Oladele, R.O.; Gago, S.; Moore, C.B.; Richardson, M.D. A systematic review of fluconazole resistance in clinical isolates of Cryptococcus species. Mycoses 2018, 61, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Chesdachai, S.; Rajasingham, R.; Nicol, M.R.; Meya, D.B.; Bongomin, F.; Abassi, M.; Skipper, C.; Kwizera, R.; Rhein, J.; Boulware, D.R. Minimum inhibitory concentration distribution of Fluconazole against Cryptococcus species and the Fluconazole exposure prediction model. Open Forum Infect. Dis. 2019, 6, ofz369. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A.; Diekema, D.J.; Gibbs, D.L.; Newell, V.A.; Bijie, H.; Dzierzanowska, D.; Klimko, N.N.; Letscher-Bru, V.; Lisalova, M.; Muehlethaler, K.; et al. Results from the ARTEMIS DISK Global Antifungal Surveillance Study, 1997 to 2007: 10.5-year analysis of susceptibilities of noncandidal yeast species to fluconazole and voriconazole determined by CLSI standardized disk diffusion testing. J. Clin. Microbiol. 2009, 47, 117–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aller, A.I.; Martin-Mazuelos, E.; Lozano, F.; Gomez-Mateos, J.; Steele-Moore, L.; Holloway, W.J.; Gutiérrez, M.J.; Recio, F.J.; Espinel-Ingroff, A. Correlation of fluconazole MICs with clinical outcome in cryptococcal infection. Antimicrob. Agents Chemother. 2000, 44, 1544–1548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berman, J.; Krysan, D.J. Drug resistance and tolerance in fungi. Nat. Rev. Microbiol. 2020, 18, 319–331. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Oliveira, N.K.; Savitt, A.G.; Silva, V.K.A.; Krausert, R.B.; Ghebrehiwet, B.; Fries, B.C. Low glucose mediated Fluconazole tolerance in Cryptococcus neoformans. J. Fungi 2021, 7, 489. [Google Scholar] [CrossRef] [PubMed]
- Carlson, T.; Lupinacci, E.; Moseley, K.; Chandrasekaran, S. Effects of environmental factors on sensitivity of Cryptococcus neoformans to fluconazole and amphotericin B. FEMS Microbiol. Lett. 2021, 368, fnab040. [Google Scholar] [CrossRef] [PubMed]
- Castelnuovo, B.; Manabe, Y.C.; Kiragga, A.; Kamya, M.; Easterbrook, P.; Kambugu, A. Cause-Specific mortality and the contribution of immune reconstitution inflammatory Syndrome in the first 3 years after Antiretroviral therapy initiation in an urban african cohort. Clin. Infect. Dis. 2009, 49, 965–972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceita, G.D.O.; Ceita, G.D.O.; Vilas-Boas, L.A.; Castilho, M.S.; Carazzolle, M.F.; Pirovani, C.P.; Selbach-Schnadelbach, A.; Gramacho, K.P.; Ramos, P.I.P.; Barbosa, L.V.; et al. Analysis of the ergosterol biosynthesis pathway cloning, Molecular characterization and phylogeny of lanosterol 14 α-demethylase (ERG11) gene of moniliophthora perniciosa. Genet. Mol. Biol. 2014, 37, 683–693. [Google Scholar] [CrossRef]
- Dos Santos Silva, D.B.; Carbonera Rodrigues, L.M.; De Almeida, A.A.; de Oliveira, K.M.P.; Grisolia, A.B. Novel point mutations in the ERG11 gene in clinical isolates of azole resistant Candida species. Mem. Inst. Oswaldo Cruz 2016, 111, 192–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Onyewu, C.; Yoell, H.J.; Ali, R.Y.; Vilgalys, R.J.; Mitchell, T.G. Dynamic and heterogeneous mutations to fluconazole resistance in Cryptococcus neoformans. Antimicrob. Agents Chemother. 2001, 45, 420–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, S.; Dadwal, R.; Singh, S.; Shaw, D.; Chakrabarti, A.; Rudramurthy, S.M.; Ghosh, A.K. Rapid detection of ERG11 polymorphism associated azole resistance in Candida tropicalis. PLoS ONE 2021, 16, e0245160. [Google Scholar] [CrossRef] [PubMed]
- Bosco-Borgeat, M.E.; Mazza, M.; Taverna, C.G.; Córdoba, S.; Murisengo, O.A.; Vivot, W.; Davel, G. Amino acid substitution in Cryptococcus neoformans lanosterol 14-α-demethylase involved in fluconazole resistance in clinical isolates. Rev. Argent. Microbiol. 2016, 48, 137–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akapo, O.O.; Macnar, J.M.; Kryś, J.D.; Syed, P.R.; Syed, K.; Gront, D. In silico structural modeling and analysis of interactions of tremellomycetes cytochrome p450 monooxygenases cyp51s with substrates and azoles. Int. J. Mol. Sci. 2021, 22, 7811. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Low IC50 | High IC50 | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Fluconazole IC50 (μg/mL) | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | 64 |
| N [proportion] | 0 (0) | 7 (2.2) | 16 (5.16) | 44 (14.2) | 107 (34.5) | 93 (30) | 39 (12.6) | 0 (0) | 4 (1.29) |
| Parameter | Results n = 37 |
|---|---|
| Age in years: median (range) | 35.9 (20–65) |
| Gender | |
| Male | 21 (56.7%) |
| Female | 16 (43.2%) |
| CM Relapse | |
| History of CM | 3 (8.1%) |
| No History of CM | 34 (91.9%) |
| Mortality | |
| Alive | 28 (75.7%) |
| Dead | 9 (24.3%) |
| Isolate | SNP Type | SNP | IC50 (μg/mL) | GenBank Codes |
|---|---|---|---|---|
| 110159 | A1861G | Synonymous | 8 | MZ673063 |
| 110166 | A1861G | Synonymous | 8 | MZ673065 |
| 110174 | A1861G | Synonymous | 16 | MZ673067 |
| 110180 | C1741T | Synonymous | 8 | MZ673055 |
| 110183 | A1861G | Synonymous | 2 | MZ673066 |
| 110242 | A1861G | Synonymous | 4 | MZ673077 |
| 110246 | A1861G | Synonymous | 8 | MZ673064 |
| 110252 | A1861G | Synonymous | 0.5 | MZ673070 |
| 110271 | A1861G | Synonymous | 2 | MZ673058 |
| 110288 | C1741T, A1861G | Synonymous | 2 | MZ673080 |
| 110290 | C1741T, A1861G | Synonymous | 4 | MZ673084 |
| 110301 | A1861G | Synonymous | 8 | MZ673071 |
| 110352 | A1861G | Synonymous | 4 | MZ673060 |
| 110353 | A1861G | Synonymous | 4 | MZ673083 |
| 110355 | A1861G | Synonymous | 8 | MZ673074 |
| 110389 | A1861G | Synonymous | 8 | MZ673059 |
| 110390 | A1861G | Synonymous | 2 | MZ673052 |
| 110395 | A1861G | Synonymous | 2 | MZ673078 |
| 110399 | C1684T, T1699C, T1753C, A1801G, C1882T, ‡ G1866C | Missense mutation S460T 1 | 4 | MZ673089 |
| 110404 | A1861G | Synonymous | 4 | MZ673054 |
| 110413 | A1861G | Synonymous | 8 | MZ673081 |
| 110414 D1 | C1684T, T1699C, T1753C, C1882T, T1885C, ‡ G1866C | Missense mutation S460T 1 | 2 | MZ673086 |
| 110414 D3 | C1684T, T1699C, T1753C, A1801G, C1882T, T1885C, | Synonymous | 2 | MZ673087 |
| 110416 | A1861G | Synonymous | 8 | MZ673056 |
| 110418 | A1861G | Synonymous | 4 | MZ673079 |
| 110420 | G1651C, C1657T, T1675C, T1699C, C1705T, A1720G, C1741T, T1753C, T1765C, T1768Y, A1801G, T1849C, ‡ C1857T, C1873G, T1903C, A1933T, T1939C, T1960C, C1963T, A1984C | Missense mutation A457V 1 | 4 | MZ673090 |
| 110422 | A1861G | Synonymous | 4 | MZ673068 |
| 110428 | - | Identical to reference wild-type C. neoforman sequence AY265353.1 | 4 | MZ673075 |
| 110429 | A1861G | Synonymous | 4 | MZ673085 |
| 110433 | A1861G | Synonymous | 8 | MZ673076 |
| 110433 * | A1861G | Synonymous | 4 | MZ673057 |
| 110435 | A1861G | Synonymous | 4 | MZ673073 |
| 110439 | A1861G | Synonymous | 4 | MZ673082 |
| 110441 | A1861G | Synonymous | 4 | MZ673062 |
| 110444 D1 | A1861G | Synonymous | 4 | MZ673061 |
| 110444 D7 | A1861G | Synonymous | 8 | MZ673051 |
| 110449 | - | Identical to reference wild-type C.neoforman sequence AY265353.1 | 1 | MZ673088 |
| 110450 | A1861G | Synonymous | 8 | MZ673053 |
| 110451 | A1861G | Synonymous | 4 | MZ673072 |
| 110461 | A1861G | Synonymous | 8 | MZ673069 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atim, P.B.; Meya, D.B.; Gerlach, E.S.; Muhanguzi, D.; Male, A.; Kanamwanji, B.; Nielsen, K. Lack of Association between Fluconazole Susceptibility and ERG11 Nucleotide Polymorphisms in Cryptococcus neoformans Clinical Isolates from Uganda. J. Fungi 2022, 8, 508. https://doi.org/10.3390/jof8050508
Atim PB, Meya DB, Gerlach ES, Muhanguzi D, Male A, Kanamwanji B, Nielsen K. Lack of Association between Fluconazole Susceptibility and ERG11 Nucleotide Polymorphisms in Cryptococcus neoformans Clinical Isolates from Uganda. Journal of Fungi. 2022; 8(5):508. https://doi.org/10.3390/jof8050508
Chicago/Turabian StyleAtim, Priscilla Belbir, David B. Meya, Elliot S. Gerlach, Dennis Muhanguzi, Allan Male, Benedict Kanamwanji, and Kirsten Nielsen. 2022. "Lack of Association between Fluconazole Susceptibility and ERG11 Nucleotide Polymorphisms in Cryptococcus neoformans Clinical Isolates from Uganda" Journal of Fungi 8, no. 5: 508. https://doi.org/10.3390/jof8050508