
Mechanism of Wheat Leaf Rust Control Using Chitosan Nanoparticles and Salicylic Acid

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cultivation of Wheat Plants
2.2. Morphogenesis of the Disease on the Susceptible Variety
2.2.1. Effect of Chitosan Nanoparticles and Salicylic Acid Treatments on Puccinia triticina Spores
2.2.2. Latent and Incubation Periods and Number of Pustules
2.2.3. Measurement of Pustule Size
2.3. Biochemical Assays of Antioxidant Enzymes
2.4. Histochemical Analysis of Reactive Oxygen Species (ROS)
2.5. Anatomical Studies
2.6. Defense-Related Genes Transcriptional Levels
2.7. Statistical Analysis
3. Results
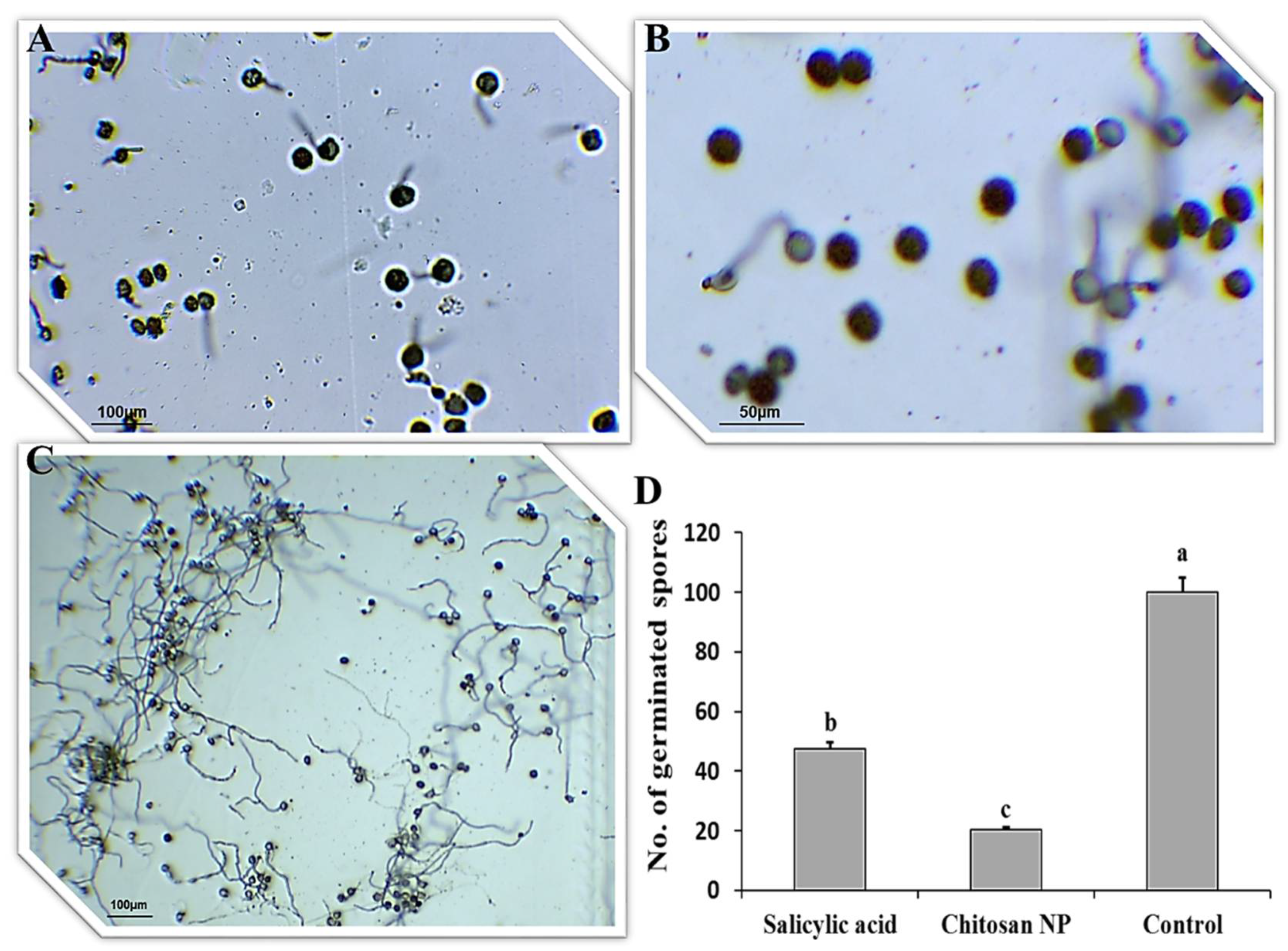
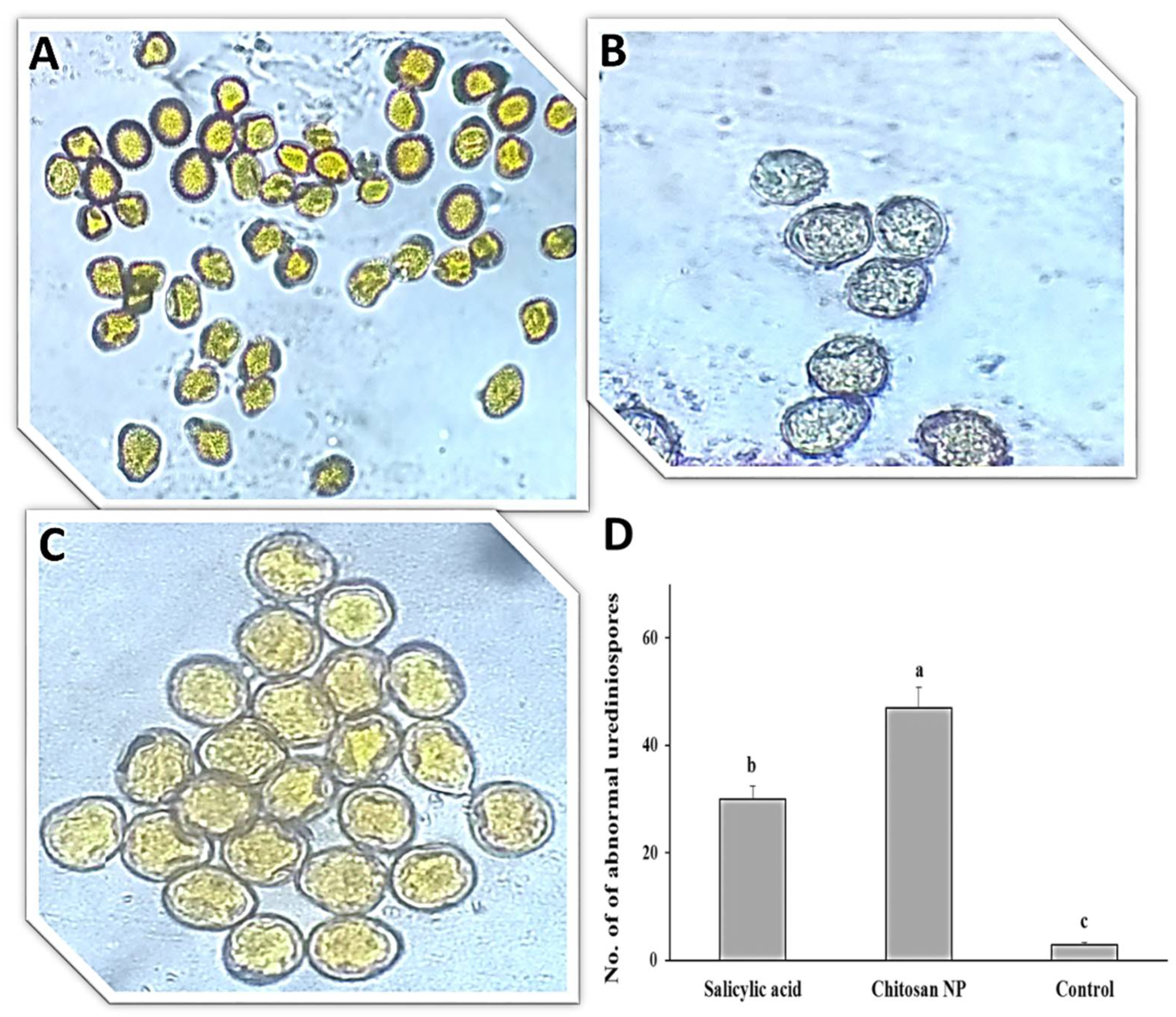
3.1. Effect of Chitosan Nanoparticles and Salicylic Acid on Urediniospores Germination
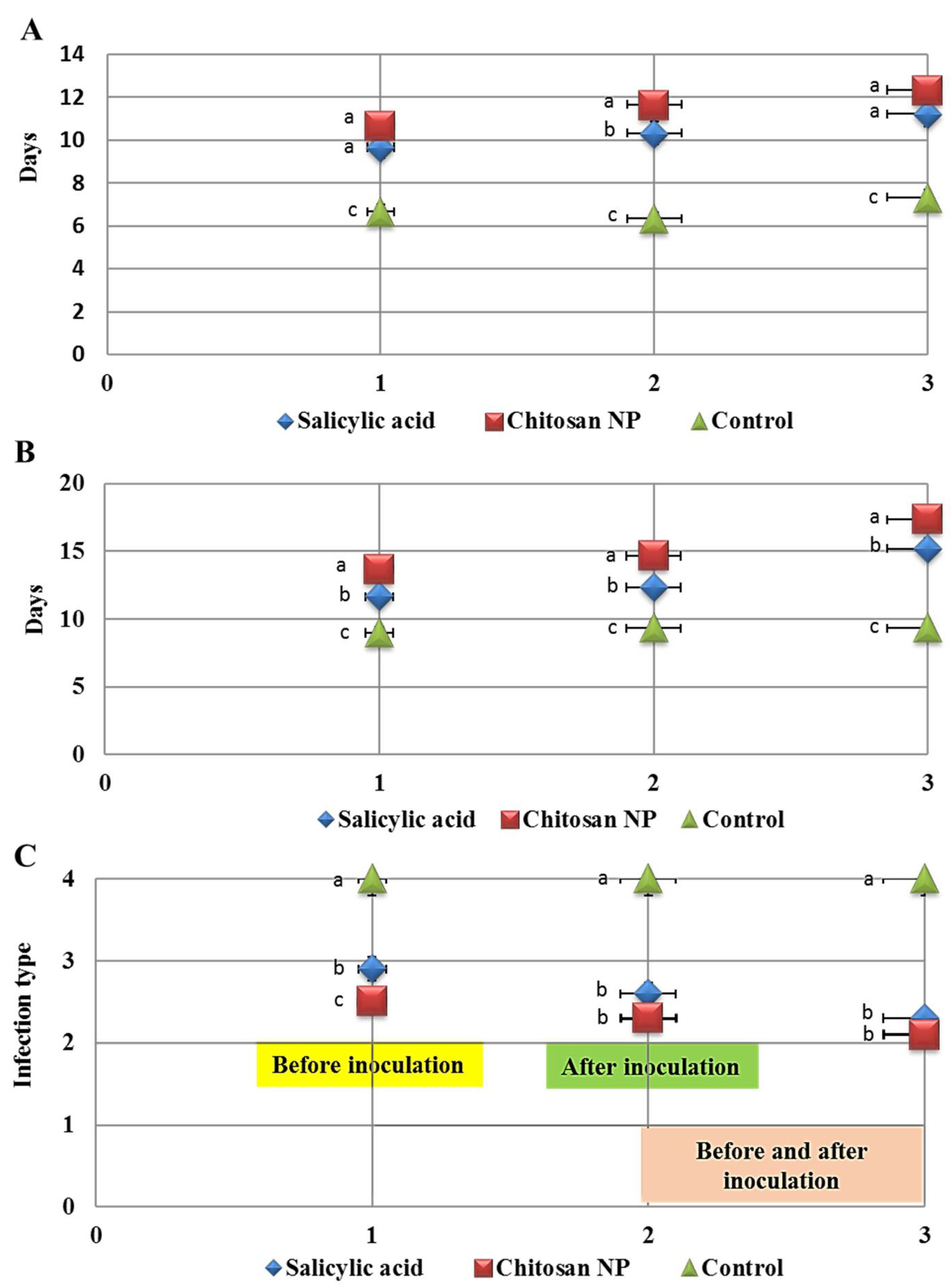
3.2. The Effect of Chitosan Nanoparticles and Salicylic Acid on the Development of Wheat Leaf Rust
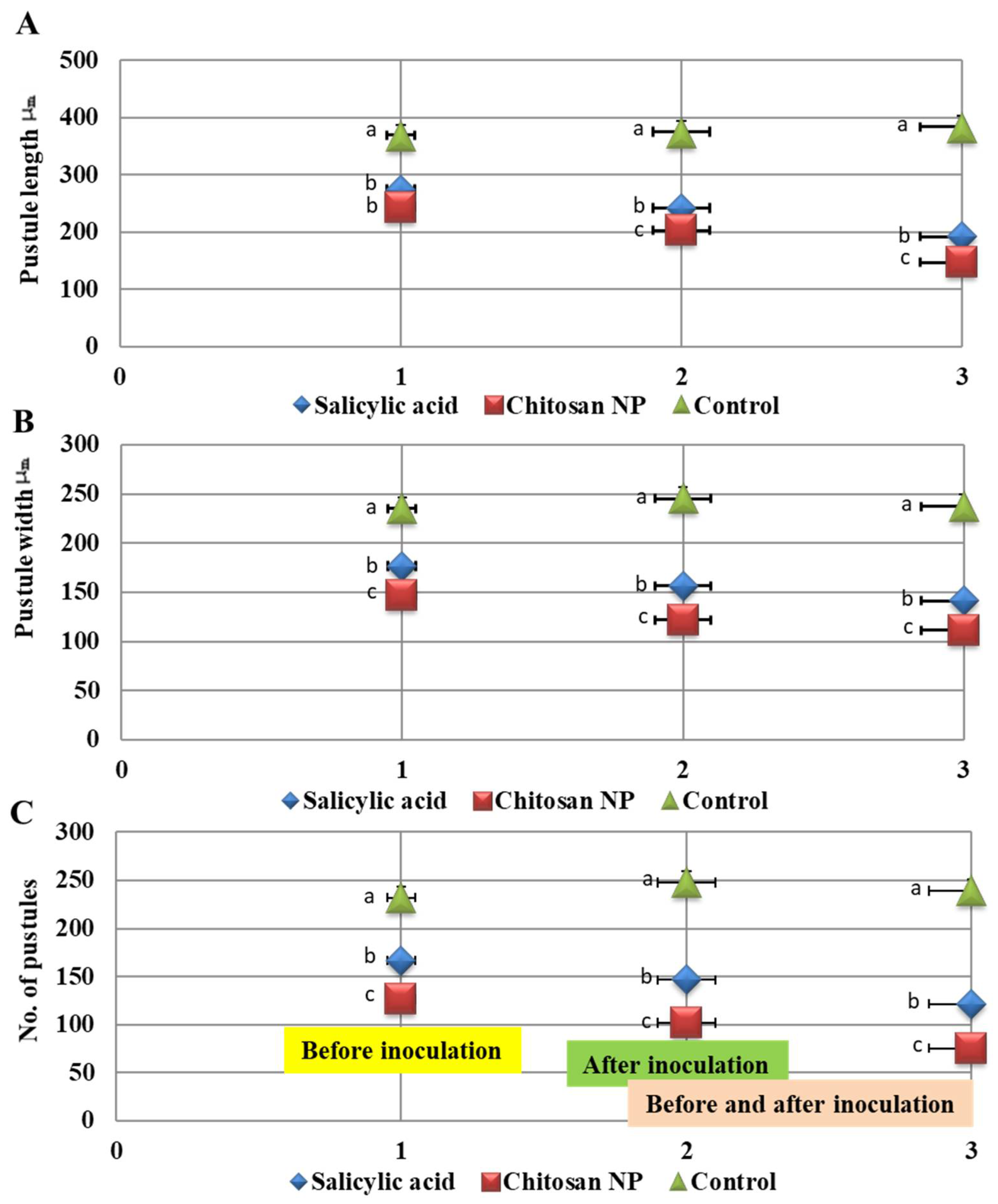
3.3. Effect of Chitosan Nanoparticles and Salicylic acid on Pustules Size and Receptivity
3.4. Effect of Chitosan Nanoparticles and Salicylic Acid on Disease Symptoms
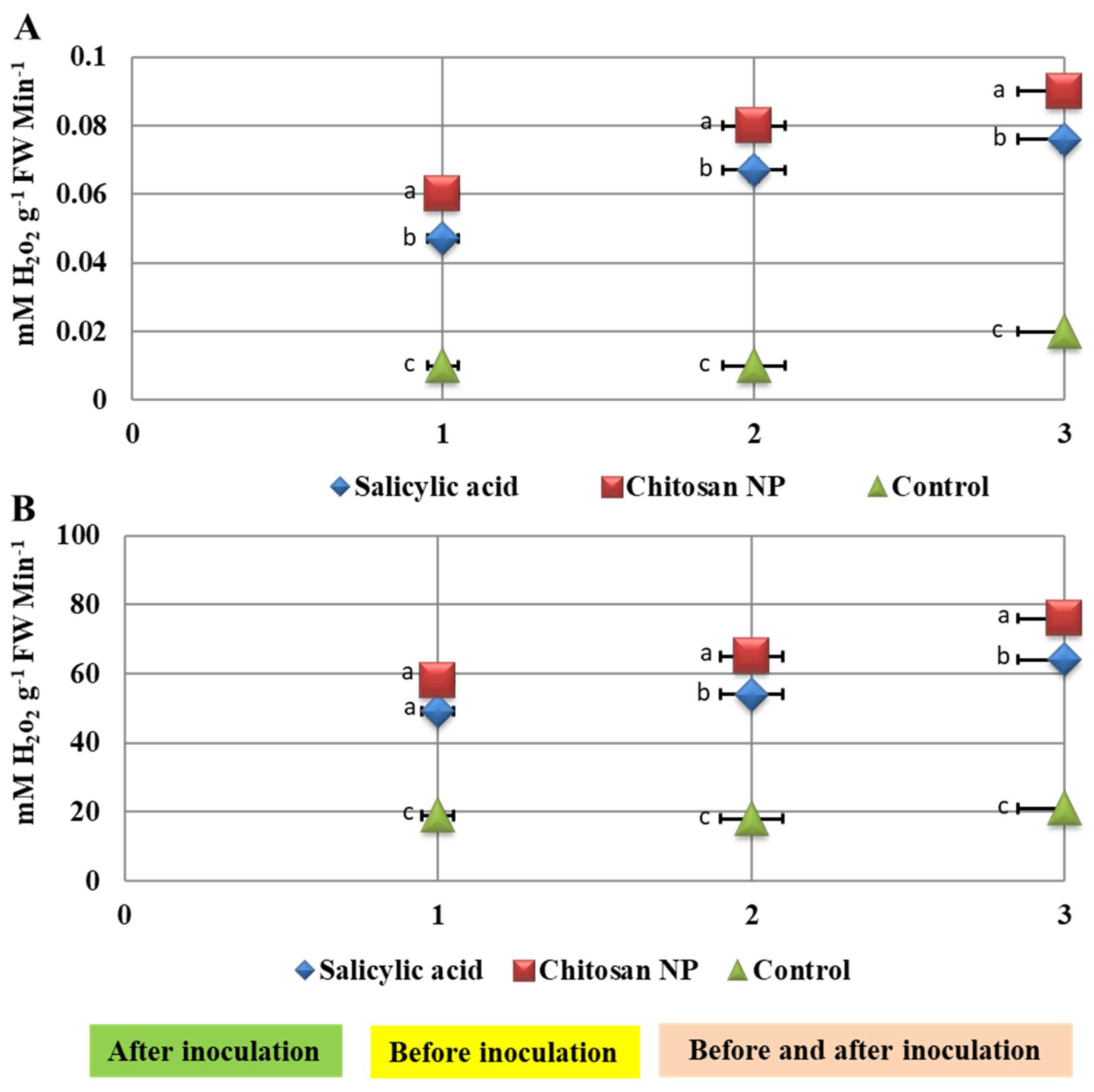
3.5. Effect of Chitosan Nanoparticles and Salicylic Acid Treatments on Enzyme Activities
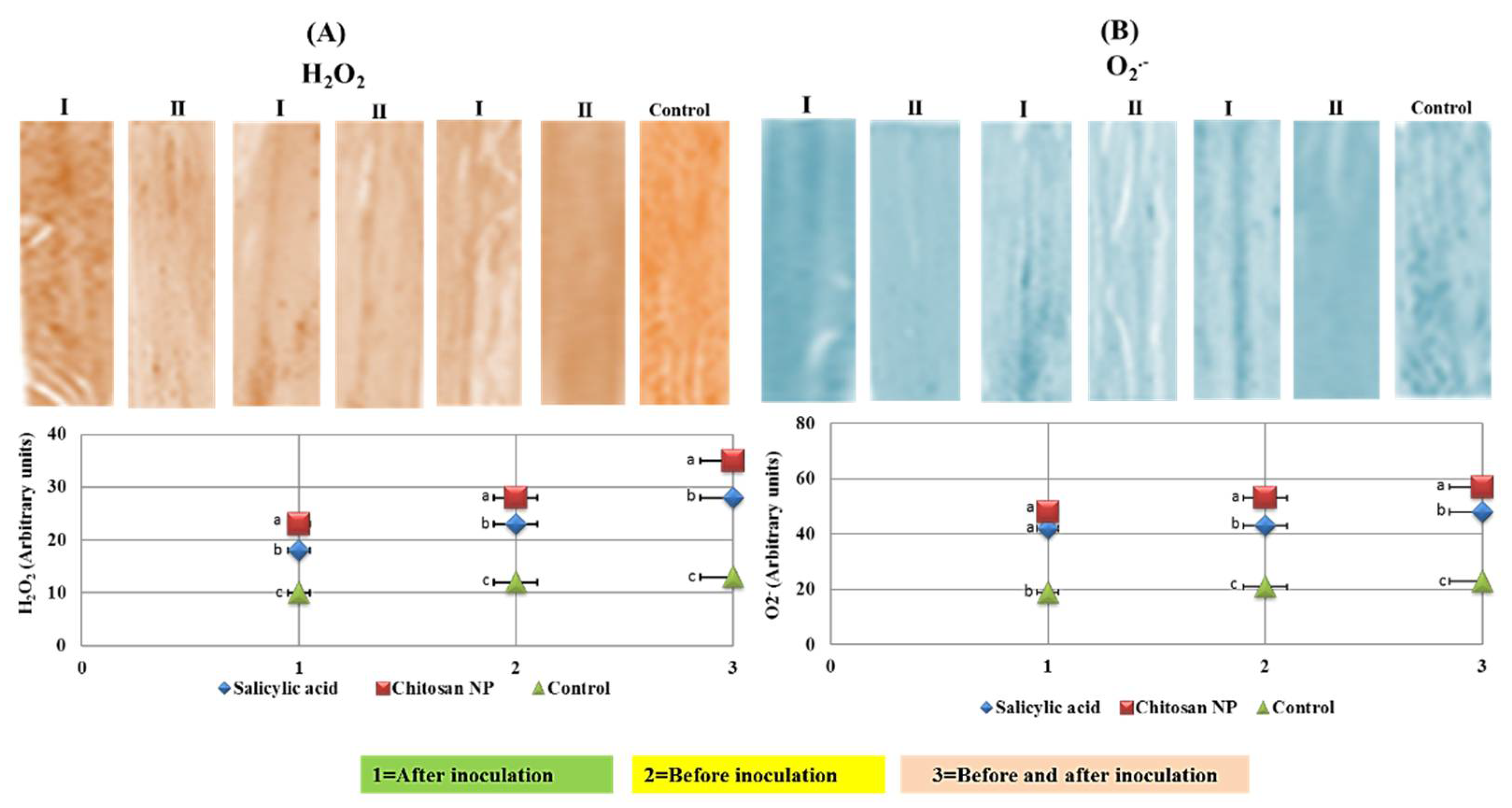
3.6. Histochemical Analysis of Reactive Oxygen Species (ROS)
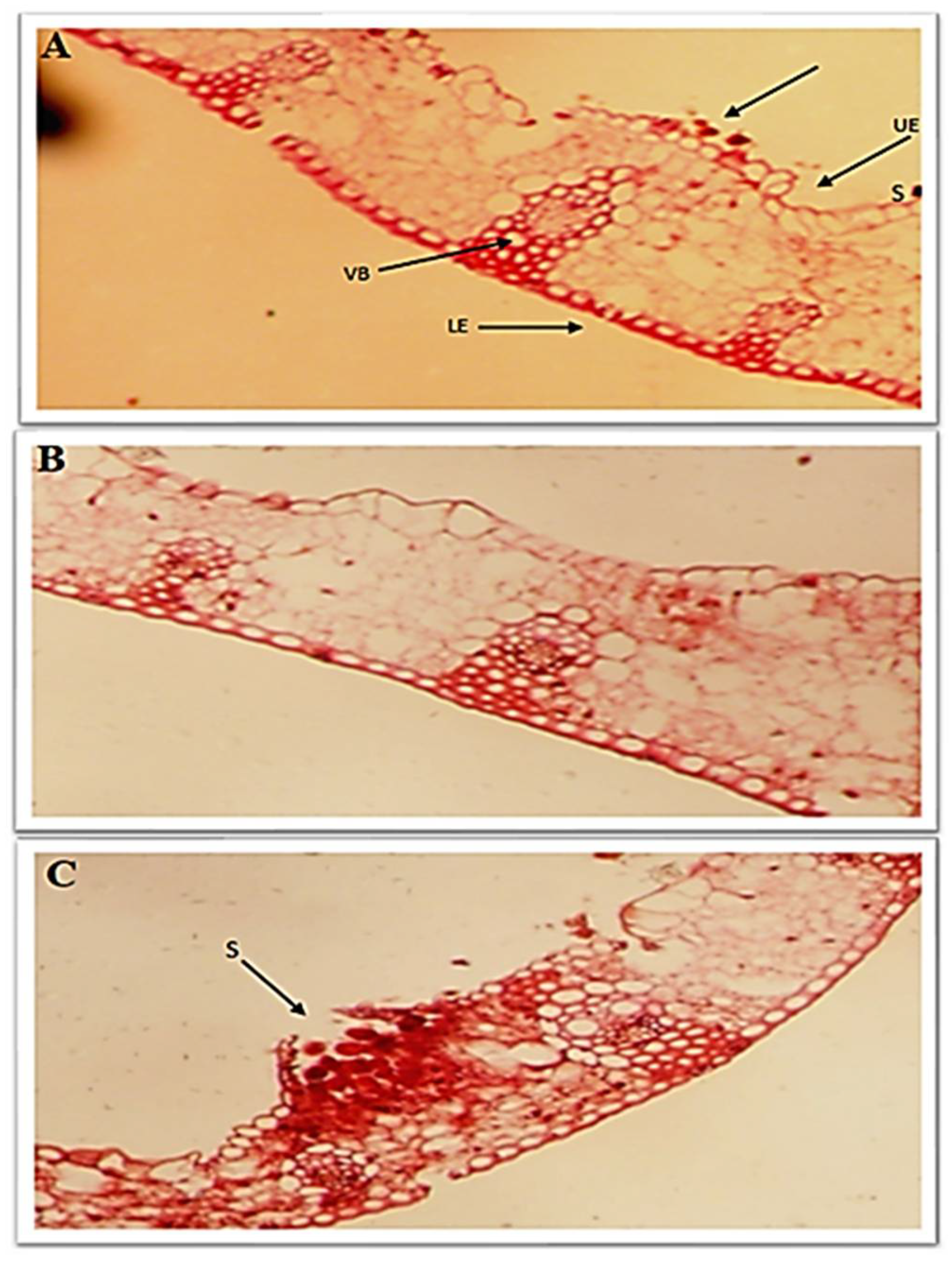
3.7. Effect of Chitosan Nanoparticles and Salicylic Acid on Anatomical Traits
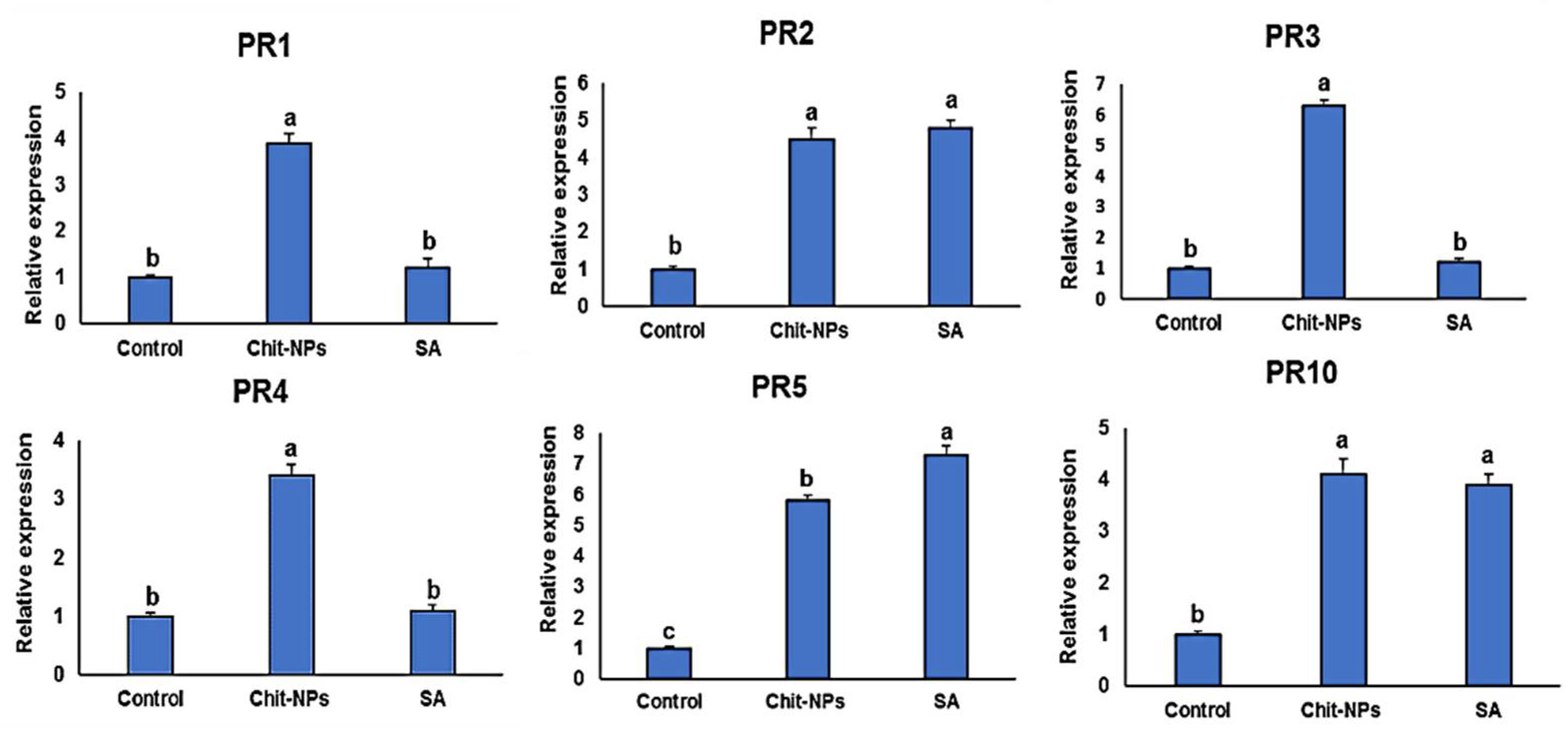
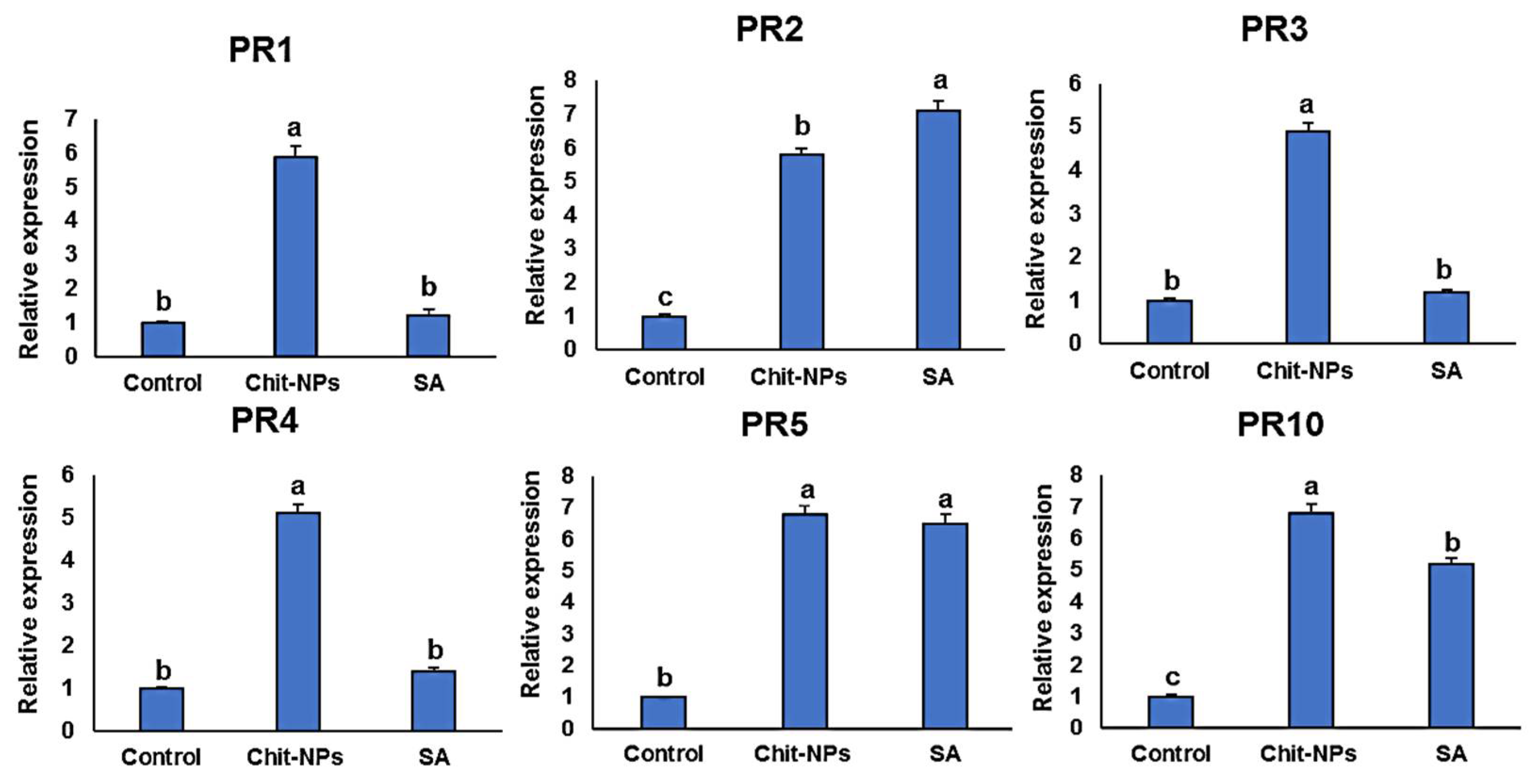
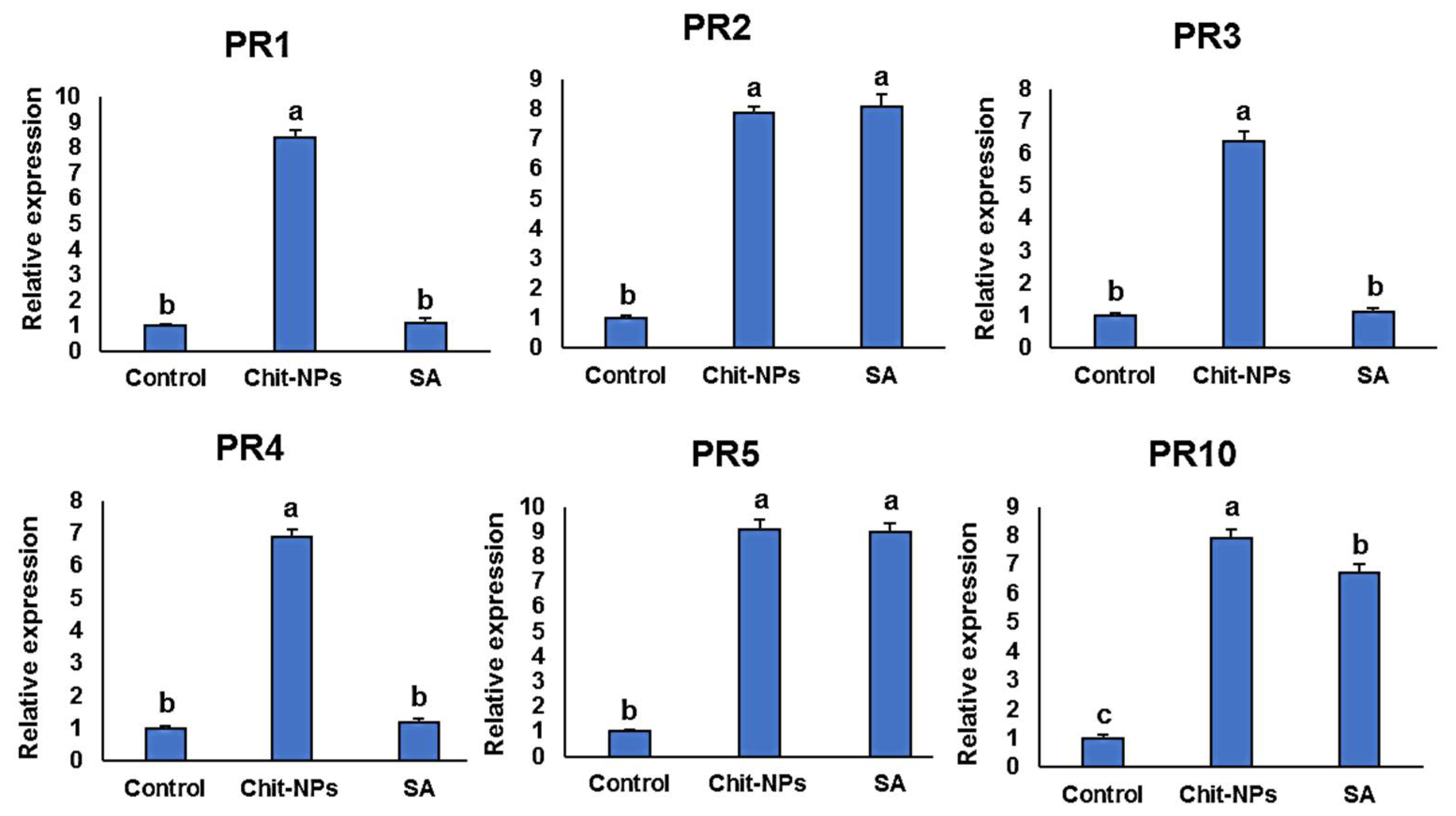
4. Defense-Related Genes Transcriptional Levels
5. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Omara, R.I.; Shahin, A.A.; Ahmed, S.M.; Mostafa, Y.S.; Alamri, S.A.; Hashem, M.; Elsharkawy, M.M. Wheat resistance to stripe and leaf rusts conferred by introgression of slow rusting resistance genes. J. Fungi 2021, 7, 622. [Google Scholar] [CrossRef] [PubMed]
- Nazim, M.; El-Shehidi, A.A.; Abdou, Y.A.; El-Daoudi, Y.H. Yield loss caused by leaf rust on four wheat cultivars under epiphytotic levels. In Proceedings of the 4th Conference of Microbiology, Cairo, Egypt, 24–28 December 1980; pp. 17–27. [Google Scholar]
- Ali, R.G.; Omara, R.I.; Ali, Z.A. Effect of leaf rust infection on yield and technical properties in grains of some Egyptian wheat cultivars. Menoufia J. Plant Prot. 2016, 1, 19–35. [Google Scholar] [CrossRef]
- Gad, A.M.; Abdel-Halim, K.Y.; Seddik, F.A.; Soliman, H.M.A. Comparative of fungicidal efficacy against yellow rust disease in wheat plants in compatibility with some biochemical alterations. Menoufia J. Plant Prot. 2020, 5, 29–38. [Google Scholar] [CrossRef]
- Omara, R.I.; Nehela, Y.; Mabrouk, O.I.; Elsharkawy, M.M. The emergence of new aggressive leaf rust races with the potential to supplant the resistance of wheat cultivars. Biology 2021, 10, 925. [Google Scholar] [CrossRef]
- Xi, K.; Kumar, K.; Holtz, M.D.; Turkington, T.K.; Chapman, B. Understanding the development and management of stripe rust in central Alberta. Can. J. Plant Pathol. 2015, 37, 21–39. [Google Scholar] [CrossRef]
- Hafez, Y.M.; El-Baghdady, N.A. Role of reactive oxygen species in suppression of barley powdery mildew fungus, Blumeria graminis f. sp. hordei with benzothiadiazole and riboflavin. Egypt J. Biol. Pest. Cont. 2013, 23, 125–132. [Google Scholar]
- Hafez, Y.M.; Soliman, N.K.; Saber, M.M.; Imbaby, I.A.; Abd-Elaziz, A.S. Induced resistance against Puccinia triticina the causal agent of wheat leaf rust by chemical inducers. Egypt J. Biol. Pest. Cont. 2014, 24, 173–181. [Google Scholar]
- Omara, R.I.; Kamel, S.M.; Hafez, Y.M.; Morsy, S.Z. Role of non-traditional control treatments in inducing resistance against wheat leaf rust caused by Puccinia triticina. Egypt J. Biol. Pest. Cont. 2015, 25, 335–344. [Google Scholar]
- Al-Juboory, H.; Al-Hadithy, H. Evaluation of chitosan and salicylic acid to induce systemic resistance in eggplant against under plastic Botrytis cinerea house conditions. Indian J. Ecol. 2021, 48, 213–221. [Google Scholar]
- Omara, R.I.; Abdelaal, K.A.A. Biochemical, histopathological and genetic analysis associated with leaf rust infection in wheat plants (Triticum aestivum L.). Physiol. Mol. Plant Pathol. 2018, 104, 48–57. [Google Scholar] [CrossRef]
- El-Saadony, M.T.; Alkhatib, F.M.; Alzahrani, S.O.; Shafi, M.E.; El Abdel-Hamid, S.; Taha, T.F.; Aboelenin, S.M.; Soliman, M.M.; Ahmed, N.H. Impact of mycogenic zinc nanoparticles on performance, behavior, immune response, and microbial load in Oreochromis niloticus. Saudi J. Biol. Sci. 2021, 28, 4592–4604. [Google Scholar] [CrossRef] [PubMed]
- Bano, A. Interactive effects of Ag-nanoparticles, salicylic acid, and plant growth promoting rhizobacteria on the physiology of wheat infected with yellow rust. J. Plant Pathol. 2020, 102, 1215–1225. [Google Scholar] [CrossRef]
- Jasrotia, P.; Kashyap, P.L.; Bhardwaj, A.K.; Kumar, S.; Singh, G.P. Scope and applications of nanotechnology for wheat production: A review of recent advances. Wheat Barley Res. 2018, 10, 1–14. [Google Scholar]
- El-Saadony, M.T.; Saad, A.M.; Najjar, A.A.; Alzahrani, S.O.; Alkhatib, F.M.; Shafi, M.E.; Selem, E.; Desoky, E.-S.; Fouda, S.E.E.; El-Tahan, A.M.; et al. The use of biological selenium nanoparticles to suppress Triticum aestivum L. crown and root rot diseases induced by Fusarium species and improve yield under drought and heat stress. Saudi J. Biol. Sci. 2021, 28, 4461–4471. [Google Scholar] [CrossRef] [PubMed]
- El-Saadony, M.T.; Sitohy, M.Z.; Ramadan, M.F.; Saad, A.M. Green nanotechnology for preserving and enriching yogurt with biologically available iron (II). Innov. Food Sci. Emerg. Technol. 2021, 69, 102645. [Google Scholar] [CrossRef]
- Elizabath, A.; Babychan, M.; Mathew, A.M.; Syriac, G.M. Application of nanotechnology in agriculture. Int. J. Pure Appl. Biosci. 2019, 7, 131–139. [Google Scholar] [CrossRef]
- Elieh-Ali-Komi, D.; Hamblin, M.R. Chitin and chitosan: Production and application of versatile biomedical nanomaterials. Int J Adv. Res. 2016, 4, 411–427. [Google Scholar]
- Kocięcka, J.; Liberacki, D. The potential of using chitosan on cereal crops in the face of climate change. Plants 2021, 10, 1160. [Google Scholar] [CrossRef]
- Abd El-Hack, M.E.; El-Saadony, M.T.; Shafi, M.E.; Zabermawi, N.M.; Arif, M.; Batiha, G.E.; Khafaga, A.F.; Abd El-Hakim, Y.M.; Al-Sagheer, A.A. Antimicrobial and antioxidant properties of chitosan and its derivatives and their applications: A review. Int. J. Biol. Macromol. 2020, 164, 2726–2744. [Google Scholar] [CrossRef]
- Omar, H.S.; Al Mutery, A.; Osman, N.H.; Reyad, N.E.H.A.; Abou-Zeid, M.A. Genetic diversity, antifungal evaluation and molecular docking studies of Cu-chitosan nanoparticles as prospective stem rust inhibitor candidates among some Egyptian wheat genotypes. PLoS ONE 2021, 16, e0257959. [Google Scholar] [CrossRef]
- Pichyangkura, R.; Chadchawan, S. Biostimulant activity of chitosan in horticulture. Sci. Hortic. 2015, 196, 49–65. [Google Scholar] [CrossRef]
- Soleiman, N.H.; Solis, I.; Ammar, A.; Dreisigacker, S.; Soleiman, M.H.; Martinez, F. Resistance to leaf rust in a set of durum wheat cultivars and landraces in Spain. J. Plant Pathol. 2014, 96, 353–362. [Google Scholar]
- Johnston, C.O.; Browder, L.E. Seventh revision of the international register of physiologic races of Puccinia recondita f.sp. tritici. Plant Dis. Report 1966, 50, 756–760. [Google Scholar]
- Nair, K.R.S.; Ellingboe, A.H. A method of controlled inoculations with condiospores of Erysiphe graminis var. tritici. Phytopatholology 1962, 52, 417. [Google Scholar]
- Reifshneider, F.J.B.; Bolitexa, L.S.; Occhiena, E.M. Powdery mildew of melon (Cucumis melo) caused by Sphaerotheca fuliginea in Brazil. Plant Dis. 1985, 69, 1069–1070. [Google Scholar]
- Parlevliet, J.E. Partial resistance of barley to leaf rust, Puccinia hordei. I. effect of cultivars and development stage on latent period. Euphytica 1975, 24, 21–27. [Google Scholar] [CrossRef]
- Parlevliet, J.E.; Kuiper, H.J. Partial resistance of barley to leaf rust, Puccinia hordei. IV. Effect of cultivars and development stage on infection frequency. Euphytica 1977, 26, 249–255. [Google Scholar] [CrossRef]
- Menzies, J.G.; Belanger, R.R. Recent advances in cultural management of diseases of greenhouse crops. Can. J. Plant Pathol. 1996, 18, 186–193. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Hammerschmidt, R.; Nuckles, E.M.; Kuć, J. Association of enhanced peroxidase activity with induced systemic resistance of cucumber to Colletotrichum lagenarium. Physiol. Plant Pathol. 1982, 20, 73–82. [Google Scholar] [CrossRef]
- Huckelhoven, R.; Fodor, J.; Preis, C.; Kogel, K.H. Hypersensitive cell death and papilla formation in barley attacked by the powdery mildew fungus are associated with hydrogen peroxide but not with salicylic acid accumulation. Plant Physiol. 1999, 119, 1251–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nassar, M.A.; El-Sahhar, K.F. Botanical Preparations and Microscopy (Microtechnique); Academic Bookshop: Dokki, Giza, Egypt, 1998; p. 219. [Google Scholar]
- Elsharkawy, M.; Derbalah, A.; Hamza, A.; El-Shaer, A. Zinc oxide nanostructures as a control strategy of bacterial speck of tomato caused by Pseudomonas syringae in Egypt. Environ. Sci. Pollut Res. 2020, 27, 19049–19057. [Google Scholar] [CrossRef]
- Desmond, O.J.; Edgar, C.I.; Manners, J.M.; Maclean, D.J.; Schenk, P.M.; Kazan, K. Methyl jasmonate induced gene expression in wheat delays symptom development by the crown rot pathogen Fusarium pseudograminearum. Physiol. Mol. Plant Pathol. 2006, 67, 171–179. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.M. Epidemiology and control of stripe rust (Puccinia striiformis f sp. tritici) on wheat. Can. J. Plant Pathol. 2005, 27, 314–337. [Google Scholar] [CrossRef]
- Hossard, L.; Philibert, A.; Bertrand, M.; Colnenne-David, C.; Debaeke, P.; Munier-Jolain, N.; Jeuffroy, M.H.; Richard, G.; Makowski, D. Effects of halving pesticide use on wheat production. Sci. Rep. 2015, 4, 4405. [Google Scholar] [CrossRef] [Green Version]
- El Hadrami, A.; Adam, L.R.; El Hadrami, I.; Daayf, F. Chitosan in plant protection. Mar. Drugs 2010, 8, 968–987. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhao, X.; Han, X.; Du, Y. Antifungal activity of oligo chitosan against Phytophthora capsici and other plant pathogenic fungi in vitro. Pestic. Biochem. Physiol. 2007, 87, 220–228. [Google Scholar] [CrossRef]
- Ata, A.A.; El-Samman, M.G.; Moursy, M.A.; Mostafa, M.H. Inducing resistance against rust disease of sugar beet by certain chemical compounds. Egypt J. phytopathol. 2008, 36, 113–132. [Google Scholar]
- Hassan, O.; Chang, T. Chitosan for eco-friendly control of plant disease. Asian J. Plant Pathol. 2017, 11, 53–70. [Google Scholar] [CrossRef]
- Benhamou, N.; Kloepper, J.W.; Tuzan, B.B. Induction of resistance against Fusarium wilt of tomato by combination of chitosan with an endophytic bacterial stain: Ultrastructure and cytochemistry of host response. Planta 1998, 204, 153–168. [Google Scholar] [CrossRef]
- Ragab, M.M.; Ragab, M.M.; El-Nagar, M.A.; Farrag, E.S. Effect of chitosan and its derivatives as an antifungal and preservative agent on storage of tomato fruits. Egypt J. Phytopathol. 2001, 29, 107–116. [Google Scholar]
- Bautista, B.S.; Hern, N.; Indez, L.; Bosquez, M.E.; Wil, C.L. Effect of chitosan and plant extracts on growth of Colletotrichum gloeosporioides, anthracnose level and quality of papaya fruit. Crop Prot. 2003, 22, 1087–1092. [Google Scholar] [CrossRef]
- Rabea, E.; Badawy, M.T.; Stevens, C.; Smagghe, G.; Steurbaut, W. Chitosan as antimicrobial agent: Applications and Mode of action. Biomacromolecules 2003, 4, 1457–1465. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.K.; Das, A.K.; Gantait, S.; Kumar, V.; Gurel, E. Applications of carbon nanomaterials in the plant system: A perspective view on the pros and cons. Sci. Total Environ. 2019, 667, 485–499. [Google Scholar] [CrossRef]
- Catherine, B.; Marc, O.; Philippe, T.; Jacques, D. Cloning and expression analysis of cDNAs corresponding to genes activated in cucumber showing systemic acquired resistance after BTH treatment. BMC Plant Biol. 2004, 4, 15. [Google Scholar]
- Yang, S.; Gaojian, L. ROS and diseases: Role in metabolism and energy supply. Mol. Cell. Biochem. 2020, 467, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Xing, K.; Zhu, X.; Peng, X.; Qin, S. Chitosan antimicrobial and eliciting properties for pest control in agriculture: A review. Agron. Sustain. Dev. 2015, 35, 569–588. [Google Scholar] [CrossRef] [Green Version]
- Christian, N.; Jean, P.M. Salicylic acid induction defiant mutants of Arabidiopsis express PR-2 and PR-5 and accumulate high levels of camalexin after pathogen inoculation. Plant Cell 1999, 11, 1393–1404. [Google Scholar]
- Omara, R.I.; Essa, T.A.; Khalil, A.A.; Elsharkawy, M.M. A case study of non-traditional treatments for the control of wheat stem rust disease. Egypt J. Biol. Pest. Cont. 2020, 30, 83. [Google Scholar] [CrossRef]
- Quiroga, M.; Guerrero, C.; Botella, M.A.; Barcelo, A.; Amaya, I.; Meding, M.; Alonso, F.J.; Forcheti, S.M.; Tigir, H.; Vulpuesta, V.A. Tomato peroxidase involved in the synthesis of lignin and suberin. Plant Physiol. 2000, 122, 1119–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruno, M.M.; Ulrik, N.; Lilliane, G.; Hans-joachim, R. Specific inhibition of lignifications breaks hypersensitive resistance of wheat to stem rust. Plant Physiol. 1990, 93, 465–470. [Google Scholar]
- Cohen, Y. The BABA story of induced resistance. Phytoparasitica 2001, 29, 375–378. [Google Scholar] [CrossRef]
- Chang, M.M.; Hadwiger, L.A.; Horovits, D. Molecular characterization of a pea 1,3 glucanase induced by Fusarium solani and chitosan challenge. Plant Mol. Biol. 1992, 20, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Siddaiah, C.N.; Prasanth, K.V.H.; Satyanarayana, N.R.; Mudili, V.; Gupta, V.K.; Kalagatur, N.K.; Satyavati, T.; Dai, X.-F.; Chen, J.-Y.; Mocan, A. Chitosan nanoparticles having higher degree of acetylation induce resistance against pearl millet downy mildew through nitric oxide generation. Sci. Rep. 2018, 8, 2485. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Wang, Z.-Y.; Li, T.-T.; Wang, F.; An, J. The role of chitosan in polyphenols accumulation and induction of defense enzymes in Pinus koraiensis seedlings. Chin. J. Plant Ecol. 2014, 38, 749. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Forward Primer (5′ . . . . . 3′) | Reverse Primer (5′ . . . . . 3′) |
|---|---|---|
| PR1 (basic) | CTGGAGCACGAAGCTGCAG | CGAGTGCTGGAGCTTGCAGT |
| PR2 | CTCGACATCGGTAACGACCAG | GCGGCGATGTACTTGATGTTC |
| PR3 | AGAGATAAGCAAGGCCACGTC | GGTTGCTCACCAGGTCCTTC |
| PR4 | CGAGGATCGTGGACCAGTG | GTCGACGAACTGGTAGTTGACG |
| PR5 | ACAGCTACGCCAAGGACGAC | CGCGTCCTAATCTAAGGGCAG |
| PR10 | TTAAACCAGCACGAGAAACATCAG | ATCCTCCCTCGATTATTCTCACG |
| β-tubulin | GCCATGTTCAGGAGGAAGG | CTCGGTGAACTCCATCTCGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elsharkawy, M.M.; Omara, R.I.; Mostafa, Y.S.; Alamri, S.A.; Hashem, M.; Alrumman, S.A.; Ahmad, A.A. Mechanism of Wheat Leaf Rust Control Using Chitosan Nanoparticles and Salicylic Acid. J. Fungi 2022, 8, 304. https://doi.org/10.3390/jof8030304
Elsharkawy MM, Omara RI, Mostafa YS, Alamri SA, Hashem M, Alrumman SA, Ahmad AA. Mechanism of Wheat Leaf Rust Control Using Chitosan Nanoparticles and Salicylic Acid. Journal of Fungi. 2022; 8(3):304. https://doi.org/10.3390/jof8030304
Chicago/Turabian StyleElsharkawy, Mohsen Mohamed, Reda Ibrahim Omara, Yasser Sabry Mostafa, Saad Abdulrahman Alamri, Mohamed Hashem, Sulaiman A. Alrumman, and Abdelmonim Ali Ahmad. 2022. "Mechanism of Wheat Leaf Rust Control Using Chitosan Nanoparticles and Salicylic Acid" Journal of Fungi 8, no. 3: 304. https://doi.org/10.3390/jof8030304