
Myco-Synthesized Molluscicidal and Larvicidal Selenium Nanoparticles: A New Strategy to Control Biomphalaria alexandrina Snails and Larvae of Schistosoma mansoni with an In Silico Study on Induced Oxidative Stress


 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation Conditions of the Fungal Isolate
2.2. Molecular Identification
2.3. SeNPs Myco-Synthesis by P. chrysogenum
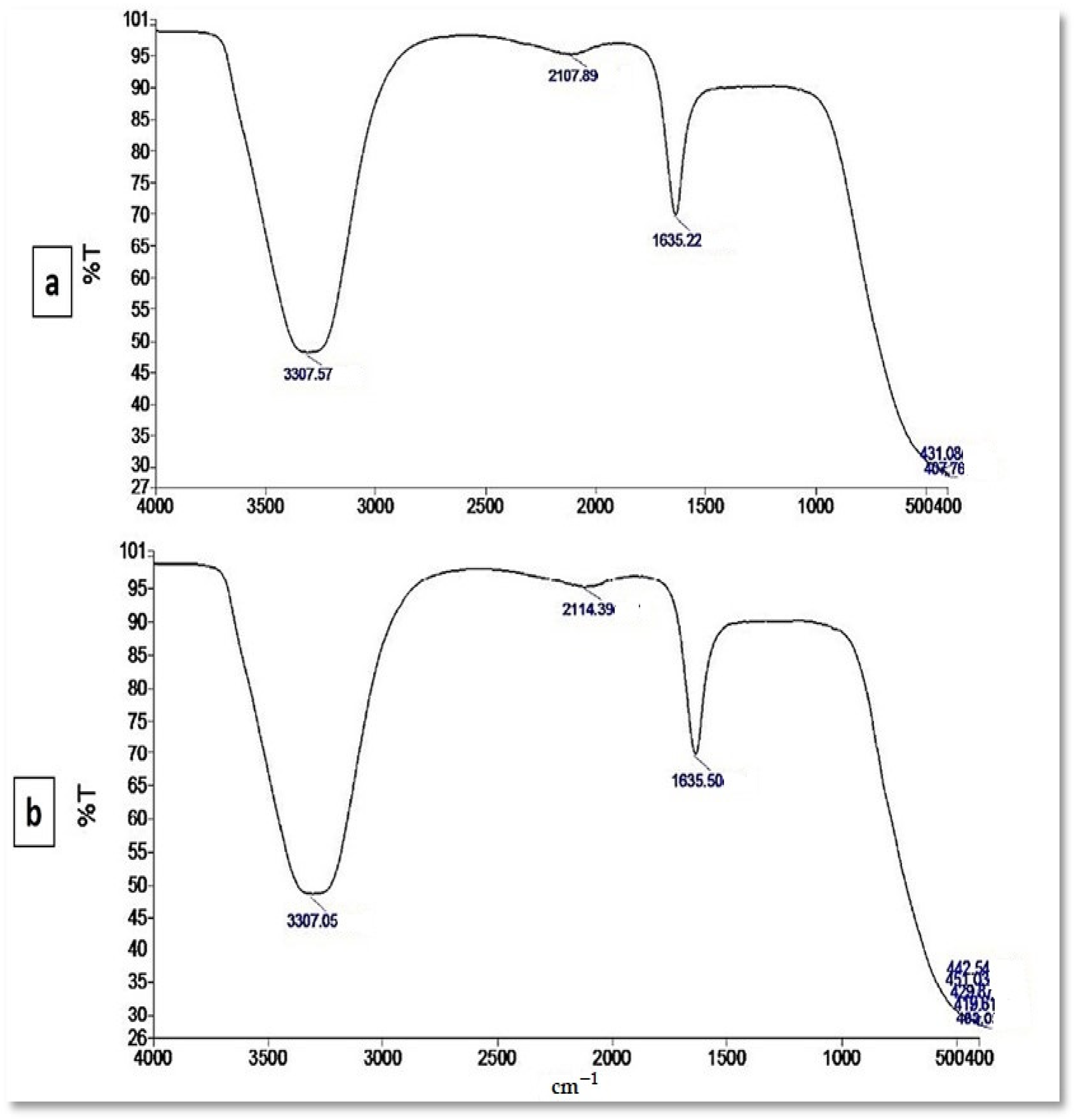
2.4. Characterization of the Myco-Synthesized SeNPs
2.5. Investigation of Molluscicidal Activity of SeNPs
2.5.1. Snails
2.5.2. Molluscicidal Activity of SeNPs
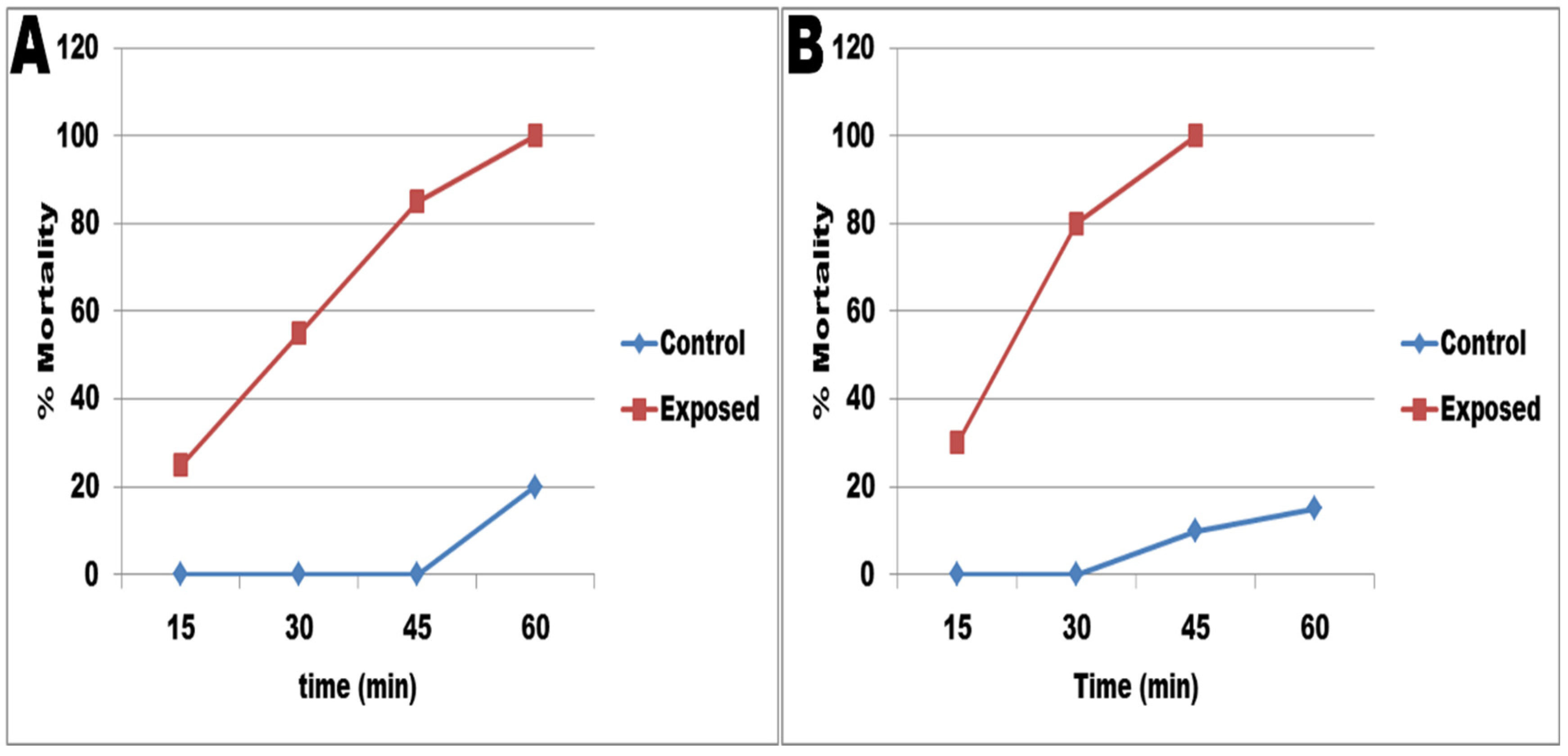
2.5.3. Miracidicidal and Cercaricidal Activity
2.6. Experimental Design
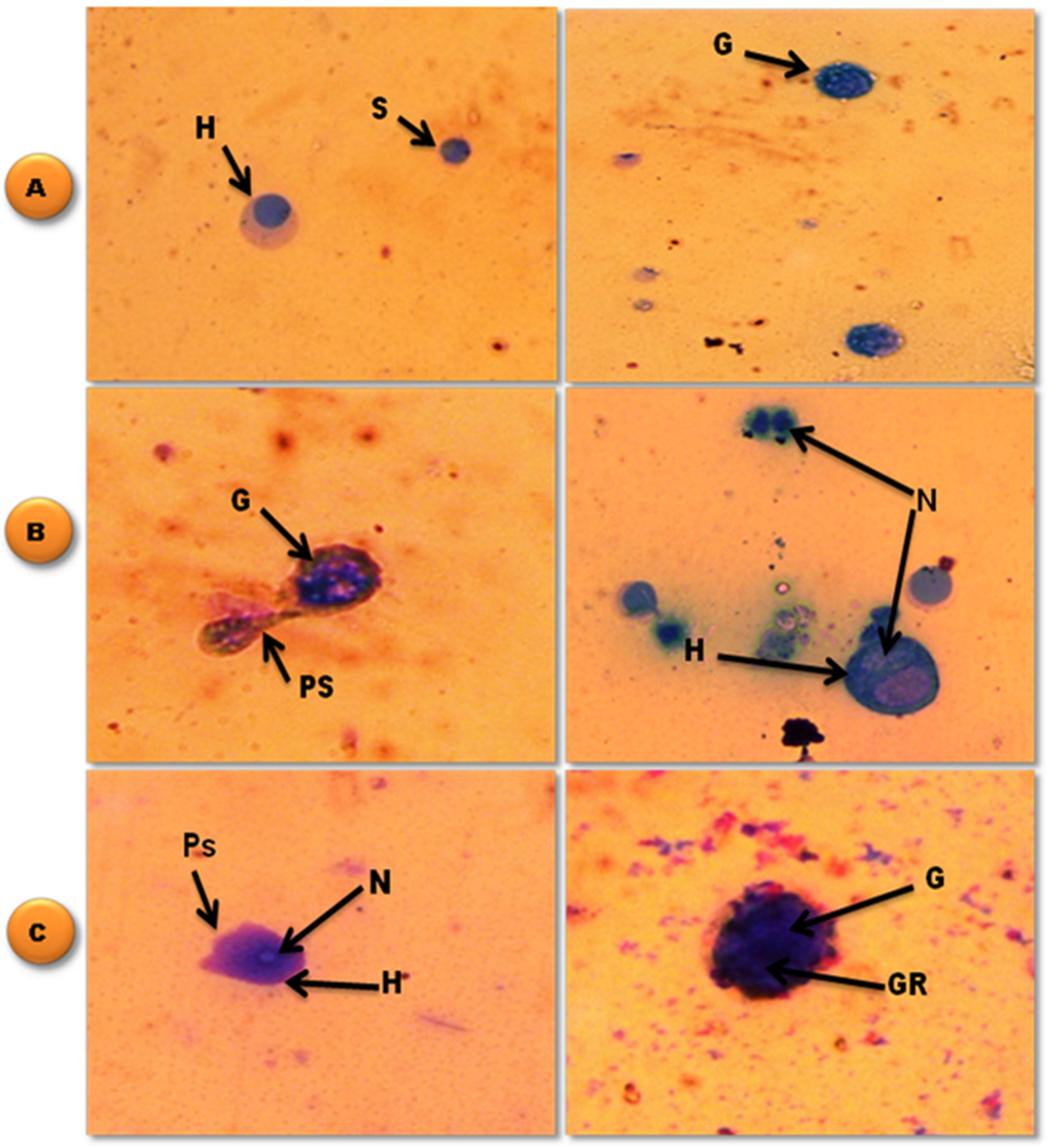
2.6.1. Hemolymph and Light Microscopy Preparation
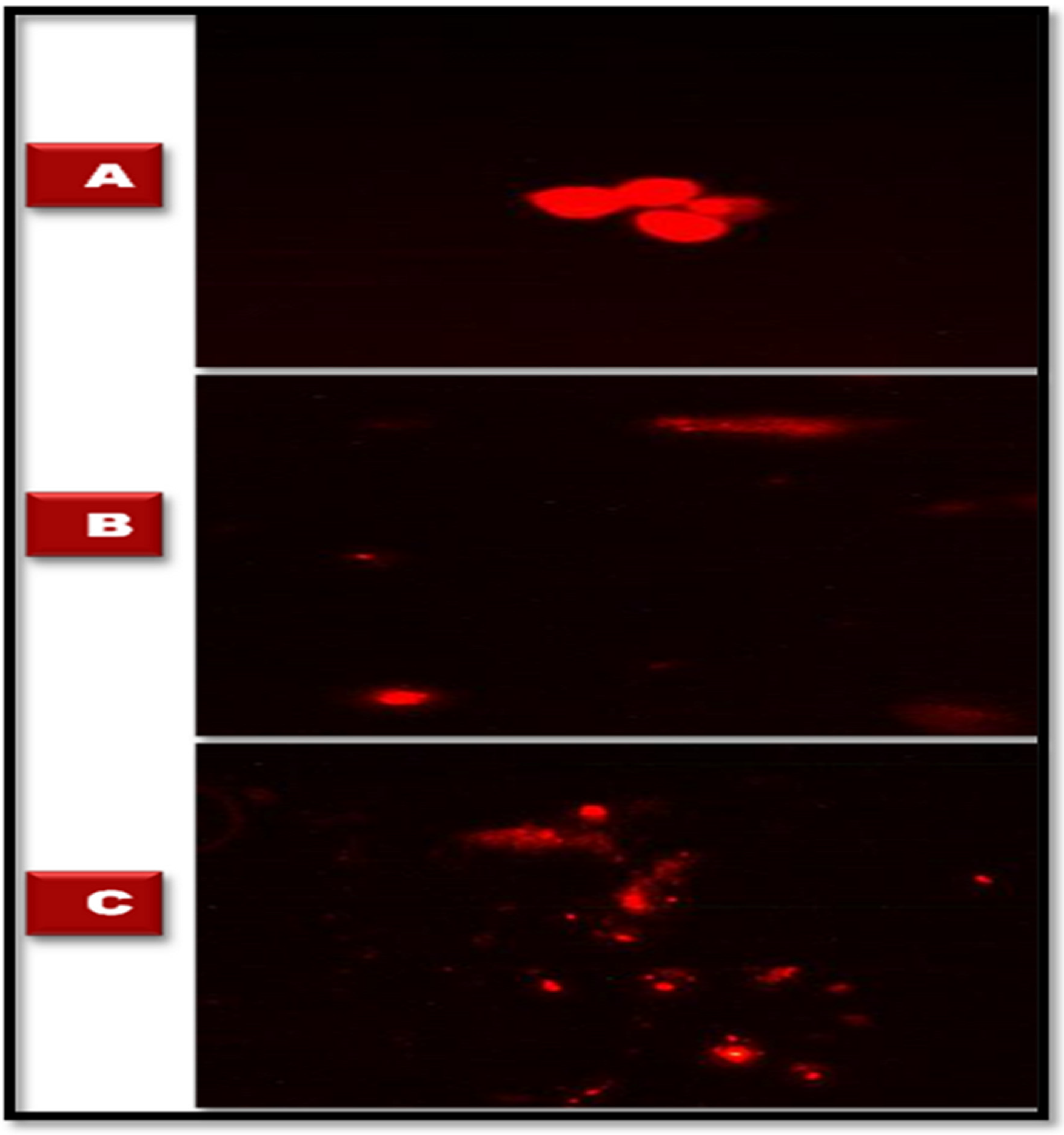
2.6.2. Comet Assay
2.6.3. Tissue Preparation
Determination of Testosterone (T) and Estradiol (E2) Hormones Concentrations
Investigation of the Antioxidant Responses: Superoxide Dismutase (SOD); Glutathione S-Transferase (GST), Nitric Oxide (NO), Malondialdehyde (MDA), and Total Antioxidant Capacity (TAC)
2.7. The Molecular Docking Study
2.8. Statistical Analysis
3. Results
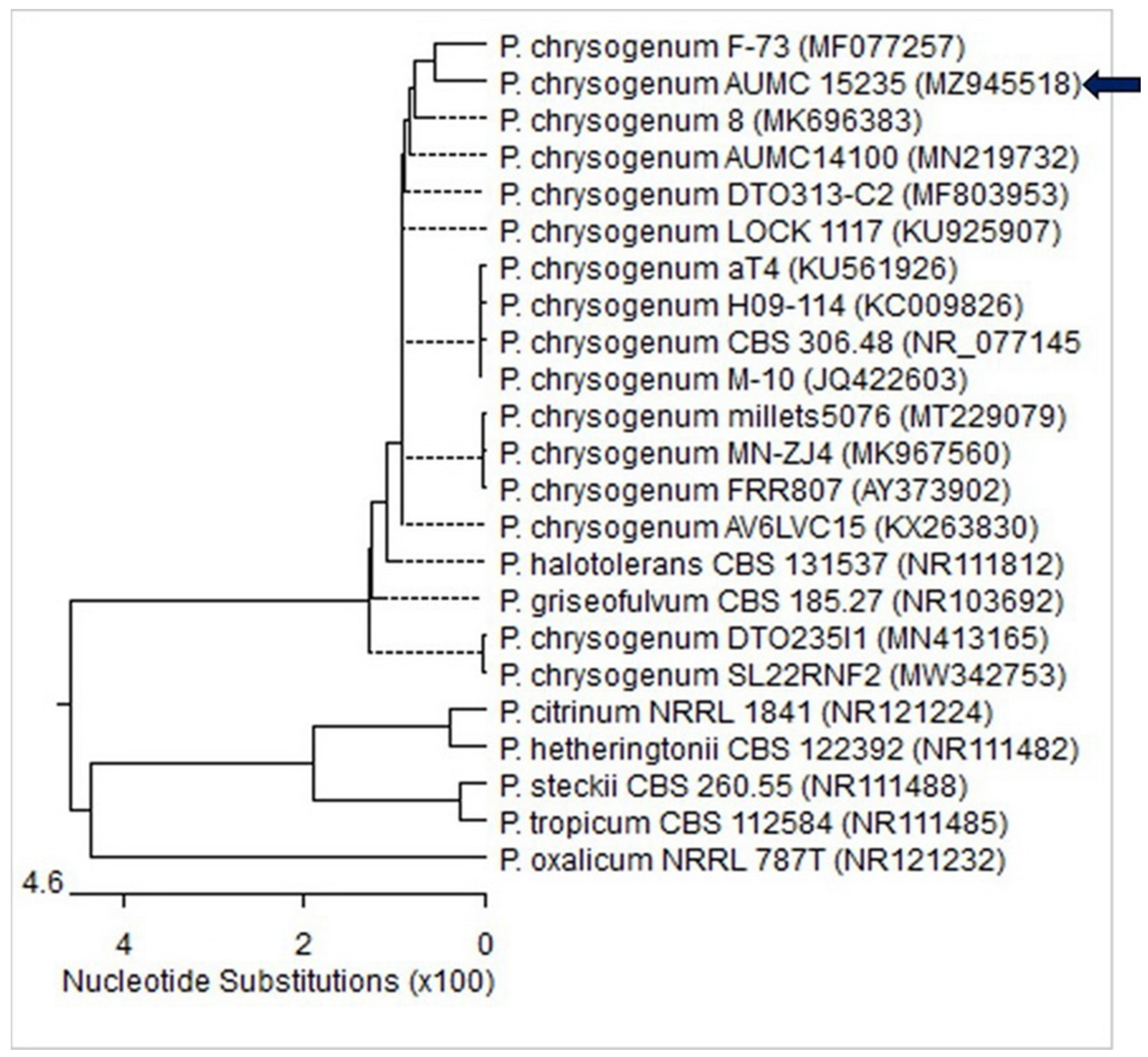
3.1. Molecular Identification of the Fungal Strain
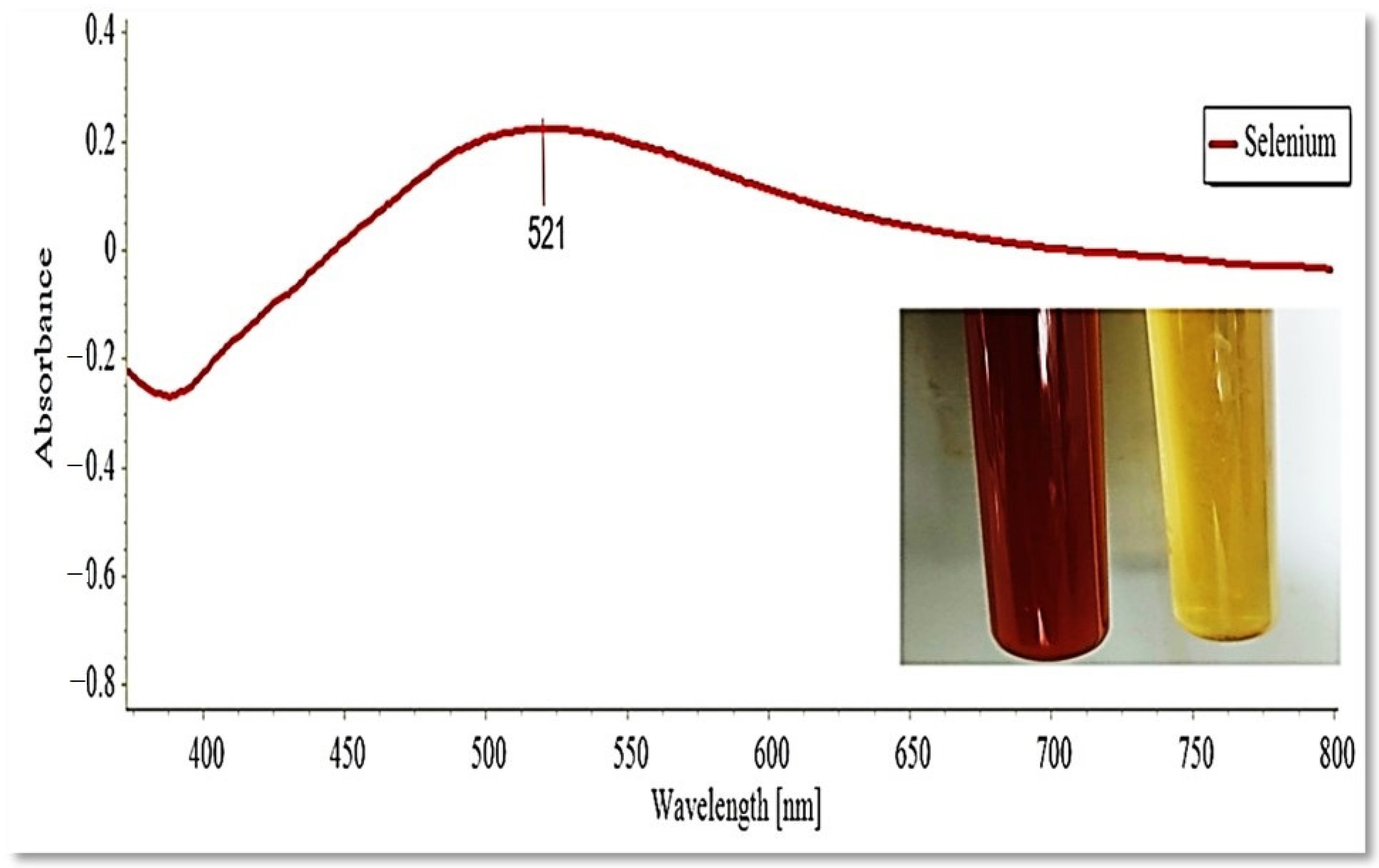
3.2. The Myco-Synthesis of SeNPs by P. chrysogenum
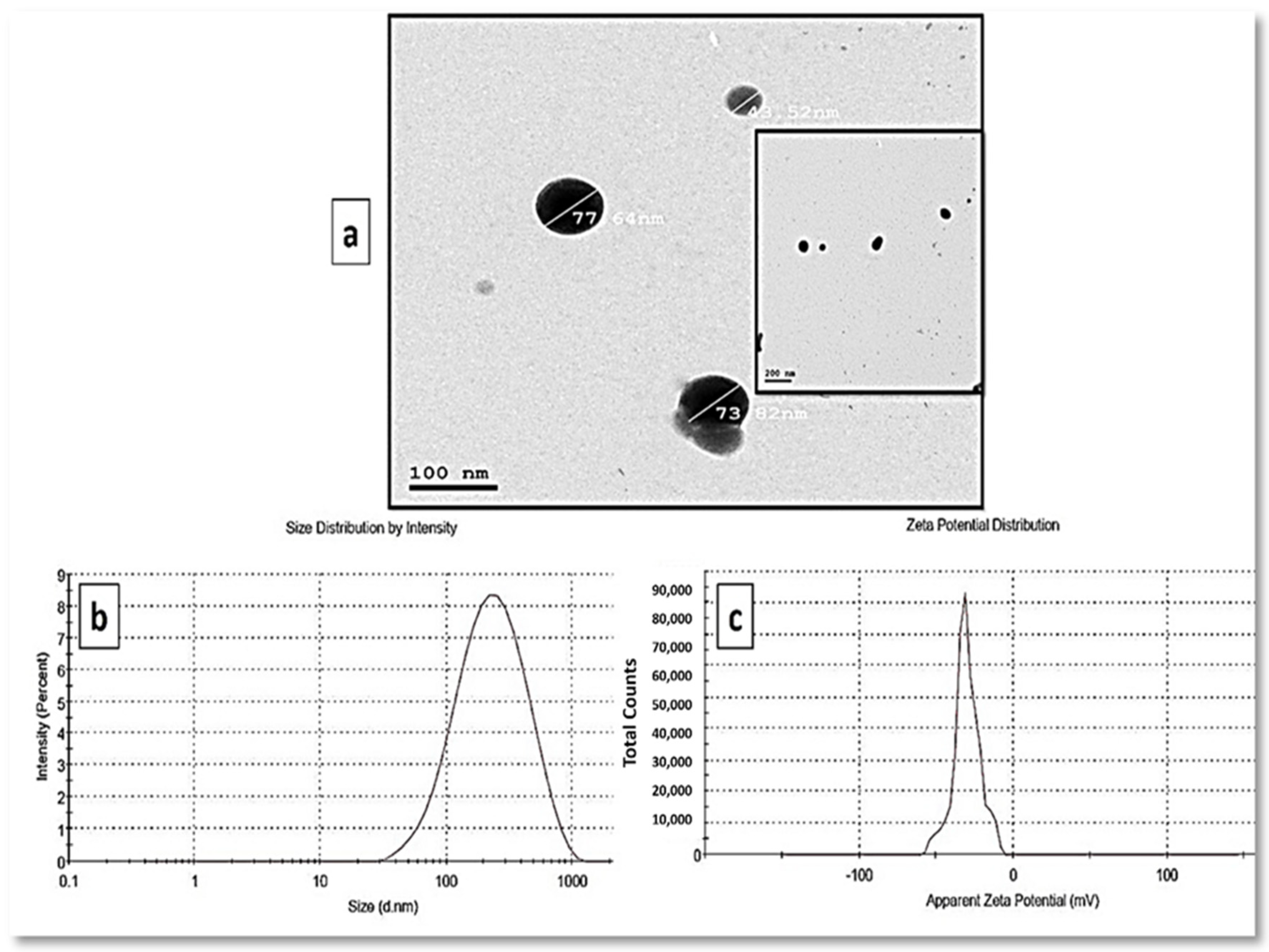
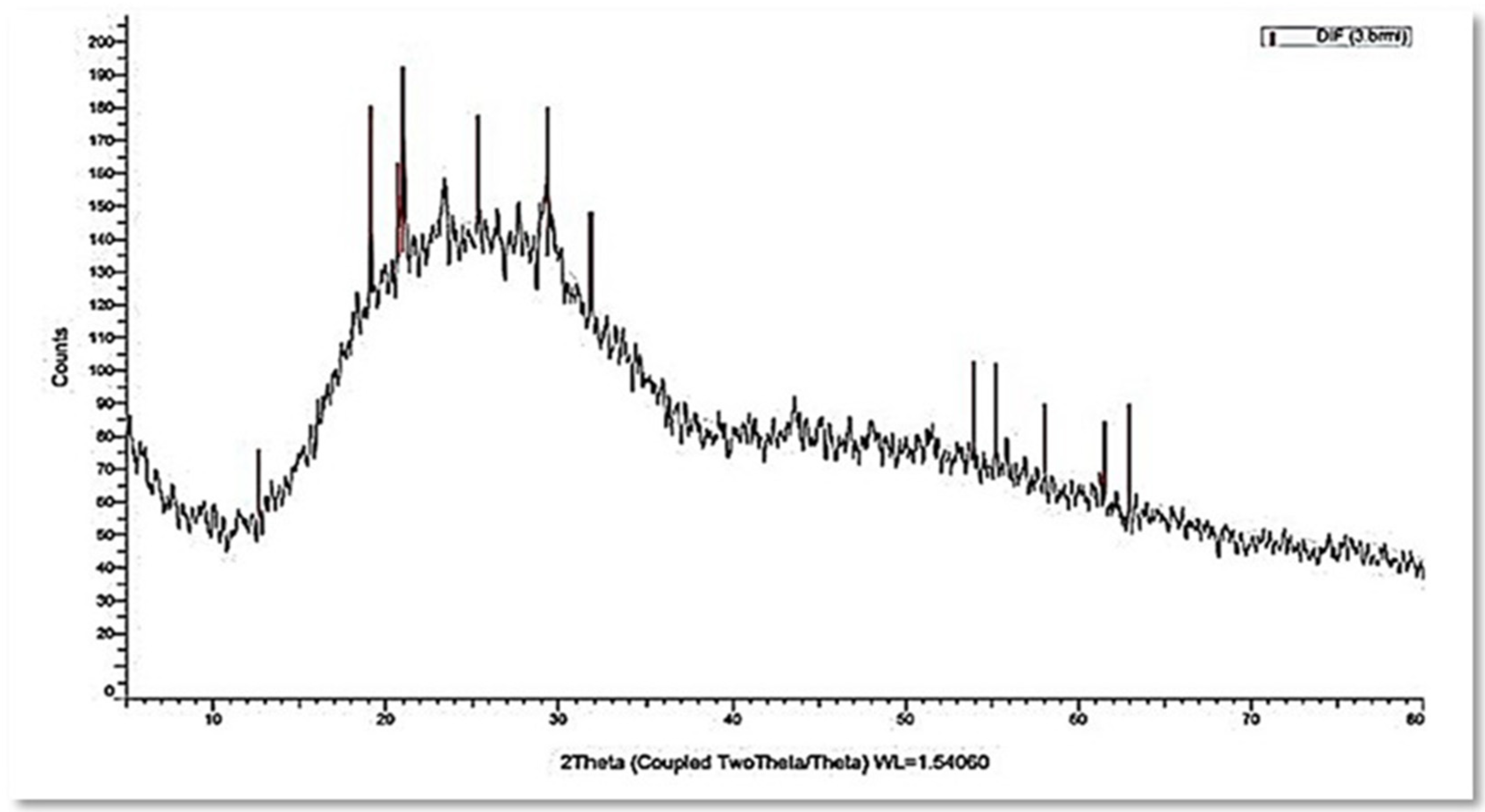
3.3. Characterization of the Myco-Synthesized SeNPs
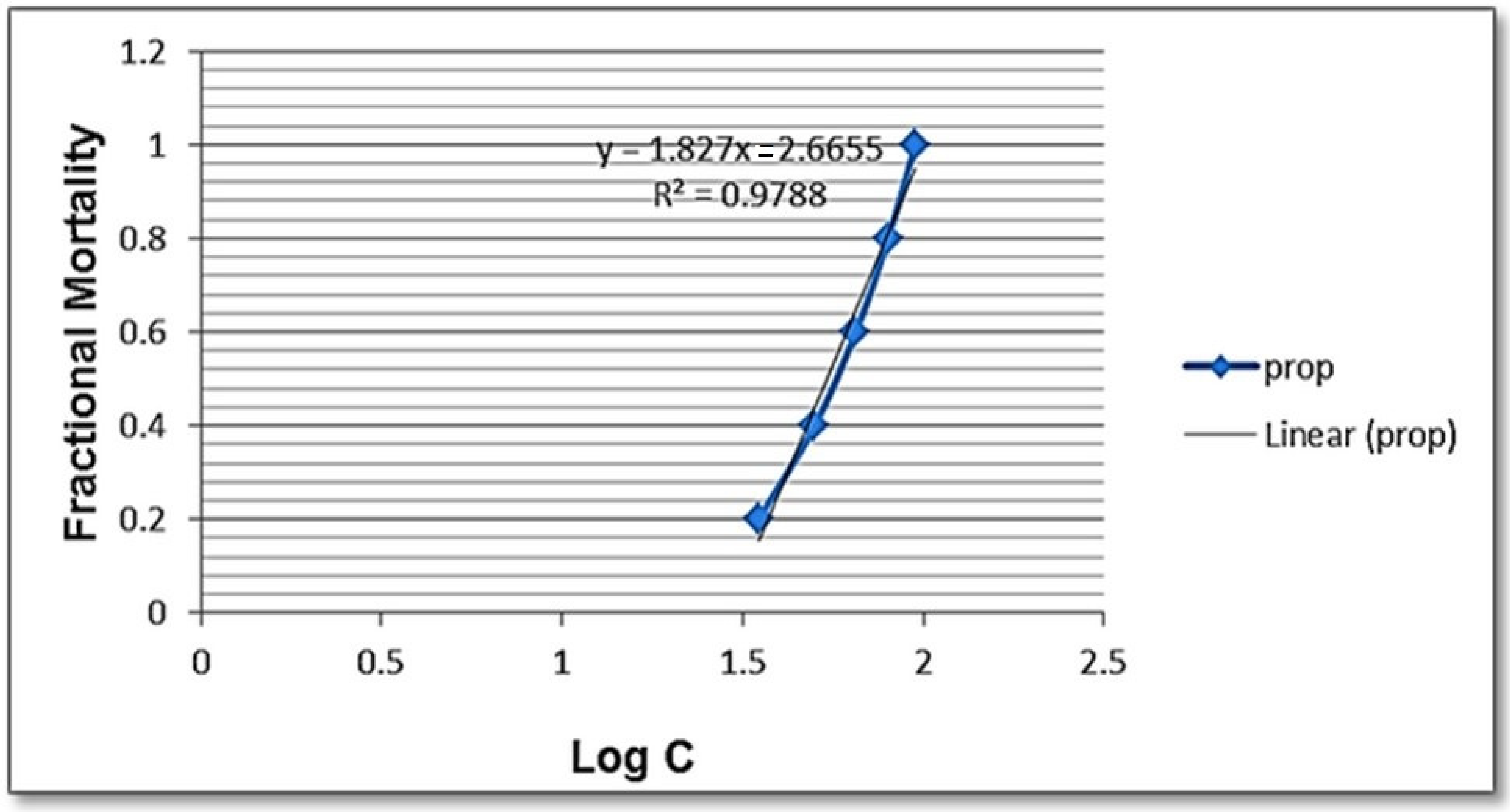
3.4. Effects of Selenium Nanoparticles against B. alexandrina Snails
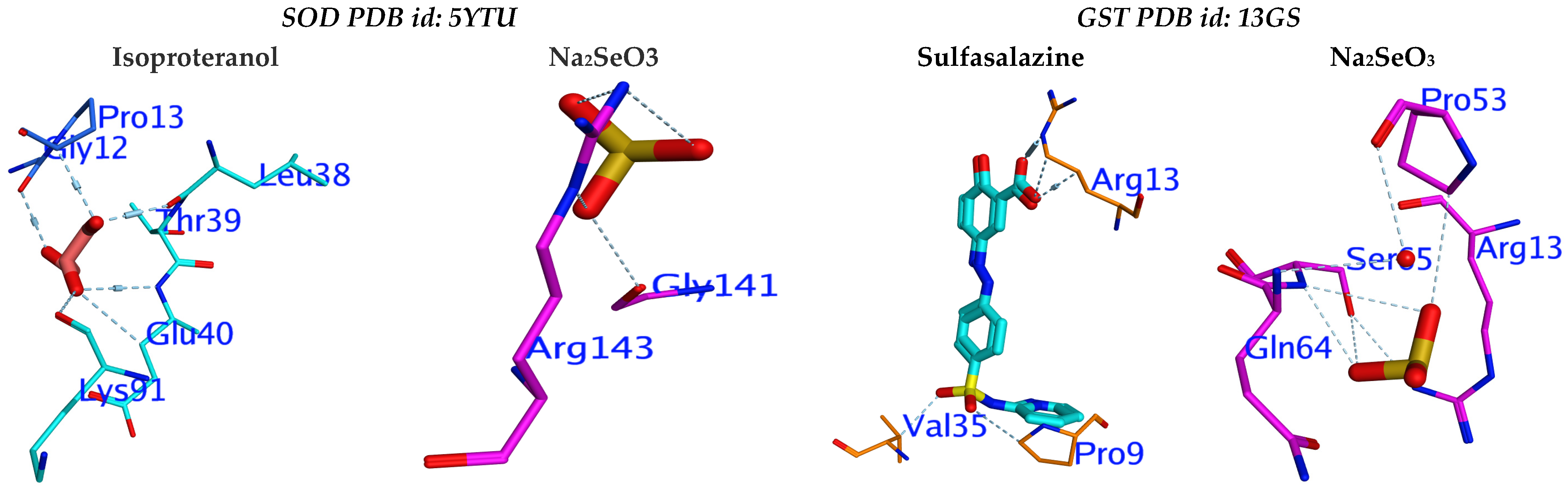
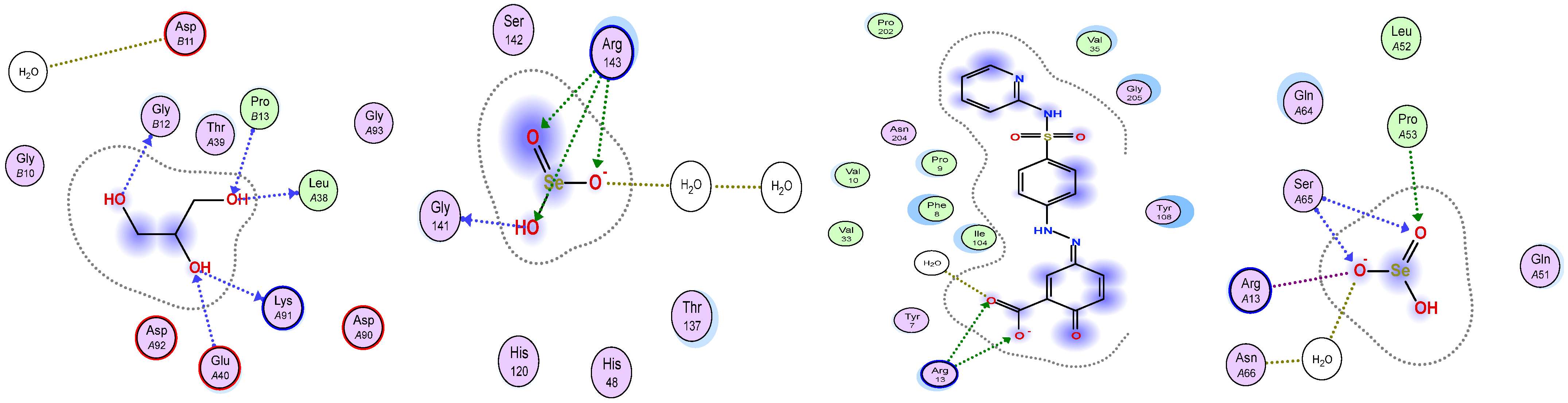
3.5. Molecular Docking Study
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO Schistosomiasis. 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/schistosomiasis (accessed on 11 February 2022).
- Ibrahim, A.M.; Sayed, S.S.M. Assessment of the molluscicidal activity of the methanolic seed extracts of Ziziphus spina-christi and Carica papaya on immunological and molecular aspects of Biomphalaria alexandrina snails. Aquac. Res. 2021, 52, 2014–2024. [Google Scholar] [CrossRef]
- Loker, E.S.; Mkoji, G.M. Schistosomes and Their Snail Hosts BT—Schistosomiasis. In Schistosomiasis; Secor, W.E., Colley, D.G., Eds.; Springer: Boston, MA, USA, 2005; pp. 1–11. ISBN 978-0-387-23362-8. [Google Scholar]
- Faust, C.L.; Crotti, M.; Moses, A.; Oguttu, D.; Wamboko, A.; Adriko, M.; Adekanle, E.K.; Kabatereine, N.; Tukahebwa, E.M.; Norton, A.J.; et al. Two-year longitudinal survey reveals high genetic diversity of Schistosoma mansoni with adult worms surviving praziquantel treatment at the start of mass drug administration in Uganda. Parasites Vectors 2019, 12, 607. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Tawab, H.; Ibrahim, A.M.; Hussein, T.; Mohamed, F. Mechanism of action and toxicological evaluation of engineered layered double hydroxide nanomaterials in Biomphalaria alexandrina snails. Environ. Sci. Pollut. Res. 2021, 29, 11765–11779. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.M.; Bakry, F.A. Assessment of the molluscicidal impact of extracted chlorophyllin on some biochemical parameters in the nervous tissue and histological changes in Biomphalaria alexandrina and Lymnaea natalensis snails. Invertebr. Neurosci. 2019, 19, 7. [Google Scholar] [CrossRef] [PubMed]
- Elsareh, F.; Abdalla, R.; Abdalla, E. The effect of aqueous leaves extract of Solenostemma argel (Del Hayne) on egg masses and neonates of Biomphalaria pfeifferi snails. J. Med. Plants 2016, 4, 271–274. [Google Scholar]
- Abd El Ghaffar, M.M.; Sadek, G.S.; Harba, N.M.; Abd El Samee, M.F. Evaluation of the effect of some plant molluscicides on the infectivity of Schistosoma mansoni cercariae. Menoufia Med. J. 2018, 31, 1448–1455. [Google Scholar] [CrossRef]
- Rani, P.; Kaur, G.; Rao, K.V.; Singh, J.; Rawat, M. Impact of Green Synthesized Metal Oxide Nanoparticles on Seed Germination and Seedling Growth of Vigna radiata (Mung Bean) and Cajanus cajan (Red Gram). J. Inorg. Organomet. Polym. Mater. 2020, 30, 4053–4062. [Google Scholar] [CrossRef]
- Fouda, A.; Abdel-Maksoud, G.; Abdel-Rahman, M.A.; Salem, S.S.; Hassan, S.E.D.; El-Sadany, M.A.H. Eco-friendly approach utilizing green synthesized nanoparticles for paper conservation against microbes involved in biodeterioration of archaeological manuscript. Int. Biodeterior. Biodegrad. 2019, 142, 160–169. [Google Scholar] [CrossRef]
- Pham, D.T.N.; Khan, F.; Phan, T.T.V.; Park, S.-k.; Manivasagan, P.; Oh, J.; Kim, Y.M. Biofilm inhibition, modulation of virulence and motility properties by FeOOH nanoparticle in Pseudomonas aeruginosa. Braz. J. Microbiol. 2019, 50, 791–805. [Google Scholar] [CrossRef]
- Raliya, R.; Tarafdar, J.C.; Choudhary, K.; Mal, P.; Raturi, A.; Gautam, R.K.S.; Singh, S.K. Synthesis of MgO Nanoparticles Using Aspergillus tubingensis TFR-3. J. Bionanosci. 2014, 8, 34–38. [Google Scholar] [CrossRef]
- Fatimah, I. Biosynthesis and characterization of ZnO nanoparticles using rice bran extract as low-cost templating agent. J. Eng. Sci. Technol. 2018, 13, 409–420. [Google Scholar]
- El-Belely, E.F.; Farag, M.M.S.; Said, H.A.; Amin, A.S.; Azab, E.; Gobouri, A.A.; Fouda, A. Green synthesis of zinc oxide nanoparticles (Zno-nps) using Arthrospira platensis (class: Cyanophyceae) and evaluation of their biomedical activities. Nanomaterials 2021, 11, 95. [Google Scholar] [CrossRef] [PubMed]
- Lashin, I.; Fouda, A.; Gobouri, A.A.; Azab, E.; Mohammedsaleh, Z.M.; Makharita, R.R. Antimicrobial and in vitro cytotoxic efficacy of biogenic silver nanoparticles (Ag-nps) fabricated by callus extract of Solanum incanum L. Biomolecules 2021, 11, 341. [Google Scholar] [CrossRef]
- Velgosova, O.; Dolinská, S.; Mražíková, A.; Briančin, J. Effect of P. kessleri extracts treatment on AgNPs synthesis. Inorg. Nano-Met. Chem. 2020, 50, 842–852. [Google Scholar] [CrossRef]
- Spagnoletti, F.N.; Spedalieri, C.; Kronberg, F.; Giacometti, R. Extracellular biosynthesis of bactericidal Ag/AgCl nanoparticles for crop protection using the fungus Macrophomina phaseolina. J. Environ. Manag. 2019, 231, 457–466. [Google Scholar] [CrossRef]
- Shafiq, T.; Uzair, M.; Iqbal, M.J.; Zafar, M.; Hussain, S.J.; Shah, S.A.A. Green synthesis of metallic nanoparticles and their potential in bio-medical applications. Nano Biomed. Eng. 2021, 13, 191–206. [Google Scholar] [CrossRef]
- Fouda, A.; Abdel-Maksoud, G.; Saad, H.A.; Gobouri, A.A.; Mohammedsaleh, Z.M.; El-Sadany, M.A.H. The efficacy of silver nitrate (AgNO3) as a coating agent to protect paper against high deteriorating microbes. Catalysts 2021, 11, 310. [Google Scholar] [CrossRef]
- Fouda, A.; Awad, M.A.; Eid, A.M.; Saied, E.; Barghoth, M.G.; Hamza, M.F.; Awad, M.F.; Abdelbary, S.; Hassan, S.E.D. An eco-friendly approach to the control of pathogenic microbes and Anopheles stephensi malarial vector using magnesium oxide nanoparticles (Mg-nps) fabricated by Penicillium chrysogenum. Int. J. Mol. Sci. 2021, 22, 5096. [Google Scholar] [CrossRef]
- Khalil, A.M.A.; Hassan, S.E.D.; Alsharif, S.M.; Eid, A.M.; Ewais, E.E.D.; Azab, E.; Gobouri, A.A.; Elkelish, A.; Fouda, A. Isolation and characterization of fungal endophytes isolated from medicinal plant Ephedra pachyclada as plant growth-promoting. Biomolecules 2021, 11, 140. [Google Scholar] [CrossRef]
- Rahman, A.U.; Wei, Y.; Ahmad, A.; Khan, A.U.; Ali, R.; Ullah, S.; Ahmad, W.; Yuan, Q. Selenium Nanorods Decorated Gold Nanostructures: Synthesis, Characterization and Biological Applications. J. Clust. Sci. 2020, 31, 727–737. [Google Scholar] [CrossRef]
- Geoffrion, L.D.; Hesabizadeh, T.; Medina-Cruz, D.; Kusper, M.; Taylor, P.; Vernet-Crua, A.; Chen, J.; Ajo, A.; Webster, T.J.; Guisbiers, G. Naked Selenium Nanoparticles for Antibacterial and Anticancer Treatments. ACS Omega 2020, 5, 2660–2669. [Google Scholar] [CrossRef] [PubMed]
- Filipović, N.; Ušjak, D.; Milenković, M.T.; Zheng, K.; Liverani, L.; Boccaccini, A.R.; Stevanović, M.M. Comparative Study of the Antimicrobial Activity of Selenium Nanoparticles With Different Surface Chemistry and Structure. Front. Bioeng. Biotechnol. 2021, 8, 624621. [Google Scholar] [CrossRef] [PubMed]
- Vahdati, M.; Tohidi Moghadam, T. Synthesis and Characterization of Selenium Nanoparticles-Lysozyme Nanohybrid System with Synergistic Antibacterial Properties. Sci. Rep. 2020, 10, 510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, W.; Yu, Z.; Lin, Z.; Lei, Z.; Ning, Z.; Regenstein, J.M.; Yang, J.; Ren, J. Biofunctionalization of Selenium Nanoparticle with Dictyophora indusiata Polysaccharide and Its Antiproliferative Activity through Death-Receptor and Mitochondria-Mediated Apoptotic Pathways. Sci. Rep. 2016, 5, 18629. [Google Scholar] [CrossRef]
- Menon, S.; Agarwal, H.; Venkat Kumar, S.; Rajeshkumar, S. Chapter 8—Biomemetic synthesis of selenium nanoparticles and its biomedical applications. In Green Synthesis, Characterization and Applications of Nanoparticles; Micro and Nano Technologies; Shukla, A.K., Iravani, S., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 165–197. ISBN 978-0-08-102579-6. [Google Scholar]
- Klaine, S.J.; Alvarez, P.J.J.; Batley, G.E.; Fernandes, T.F.; Handy, R.D.; Lyon, D.Y.; Mahendra, S.; McLaughlin, M.J.; Lead, J.R. Nanomaterials in the environment: Behavior, fate, bioavailability, and effects. Environ. Toxicol. Chem. 2008, 27, 1825–1851. [Google Scholar] [CrossRef] [PubMed]
- Pitt, J.I.; Hocking, A.D. Fungi and Food Spoilage; Springer: Berlin/Heidelberg, Germany, 2009; Volume 519. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Amin, M.A.; Ismail, M.A.; Badawy, A.A.; Awad, M.A.; Hamza, M.F.; Awad, M.F.; Fouda, A. The Potency of Fungal-Fabricated Selenium Nanoparticles to Improve the Growth Performance of Helianthus annuus L. and Control of Cutworm Agrotis ipsilon. Catalysts 2021, 11, 1551. [Google Scholar] [CrossRef]
- Vahidi, H.; Kobarfard, F.; Kosar, Z.; Mahjoub, M.A.; Saravanan, M.; Barabadi, H. Mycosynthesis and characterization of selenium nanoparticles using standard Penicillium chrysogenum PTCC 5031 and their antibacterial activity: A novel approach in microbial nanotechnology. Nanomed. J. 2020, 7, 315–323. [Google Scholar] [CrossRef]
- Salem, S.S.; Fouda, M.M.G.; Fouda, A.; Awad, M.A.; Al-Olayan, E.M.; Allam, A.A.; Shaheen, T.I. Antibacterial, Cytotoxicity and Larvicidal Activity of Green Synthesized Selenium Nanoparticles Using Penicillium corylophilum. J. Clust. Sci. 2021, 32, 351–361. [Google Scholar] [CrossRef]
- Eveland, L.K.; Haseeb, M.A. Laboratory Rearing of Biomphalaria glabrata Snails and Maintenance of Larval Schistosomes In Vivo and In Vitro. In Biomphalaria Snails and Larval Trematodes; Springer: New York, NY, USA, 2011; pp. 33–55. [Google Scholar]
- WHO. Report of the Scientific Working Group on Plant Molluscicide & Guidelines for Evaluation of Plant Molluscicides; TDR/SCHSWE (4)/83.3; WHO: Geneva, Switzerland, 1983. [Google Scholar]
- WHO. Molluscicide screening and evaluation. Bull. World Health Organ. 1965, 33, 567–581. [Google Scholar]
- Hamdi, S.A.H.; Ibrahim, A.M.; Ghareeb, M.A.; Fol, M.F. Chemical characterization, biocidal and molluscicidal activities of chitosan extracted from the crawfish Procambarus clarkii (Crustacea: Cambaridae). Egypt. J. Aquat. Biol. Fish. 2021, 25, 355–371. [Google Scholar] [CrossRef]
- Eissa, M.M.; El Bardicy, S.; Tadros, M. Bioactivity of miltefosine against aquatic stages of Schistosoma mansoni, Schistosoma haematobium and their snail hosts, supported by scanning electron microscopy. Parasit. Vectors 2011, 4, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nduku, W.K.; Harrison, A.D. Cationic responses of organs and haemolymph of Biomphalaria pfeifferi (Krauss), Biomphalaria glabrata (Say) and Helisoma trivolvis (Say)(Gastropoda: Planorbirdae) to cationic alterations of the medium. Hydrobiologia 1980, 68, 119–138. [Google Scholar] [CrossRef]
- Abdul-Salam, J.M.; Michelson, E.H. Biomphalaria glabrata amoebocytes: Effect of Schistosoma mansoni infection on in vitro phagocytosis. J. Invertebr. Pathol. 1980, 35, 241–248. [Google Scholar] [CrossRef]
- Grazeffe, V.S.; de Freitas Tallarico, L.; de Sa Pinheiro, A.; Kawano, T.; Suzuki, M.F.; Okazaki, K.; de Bragança Pereira, C.A.; Nakano, E. Establishment of the comet assay in the freshwater snail Biomphalaria glabrata (Say, 1818). Mutat. Res.—Genet. Toxicol. Environ. Mutagen. 2008, 654, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.M.; Sayed, D.A. Toxicological impact of oxyfluorfen 24% herbicide on the reproductive system, antioxidant enzymes, and endocrine disruption of Biomphalaria alexandrina (Ehrenberg, 1831) snails. Environ. Sci. Pollut. Res. Int. 2019, 26, 7960–7968. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Beutler, E.; Duron, O.; Kelly, B.M. Improved method for the determination of blood glutathione. J. Lab. Clin. Med. 1963, 61, 882–888. [Google Scholar]
- Koracevic, D.; Koracevic, G.; Djordjevic, V.; Andrejevic, S.; Cosic, V. Method for the measurement of antioxidant activity in human fluids. J. Clin. Pathol. 2001, 54, 356–361. [Google Scholar] [CrossRef] [Green Version]
- Bellos, J.K.; Perrea, D.N.; Theodotopoulou, E.; Vlachos, I.; Kostakis, A.I. Nitric oxide production as an indicator of recurrence of focal and segmental glomerulosclerosis following kidney transplantation. Saudi J. Kidney Dis. Transpl. 2011, 22, 1030–1032. [Google Scholar] [PubMed]
- Finney, D.J. Probit Analysis, 3rd ed.; Combridge University Press: New York, NY, USA, 1971. [Google Scholar]
- Murray, R. Speigel: Statistical estimation theory. In Schaum’s Outline Series: Theory and Problems of Statistics in SI Units, 1st ed.; McGraw Hill Book Company: Singapore, 1981; Volume 156. [Google Scholar]
- Oyeyemi, O.T. Application of nanotized formulation in the control of snail intermediate hosts of schistosomes. Acta Trop. 2021, 220, 105945. [Google Scholar] [CrossRef] [PubMed]
- Pohl, C.; Polli, F.; Schütze, T.; Viggiano, A.; Mózsik, L.; Jung, S.; de Vries, M.; Bovenberg, R.A.L.; Meyer, V.; Driessen, A.J.M. A Penicillium rubens platform strain for secondary metabolite production. Sci. Rep. 2020, 10, 7630. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.M.; De Britto, S.; Jogaiah, S.; Ito, S.I. Mycogenic selenium nanoparticles as potential new generation broad spectrum antifungal molecules. Biomolecules 2019, 9, 419. [Google Scholar] [CrossRef] [Green Version]
- Ramamurthy, C.H.; Sampath, K.S.; Arunkumar, P.; Kumar, M.S.; Sujatha, V.; Premkumar, K.; Thirunavukkarasu, C. Green synthesis and characterization of selenium nanoparticles and its augmented cytotoxicity with doxorubicin on cancer cells. Bioprocess Biosyst. Eng. 2013, 36, 1131–1139. [Google Scholar] [CrossRef]
- Worrall, E.A.; Hamid, A.; Mody, K.T.; Mitter, N.; Pappu, H.R. Nanotechnology for plant disease management. Agronomy 2018, 8, 285. [Google Scholar] [CrossRef] [Green Version]
- Zare, B.; Babaie, S.; Setayesh, N.; Shahverdi, A.R.; Shahverdi, A. Isolation and characterization of a fungus for extracellular synthesis of small selenium nanoparticles Extracellular synthesis of selenium nanoparticles using fungi. Nanomed. J. 2013, 1, 13–19. [Google Scholar]
- Ahmad, M.S.; Yasser, M.M.; Sholkamy, E.N.; Ali, A.M.; Mehanni, M.M. Anticancer activity of biostabilized selenium nanorods synthesized by Streptomyces bikiniensis strain Ess_amA-1. Int. J. Nanomed. 2015, 10, 3389–3401. [Google Scholar] [CrossRef] [Green Version]
- Ullah, A.; Yin, X.; Wang, F.; Xu, B.; Mirani, Z.A.; Xu, B.; Chan, M.W.H.; Ali, A.; Usman, M.; Ali, N.; et al. Biosynthesis of selenium nanoparticles (via Bacillus subtilis bsn313), and their isolation, characterization, and bioactivities. Molecules 2021, 26, 5559. [Google Scholar] [CrossRef]
- Ranjitha, V.R.; Ravishankar, V.R. Extracellular Synthesis of Selenium Nanoparticles from an Actinomycetes Streptomyces griseoruber and Evaluation of its Cytotoxicity on HT-29 Cell Line. Pharm. Nanotechnol. 2018, 6, 61–68. [Google Scholar] [CrossRef]
- Salem, S.S. Bio-fabrication of Selenium Nanoparticles Using Baker’s Yeast Extract and Its Antimicrobial Efficacy on Food Borne Pathogens. Appl. Biochem. Biotechnol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Barabadi, H.; Kobarfard, F.; Vahidi, H. Biosynthesis and Characterization of Biogenic Tellurium Nanoparticles by Using Penicillium chrysogenum PTCC 5031: A Novel Approach in Gold Biotechnology. Iran. J. Pharm. Res. IJPR 2018, 17, 87–97. [Google Scholar] [PubMed]
- Al-Brakati, A.; Alsharif, K.F.; Alzahrani, K.J.; Kabrah, S.; Al-Amer, O.; Oyouni, A.A.; Habotta, O.A.; Lokman, M.S.; Bauomy, A.A.; Kassab, R.B.; et al. Using green biosynthesized lycopene-coated selenium nanoparticles to rescue renal damage in glycerol-induced acute kidney injury in rats. Int. J. Nanomed. 2021, 16, 4335–4349. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, K.; Sandhya, M.V.S.; Koganti, S.; Burgula, S. Selenium nanoparticles synthesized using Pseudomonas stutzeri (Mh191156) show antiproliferative and anti-angiogenic activity against cervical cancer cells. Int. J. Nanomed. 2020, 15, 4523–4540. [Google Scholar] [CrossRef] [PubMed]
- Alagesan, V.; Venugopal, S. Green Synthesis of Selenium Nanoparticle Using Leaves Extract of Withania somnifera and Its Biological Applications and Photocatalytic Activities. Bionanoscience 2019, 9, 105–116. [Google Scholar] [CrossRef] [Green Version]
- Lian, S.; Diko, C.S.; Yan, Y.; Li, Z.; Zhang, H.; Ma, Q.; Qu, Y. Characterization of biogenic selenium nanoparticles derived from cell-free extracts of a novel yeast Magnusiomyces ingens. 3 Biotech 2019, 9, 221. [Google Scholar] [CrossRef]
- Osman, G.Y.; Ahmed, M.M.; Abdel Kader, A.; Mohamed, A.A. Biological and biochemical impacts of the fungal extract of Aspergillus fumigatus on Biomphalaria alexandrina snails infected with Schistosoma mansoni. Biosciences 2013, 7, 473–484. [Google Scholar]
- Mahmoudvand, H.; Shakibaie, M.; Tavakoli, R.; Jahanbakhsh, S.; Sharifi, I. In Vitro Study of Leishmanicidal Activity of Biogenic Selenium Nanoparticles against Iranian Isolate of Sensitive and Glucantime-Resistant Leishmania tropica. Iran. J. Parasitol. 2014, 9, 452–460. [Google Scholar]
- Alkhudhayri, A.; Al-Shaebi, E.M.; Qasem, M.A.A.; Murshed, M.; Mares, M.M.; Al-Quraishy, S.; Dkhil, M.A. Antioxidant and anti-apoptotic effects of selenium nanoparticles against murine eimeriosis. An. Acad. Bras. Cienc. 2020, 92, e20191107. [Google Scholar] [CrossRef]
- Dkhil, M.A.; Khalil, M.F.; Diab, M.S.M.; Bauomy, A.A.; Santourlidis, S.; Al-Shaebi, E.M.; Al-Quraishy, S. Evaluation of nanoselenium and nanogold activities against murine intestinal schistosomiasis. Saudi J. Biol. Sci. 2019, 26, 1468–1472. [Google Scholar] [CrossRef]
- De Carvalho Augusto, R.; Merad, N.; Rognon, A.; Gourbal, B.; Bertrand, C.; Djabou, N.; Duval, D. Molluscicidal and parasiticidal activities of Eryngium triquetrum essential oil on Schistosoma mansoni and its intermediate snail host Biomphalaria glabrata, a double impact. Parasites Vectors 2020, 13, 486. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.M.; Ahmed, A.K.; Bakry, F.A.; Abdel-Ghaffar, F. Hematological, physiological and genotoxicological effects of Match 5% EC insecticide on Biomphalaria alexandrina snails. Ecotoxicol. Environ. Saf. 2018, 147, 1017–1022. [Google Scholar] [CrossRef] [PubMed]
- Ali, D. Evaluation of environmental stress by comet assay on freshwater snail Lymnea luteola L. exposed to titanium dioxide nanoparticles. Toxicol. Environ. Chem. 2014, 96, 1185–1194. [Google Scholar] [CrossRef]
- Ibrahim, A.M.; Ghoname, S.I. Molluscicidal impacts of Anagallis arvensis aqueous extract on biological, hormonal, histological and molecular aspects of Biomphalaria alexandrina snails. Exp. Parasitol. 2018, 192, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, L.; Zhao, J.; Xing, B. Environmental processes and toxicity of metallic nanoparticles in aquatic systems as affected by natural organic matter. Environ. Sci. Nano 2016, 3, 240–255. [Google Scholar] [CrossRef]
- Prokhorova, E.E.; Serebryakova, M.K.; Tokmakova, A.S.; Ataev, G.L. Hemocytes of mollusc Biomphalaria glabrata (Gastropoda, pulmonata). Invertebr. Surviv. J. 2018, 15, 346–351. [Google Scholar] [CrossRef]
- Cavalcanti, M.G.S.; Filho, F.C.; Mendonça, A.M.B.; Duarte, G.R.; Barbosa, C.C.G.S.; De Castro, C.M.M.B.; Alves, L.C.; Brayner, F.A. Morphological characterization of hemocytes from Biomphalaria glabrata and Biomphalaria straminea. Micron 2012, 43, 285–291. [Google Scholar] [CrossRef]
- Abdel-Hamid, H.; Mekawey, A.A.I. Biological and hematological responses of Biomphalaria alexandrina to mycobiosynthsis silver nanoparticles. J. Egypt. Soc. Parasitol. 2014, 44, 627–637. [Google Scholar] [CrossRef]
- Ibrahim, A.M.; Ahmed, A.K.; Bakry, F.A.; Rabei, I.; Abdel-Ghaffar, F. Toxicological impact of butralin, glyphosate-isopropylammonium and pendimethalin herbicides on physiological parameters of Biomphalaria alexandrina snails. Molluscan Res. 2019, 39, 224–233. [Google Scholar] [CrossRef]
- Ray, M.; Bhunia, N.S.; Bhunia, A.S.; Ray, S. A comparative analyses of morphological variations, phagocytosis and generation of cytotoxic agents in flow cytometrically isolated hemocytes of Indian molluscs. Fish Shellfish Immunol. 2013, 34, 244–253. [Google Scholar] [CrossRef]
- Donaghy, L.; Hong, H.-K.; Lambert, C.; Park, H.-S.; Shim, W.J.; Choi, K.-S. First characterisation of the populations and immune-related activities of hemocytes from two edible gastropod species, the disk abalone, Haliotis discus discus and the spiny top shell, Turbo cornutus. Fish Shellfish Immunol. 2010, 28, 87–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalil, A.M. Toxicological effects and oxidative stress responses in freshwater snail, Lanistes carinatus, following exposure to chlorpyrifos. Ecotoxicol. Environ. Saf. 2015, 116, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Barky, F.A.; Abdelsalam, H.A.; Mahmoud, M.B.; Hamdi, S.A.H. Influence of Atrazine and Roundup pesticides on biochemical and molecular aspects of Biomphalaria alexandrina snails. Pestic. Biochem. Physiol. 2012, 104, 9–18. [Google Scholar] [CrossRef]
- Sayed, S.S.M.; Abdel-Wareth, M.T.A. The comparative effect of chlorine and Huwa-san as disinfecting agents on Biomphalaria alexandrina snails and free larval stages of Schistosoma mansoni. Parasitol. Res. 2017, 116, 2627–2635. [Google Scholar] [CrossRef]
- Wang, W.; Mao, Q.; Yao, J.; Yang, W.; Zhang, Q.; Lu, W.; Deng, Z.; Duan, L. Discovery of the pyridylphenylureas as novel molluscicides against the invasive snail Biomphalaria straminea, intermediate host of Schistosoma mansoni. Parasites Vectors 2018, 11, 291. [Google Scholar] [CrossRef]
- Saleh, H.A.; Abdel-Motleb, A.; Habib, M.R. Neurotoxicity and genotoxicity of the veterinary antibiotics oxytetracycline and trimethoprim-sulphadiazine to Biomphalaria alexandrina snails. Int. J. Environ. Stud. 2021, 78, 983–1002. [Google Scholar] [CrossRef]
- Chemical Computing Group Inc. Molecular operating environment (MOE). 2016. Available online: https://www.chemcomp.com/Products.htm (accessed on 10 February 2022).
- Manjula, R.; Wright, G.S.A.; Strange, R.W.; Padmanabhan, B. Assessment of ligand binding at a site relevant to SOD1 oxidation and aggregation. FEBS Lett. 2018, 592, 1725–1737. [Google Scholar] [CrossRef] [Green Version]
- Oakley, A.J.; Lo Bello, M.; Nuccetelli, M.; Mazzetti, A.P.; Parker, M.W. The ligandin (non-substrate) binding site of human Pi class glutathione transferase is located in the electrophile binding site (H-site). J. Mol. Biol. 1999, 291, 913–926. [Google Scholar] [CrossRef]
- Kim, G.H.; Kim, J.E.; Rhie, S.J.; Yoon, S. The Role of Oxidative Stress in Neurodegenerative Diseases. Exp. Neurobiol. 2015, 24, 325–340. [Google Scholar] [CrossRef]
- Bastos, F.F.; Tobar, S.A.L.; Dantas, R.F.; Silva, E.S.; Nogueira, N.P.A.; Paes, M.C.; Righi, B.D.P.; Bastos, J.C.; Bastos, V.L.F.C. Melatonin affects conjugation of 4-hydroxynonenal with glutathione in liver of pacu, a hypoxia-tolerant fish. Fish Physiol. Biochem. 2013, 39, 1205–1214. [Google Scholar] [CrossRef]
- Montassir, L.; Berrebaan, I.; Mellouki, F.; Zkhiri, F.; Boughribil, S.; Bessi, H. Acute toxicity and reprotoxicity of aqueous extract of a Moroccan plant (Tetraclinis articulata) on freshwater cladoceran Daphnia magna. J. Mater. Environ. Sci. 2017, 8, 770–776. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lethal Concentration Doses | LC10 | LC25 | LC50 | LC90 | Slope |
|---|---|---|---|---|---|
| Concentration (mg/L) | 31.826 | 44.15 | 57.85 | 83.87 | 1.4 |
| Comet % | Tail Length (px) | % DNA in Tail | Tail Moment | |
|---|---|---|---|---|
| Control | 14.95 | 8.15 ± 0.42 | 23.79 ± 3.75 | 1.955 ± 0.402 |
| LC10 | 15.85 | 10.507 ± 0.54 * | 33.66 ± 1.43 * | 3.529 ± 0.03 * |
| LC25 | 19.5 | 11.96 ± 1.37 * | 33.588 ± 5.7 * | 4.094 ± 1.15 * |
| Groups | Testosterone (nmol/L) | Estradiol (pg/mL) |
|---|---|---|
| Control | 20 ± 0.52 | 100 ± 3.1 |
| LC10 | 30 ± 0.71 * | 300 ± 5.2 * |
| LC25 | 35 ± 0.56 * | 1000 ± 4.6 * |
| Biochemical Parameters | MDA (nmol/g.tissue) | NO (µmol/L) | SOD (U/g.tissue) | GST (U/g.tissue) | TAC (mM/L) |
|---|---|---|---|---|---|
| Control | 9.811 ± 0.012 | 136.75 ± 0.42 | 8.36 ± 0.1 | 0.85 ± 0.05 | 1.505 ± 0.4 |
| LC10 | 10.612 ± 0.4 * | 148 ± 0.34 * | 5.045 ± 0.02 * | 0.927 ± 0.1 | 1.488 ± 0.21 |
| LC25 | 14.47 ± 0.3 * | 322 ± 0.21 * | 3.624 ± 0.2 * | 1.0545 ± 0.03 * | 1.375 ± 0.3 |
| ΔG | rmsd | E.vdw | E.Int | E.H.B. | Eele | |
|---|---|---|---|---|---|---|
| 5YTU | −5.03 | 3.86 | −266.94 | −1.86 | −11.10 | −27.82 |
| 13GS | −4.70 | 1.53 | −266.52 | −2.18 | −10.09 | −27.31 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morad, M.Y.; El-Sayed, H.; Elhenawy, A.A.; Korany, S.M.; Aloufi, A.S.; Ibrahim, A.M. Myco-Synthesized Molluscicidal and Larvicidal Selenium Nanoparticles: A New Strategy to Control Biomphalaria alexandrina Snails and Larvae of Schistosoma mansoni with an In Silico Study on Induced Oxidative Stress. J. Fungi 2022, 8, 262. https://doi.org/10.3390/jof8030262
Morad MY, El-Sayed H, Elhenawy AA, Korany SM, Aloufi AS, Ibrahim AM. Myco-Synthesized Molluscicidal and Larvicidal Selenium Nanoparticles: A New Strategy to Control Biomphalaria alexandrina Snails and Larvae of Schistosoma mansoni with an In Silico Study on Induced Oxidative Stress. Journal of Fungi. 2022; 8(3):262. https://doi.org/10.3390/jof8030262
Chicago/Turabian StyleMorad, Mostafa Y., Heba El-Sayed, Ahmed A. Elhenawy, Shereen M. Korany, Abeer S. Aloufi, and Amina M. Ibrahim. 2022. "Myco-Synthesized Molluscicidal and Larvicidal Selenium Nanoparticles: A New Strategy to Control Biomphalaria alexandrina Snails and Larvae of Schistosoma mansoni with an In Silico Study on Induced Oxidative Stress" Journal of Fungi 8, no. 3: 262. https://doi.org/10.3390/jof8030262