1. Introduction
Maize was globally the most important cereal produced in 2019 and 2020 [
1]. Maize is commonly affected by three main toxigenic fungi:
F. graminearum, producer of deoxynivalenol and zearalenone,
F. verticillioides, synthetizing more than hundred fumonisins (where FUM B
1 and B
2 have the highest importance), and
A. flavus producing aflatoxins, the most important of which is AFB1. During warm and humid seasons,
F. graminearum is more commonly found in maize; warmer and rather dry conditions are favorable to
F. verticillioides; and
A. flavus is common in the warmest corn areas [
2,
3,
4]. The ecological needs of these fungi are different; therefore, they can occur alone or combined. In regions with crossroads of different climatic influences, such as Hungary, all three may be present with their toxins in one year. The disease resistance and toxin resistance of various hybrids might be different [
5]. Reliable information on maize resistance to toxigenic fungi has seldom been provided for commercial hybrids [
6]; therefore, an evaluation of the production risks of hybrids for food safety is a global but mostly neglected task. In the commercial hybrids, no inbred lines can be tested. Therefore, genetic work is not possible. There is no reason to test inbreds in this respect, as none are used in commercial production. To influence commercial production, information about the ear rot resistance and food safety risks of the hybrids is needed to choose the less susceptible ones. An improved understanding of the ear rot resistance, the toxin relations, the artificial and natural infection comparisons, and toxin data with respect to different toxigenic fungi may help to provide farmers more reliable information on the risk of hybrids. These data are similarly important for farmers, breeders, animal husbandry, and the ethanol industry.
The economic losses due to toxigenic fungi are substantial [
7], with corn loss estimated at ranging from 52 million to 1.68 billion USD annually in the U.S. as climate change further progresses, and one review [
8] lists many other literature sources on the global toxin situation. In Hungary, the 2014 mycotoxin contamination caused 330 million USD in damage to corn production and animal husbandry [
5]. At least 10% of the global harvested grains is toxin-contaminated above limit values. Adopting this number for maize as well (about 100 MMT) is a problem that requires a more rapid treatment than traditional plant breeding can suggest [
8].
There are at least 19
Fusarium species from Europe [
9], and they produce a substantial number of mycotoxins. Two of them have global significance:
F. graminearum, which produces deoxynivalenol (DON) and zearalenone (ZEA), as well as
F. verticillioides, which synthesizes the members of the fumonisin group [
10]. In Hungary, similar
Fusarium species occur [
11] with similar significance.
Aspergillus spp. have occurred in Hungary decades before, but the high rate of
Penicillium spp. proved that samples originated from stored samples [
12]. However, the preharvest origin of the aflatoxin could not be verified until now.
A. flavus has been accepted as a preharvest aflatoxin producer in the U.S. [
10] (postharvest contamination also remains significant). Knowing this, we started testing for Aspergillus resistance, as we supposed that Hungary would eventually face the same problem when summer temperatures increase. When preharvest AFB1 is above 20 μg/kg, preventive methods with a higher resistance, supported by agronomy and other means, are crucial [
13,
14]. Toxin data from SGS Company [
15] showed that DON, fumonisins, and AFB1 have been regular components of corn grains every year since 2012, with large local differences. As the data (except the low rates) originated from mixtures from different fields and hybrids [
16], the data could not be connected to individual hybrids. In the U.S., a significant part of AFB1 is of field origin [
10]. Therefore, the inclusion of
A. flavus in the resistance test was a logical step to prepare for the preharvest identification of
A. flavus resistance and aflatoxin screening in Hungary.
The global climate models forecasting significant temperature increases [
17,
18] reported that AFB1 contamination would increase with a 2 °C increase in South Germany throughout the full Carpathian Basin. Other authors reached similar conclusions [
19,
20].
The AFB1 production of
A. flavus in maize is significantly influenced by the false positive isolates with full cluster for AFB1 synthesis but without actual toxin production ability. In Kenya two maize populations were screened for toxic rate of
A. flavus isolates: 71% and 62% of the isolates produced aflatoxin on coconut agar [
21]. In India 63% and 52% of the
A. flavus isolates were AFB1 producer in maize grains [
22]. In our lab, 42
A. flavus isolates produced AFB1 in rice medium with the presence of the whole aflatoxin gene cluster, but only eight produced AFB1 on maize ears. Thirty-four can be classified as false positive (Toth 2019, unpublished). To differentiate the false positive isolates from the positive a quintuplex RT-PCR procedure was suggested that could differentiate the false positive isolates from the real toxin producer ones [
23].
Magan et al. [
24] analyzed DON for wheat, fumonisins, and AFB1 in maize and found that increasing warmth and dryer weather will favor fumonisins and aflatoxins in maize, and warm temperatures with excessive rain will increase DON in wheat. This also applies to DON contamination in maize [
15,
25].
Areas such as Mediterranean Region are today at high risk for aflatoxins and fumonisins because the temperature is now optimum or higher than optimum for fungal infection and toxin production. This is not yet the case in Middle and Western Europe, but a higher toxin pressure was forecasted [
24]. A systemic infection at high temperatures by
F. verticillioides [
26] increases additional risk for FER infections.
The probability of infection mediated by rachilla is possible with all three pathogens [
27,
28,
29,
30]. The conclusion is that regions below the optimum for diseases or toxins will be more exposed in warmer seasons. The severely hit Mediterranean regions will remain in this category [
18] and need help. We know that the moisture content of the cob is significantly higher, which allows the fungus to grow on the surface of the cob and infect the grain and germs, which cannot be detected from the surface [
31]. Epidemiologically, this is a problem. As in earlier hybrids, the drydown of the ear and cob is closer, and their use can decrease this additional risk, as seen in Hungary in 2014.
The literature distinguishes two resistance types for ear rots [
32]: kernel resistance and silk channel resistance. Kernel resistance (KR) is tested by growing fungus on toothpicks or dipping toothpicks or steel needles in a suspension and then inoculating in the middle of the ear. Silk channel resistance (SR) is tested by injecting a given amount of suspension into the silk channel, and the infection will be mediated by the silks to the grains. Normally, there are significant correlations between the responses of the genotypes and the two types [
25,
33,
34,
35,
36,
37]. In the Hungarian dryer and warmer conditions, kernel ear rot severity is threefold higher than the silk channel data, providing an improved differentiation of the 44 genotypes [
25]. KR provided a much clearer differentiation of the genotypes. Therefore, kernel resistance was chosen for standard tests, even though it seems to be a well-supported fact that most ear inoculations are silk mediated [
10].
Resistance relationships vary between toxigenic species. Most breeding programs have concentrated on a single toxigenic species, mostly
F. verticillioides, and much less on
F. graminearum. Several authors have reported significant correlations between resistance to
F. graminearum and
F. verticillioides [
38,
39,
40,
41]. There are examples of correlated results between
F. verticillioides and
A. flavus resistance or
F. graminearum and
F. verticillioides, but no data exist for the three pathogens in a single research program. Löffler et al. [
40] found genotype/environment interactions between GER and FER, but this was never extended to all tested genotypes. The conclusion of Rose et al. [
13] on the relation between
F. verticillioides and
A. flavus was the same. In recently tested hybrids [
5,
25], 10–15% showed similar resistance to the three pathogens, and 5–10% showed high susceptibility to all; and 75–80% of the hybrids showed a highly variable resistance to the different pathogens. In several hybrids, a strong toxin overproduction has been found, e.g., a much higher toxin contamination than would follow from visual ear rot severity [
5,
25]. There are several papers describing medium to close relationships between resistance to
F. verticillioides and
A. flavus, and some FER-resistant maize inbred lines can be a source of
A. flavus and AFB1 resistance [
41,
42]. Other authors have identified AFB1 and fumonisin resistance (the resistance to disease and resistance to toxin accumulation were treated as synonyms) in two new inbred lines in a mapping population by indicating the lack of general agreement [
43,
44]. Therefore, there is likely no general agreement on the resistance to these pathogens. The conclusion is that resistance to different toxigenic species should be tested separately. A general ear rot resistance does not exist.
Significant correlations were found between the severity of visual symptoms and toxin contamination; generally, they were of medium closeness or were somewhat stronger [
45,
46,
47,
48]. Bolduan et al. [
49] calculated an unusually close correlation between ear rot and DON (r = 0.94) with respect to Fusarium ear rot (FER); the conclusion was that toxin analysis may not be necessary. In other cases, much lower correlations were found [
3]. Correlation-breaking genotypes were also identified, indicating that a general agreement in maize would not be the case. Löffler et al. [
35] analyzed the relationship between toxin contamination and visual ear rot infection severity in maize inbreds to
F. graminearum and
F. verticillioides. They found that separate divergent behavior is not an exception. For this reason, parallel testing of resistance to both pathogens were suggested. A strong agreement was found between
F. graminearum and
F. culmorum [
5,
50,
51]. Other data also support this view [
5,
25,
50].
The reduction in disease symptoms due to resistance has seldom been discussed, and even less so for toxins. Focker et al. [
52] reported a reduction in AFB1 of between 62% and 82%. Whether we can reduce toxin contamination below the official limit is another question. We think that the reduction rate is not sufficient, and that the contamination level should be below the EU limits.
In terms of the methodology, except in the pathogenicity tests performed by Mesterházy et al. [
4], all authors have worked with one isolate or a mixture of isolates, which has remained unchanged until now [
25]. Based on initial experimental results on wheat, maize hybrids inoculated with different independent isolates have been evaluated [
50,
51,
53]. In the first experiment [
50], 14 isolates (10
F. graminearum and 4
F. culmorum) were assessed on 10 hybrids. Of the 55 possible correlations in the hybrid reactions, 22 were not significant, and Hybrid 2 did not have any significant relationships with the reactions to other hybrids. Hybrid 10 showed significant correlations with all other hybrids, except Hybrid 2. In another test, four isolates were applied independently without mixing [
54]. The severity of the disease and the toxin contamination were highly variable. The differences between years were also significant, which is no surprise. Other results [
55] also support the view that more isolates are necessary to obtain a more reliable picture of resistance and toxin relationships.
In risk analysis, different philosophies can be applied. Traditional natural infection indicates that highly infected genotypes can be excluded; however, in Hungary, such severity occurs once or twice in a decade. As different years might show differing fungal species compositions, screening for complex resistance on this basis is impossible [
11]. For natural toxin contamination, we used EU limit values directly [
56,
57,
58,
59].
The conclusion is that a high rate of commercial hybrids is highly susceptible to one or more major toxigenic species. However, a smaller but significant part of the hybrids performs well with
Fusarium spp. [
5,
25]. This is clear proof that commercial hybrid programs do not pay enough attention to the screening of hybrids for breeding resistance to toxigenic species and to inhibit toxin contamination. As the aflatoxin response could not be measured earlier [
5], we aimed to include it into the testing program. Strategically, the most important step is the screening of the hybrids, as preference of more resistant hybrids can lead to a rapid improvement in food safety. The extent to which this is possible with three toxigenic species and three largely differing toxins is the question we seek to answer. Screening of the inbreds will be necessary, but their combining abilities to toxigenic species and yield are probably different. Therefore, their complex value can be decided better in their hybrids.
Our objectives were as follows:
Analyzing the relationships between symptom severity and toxin contamination and to analyze the behavior of hybrids under different ecological conditions to further understand natural infection and toxin contamination; determining how the risk of a given hybrid can be evaluated, which traits should be considered, and how they should be weighted in the analysis and evaluating the influence of the growing resistance level to better tolerate the higher amplitudes of the changing climate, which causes mostly higher temperatures and draught and is favorable to fumonisin and aflatoxin production.
2. Materials and Methods
2.1. Plant Material and Experimental Design
Maize hybrids were chosen from the registered hybrids in Hungary, from Corteva Agriscience (Pioneer, Dow AgroSciences Hungary Kft. 2040 Budaörs, Neumann János str. 1., Hungary), Bayer (DeKalb 1117 Budapest, Dombóvári str. 26., Hungary), RAGT (Budaörs, story 3, Keleti str. 7, 2040, Hungary), Syngenta (Budaörs, story 3, Keleti str. 7, 2040, Hungary), etc. In the project, 23 hybrids were tested. Therefore, we agreed that, in the first two years, 18 will be identical, with a possibility to include five new hybrids in the second year. The same was true for the second two years. However, four controls were tested across the four years. Trials conducted for this study were part of the Trials System of Hungarian Maize Club Association (a public organization), which aims to improve the knowledge of farmers.
The experiments were conducted in Kiszombor, 25 km east of Szeged, in the Maros River Valley (GPS coordinates: 46°12′49.0″ N and 20°09′57.9″ E). This is a research station of Cereal Research Ltd. Yearly precipitation varies between 350 and 1100 mm, and soil pH is 6.98. The latest soil humus content was 2.21%, which has been decreasing for a long time; the NO3-N is 5.8 mg/kg, classified as extremely low; the P2O5 is 280 mg/kg, the K2O is 317 mg/kg, the Mg is 376 mg/kg (all three are at high rate), Zn and SO4 are poor, and the rest are moderate. Therefore, in autumn, 160 kg of Genesis (8:21:21% NPP rate) was administered, and 80 kg of Nitrosol (46% carbamide) was administered in the spring. Both are products of Péti Nitrokomplex Ltd. (8105 Pét, Hősök square 14, Hungary). Irrigation was performed (40 mm per treatment) when necessary, just after sowing (between 25 April and 3 May) in mid-June to enhance ear differentiation and at the end of the inoculation time in the third week of July (or somewhat later depending on the weather). To control the European corn borer, 0.2 L/ha Decis (Bayer Inc. Leverkusen, Germany a.i. deltamethrin 50 g/L) was used (1 or 2 treatments). For weed control, Lumax from Syngenta (5732 Mezőtúr, Hungary a.i. 37.5 g/L mesotrione, 375.0 g/L S-metolachlor, and 125.0 g/L terbutaline) was applied at rate 4.5 L/ha, Dezormon from Nufarm Hungaria Ltd. (1118 Budapest Hungary, a.i. 600 g/L, 2,4-D), and Shadow 200 from BASF (BASF Hungaria Ltd., 1132 Budapest, Vaci str. 92, Hungary, a.i. 200 g/L dimethenamid-P, 200 g/L metazachlor, and 100 g/L quinmerac) at a dose of 2.5 L/ha were used, depending on the availability and weed composition.
Each plot consisted of four rows 8 m long. At half distance, a 50-cm-wide road was cut out, and the length was dissected into two 3.75-m-long parts. In the lower part of the row, Isolate 1 was used, and the upper part was inoculated by Isolate 2. The isolates for the three rows were:
F. graminearum (No. 3 and 4),
F. verticillioides (No. 1 and 2), and
Aspergillus flavus (No. 1 and 2). The fourth row was the control without artificial inoculation to allow us to observe the background ear rot severity and toxin contamination (
Table 1). Three replicates (all with four rows) were organized in a randomized block design. Due to the use of two independent isolates, for each hybrid, we had four independent biological replicates in the two years; for the control hybrids, we had eight. This amount of data is sufficient for drawing reliable conclusions.
2.2. Isolates and Inoculation
Two isolates were used for all three fungal species. For
F. graminearum isolates, No. 3 and 4 were used; for
F. verticillioides, No. 1 and 8 were used; for
A. flavus, No. 1 and 2 were used. All were isolated from naturally infected grains. A PCR-based method was used to identify
A. flavus isolates. Here, a region of the calmodulin gene of the fungal DNA was amplified using the primers cmd5 and cmd6, as described by Hong et al. [
60]. For
F. graminearum, the IGS-RFLP method was used [
61]. For
F. verticillioides, the method of Baird et al. [
62] was followed by testing for the presence of the FUM1 gene from the fumonisin gene cluster. For DNA extraction, the methodology of the cited papers was followed [
60,
61,
62]. The strains were deposited in the Microbe Gene Bank of Cereal Research Nonprofit Ltd., which is part of the Hungarian National Centre for Plant Diversity and is freely accessible. Their deposit numbers are as follows: Fg3: NGBAB142629; Fg4: NGBAB142696; Fv1: NGBAB142625; Fv2: NGBAB142624; Af1: NGBAB142601; Af2: NGBAB142602. The nucleotide sequences for species determination were as follows:
F. graminearum and
F. verticillioides: EF1-α primers: ef1: ATGGGTAAGGARGACAAGAC; ef2: GGARGTACCAGTSATCATGTT;
A. flavus: CaM primers: CMD5: CCGAGTACAAGGARGCCTTC; CMD6: CCGATRGAGGTCATRACGTGG. For inoculation, the toothpick method developed by Young [
63] and modified by Mesterházy [
35] was used measuring kernel resistance. Wood toothpicks were washed three timed to be free of tannins and other fungal growth inhibitors. Once air dried, toothpicks were placed into glass flasks and supplied with a liquid Czapek-Dox medium for an hour. After removal of the medium, except for 5 mm at the bottom of the flask, the mouth of the glass was closed by a cotton cork. They were autoclaved at 120 °C for an hour. After cooling in a sterile inoculation box, 2–3 inocula were transmitted to the wet toothpicks. The toothpicks were ready to use in three weeks stored at room temperature without direct sunshine. As no suspension was used, no conidium concentration could be measured. The aggressiveness of the isolates was tested in previous years on maize ears [
5,
25]. Silk channel inoculation, which mostly works under cooler and more humid conditions [
49], has failed in Hungary [
25]. Inoculation was performed for all three pathogens and their isolates 6 days after 50% silking by inserting infested toothpicks into the middle of the upper ear in a hole made by an awl 15-mm-long and 1.5-mm-wide. As most hybrids in Hungary belong to the FAO 200–300 and early 400, and the dry and mostly hot temperature speeds up plant development, we had to make the KR inoculation 6 days after midsilking. Normally, 15–18 ears in a row were inoculated. There was only one inoculation time per row, and late flowering ears were discarded. The toothpicks were left in the ears until harvest, so that the inoculated ears could be identified.
2.3. Evaluation of Symptoms and Risks
For all three pathogens, the same percent scale was used [
5,
25]. Its origin is the scale suggested by Reid et al. [
32], but it should have been modified to differentiate the ear rot severities more clearly for toxin comparison. In a regular ear, 700–800 grains grow. At 1% infection, 7–8 grains show visible infection. When only one grain is infected, the rate is about 0.15%. Rates from above 5% to 100% were considered on a scale by 5% steps. For evaluation, only ears where the mark of the toothpick could be identified were considered. This was to secure improved sampling. This was important because, at higher resistance or lower aggressiveness (such as
A. flavus and
F. verticillioides), some ears were not infected, but the trace of the toothpick signaled that the inoculation was performed. All ears were assigned two numbers: the first was the percentage of the visually detectable grains whose infection seemingly originated from the infested toothpick; the other was assigned to grains seemingly infected independently of the toothpick. This latter was considered an additional natural infection. The insect-wounded and Fusarium-infected ears were not considered to avoid mixing artificial infection with the insect damage caused Fusarium and toxin contamination. For correct sampling, this was an important precondition. No such description was found in the literature in our research.
For the control role, the two hybrids chosen were not perfectly suitable, as these reacted differently to diseases and toxins. Therefore, we related the performance of the hybrids to the arithmetical mean of the 18 hybrids. In the tables the genotypes (low risk) that were lower than 50% of the experimental mean are highlighted in dark green. The hybrids (from a low to medium risk) between 51% and the experimental mean (100%) are marked in light green. By adding 50% of the experimental mean to this mean, the medium- to high-risk hybrids are grouped and highlighted in yellow. All other hybrids were classified as high risk, highlighted in orange. In the naturally infected materials, the toxin limit agrees with the limits of EC regulations [
56,
57,
58,
59].
2.4. Preparing Samples for Toxin Analyses
For toxin analyses, we followed our own procedure [
25]. Briefly, five maize ears without insect damage and average ear rot values were selected. After an evaluation of ear symptoms (within 24 h), they were placed into a plastic string web Rashel bag and kept in a dry room until dry. Following shelling, the whole amount about of 1 kg was roughly ground to 1–2-mm particles, which were thoroughly mixed. This was conducted separately for all replicates. Afterwards, from each replicate, 100 g were separated, pooled, and mixed again to decrease the sampling error to the lowest possible. From this roughly mixed material, 100 g were separated for toxin analysis and sent to Nagyigmand, an accredited laboratory. As such, the infected particles could be better homogenized for toxin distribution than any other methods used for whole grains. This was useful for
F. graminearum, but even more so for
F. verticillioides and
A. flavus, for which the infection severity of 1% or lower was rather characteristic and therefore much more critical.
2.5. Toxin Analysis
For the analysis, the methodology of the accredited Bonafarm Feed Laboratory Nagyigmand, Hu., was used [
64]. All the reagents and solvents were bought from Thomasker. The solvents and glacial acetic acid (AcOH) were bought from Honeywell™. Ultrapure water was made freshly every day using a Human Corp. Zeneer Power I (Human Building 36, Garak-ro, Songpa-gu, Seoul, Republic of Korea, Postal Code 05694) water purification system. Mycotoxin reference standards were supplied by Romer Labs
® (Romer Labs Inc., 130 Sandy Drive Newark, DE 19713, USA).
For the preparation of standards, briefly, for AFB1, the RomerBiopureTM Mix 5 was diluted five times to obtain a stock solution. For the toxin analysis of fumonisins, RomerBiopureTM Mix 3 was applied. To produce the stock solutions, a 60-fold dilution was applied. The RomerBioPurTM DON was diluted 66-fold to prepare the stock solution.
For sample preparation, the maize samples were ground using a Perten Laboratory mill (Type: 3310, Perten Instruments, 126 53 Hagersten, Sweden). Five grams from each sample were separated and placed into 50-mL PP centrifuge tubes. This consisted of 0.5–1 g of the subsamples that were collected from different parts of the 100 g sample. Afterwards, 40 mL of an acetonitrile/water (AcOH, 20:79:1, v/v/v) solution was added to AFB1 B1 and fumonisin B1+B2. For DON, ultrapure water was used for extraction. The mixture was vortexed and extracted by a rotary shaker for 1.5 h at 200 shakes/minute. Following centrifugation at 5000 rpm for 15 min, the supernatant was passed through a PTFE filter (0.2 mm pore size).
For chromatography, a Kinetex® C18 100 Å UPLC column (2.1 × 100 mm, 1.7 μm) was used and kept at 30 °C. The flow rate was 0.35-mL min-1, and a 5-µL partial loop injection was used (a loop of 20 µL). Mobile phases were buffered with 5 mM of ammonium-acetate. Ultrapure water with 1% AcOH and 4% MeOH was used as Mobile Phase A, and MeOH containing 1% AcOH with 2% ultrapure water was used as Mobile Phase B.
UPLC-MS/MS was performed in MRM mode on an AB Sciex QTRAP® 6500 tandem mass spectrometer connected to a 1290 Agilent Infinity II-UPLC system (Agilent Technologies, 5301 Stevens Creek Blvd. Santa Clara, CA 95051 USA) equipped with Agilent components as follows: a G1316C Thermostatted Column Compartment, a G1330B Thermostat, a G4220A Binary Pump, and a G4226A Sampler.
The main mass spectrometer worked at a source temperature of 350 °C, a curtain gas of 40 psig, an ion source gas of 35 psig, a detector voltage of 5 kV, and an entrance potential of 10 V. The collision gas was set to medium. MS/MS conditions were optimized in the Analyst
®1.6.2 compound optimization module using the direct infusions of each analyte’s standard. The detailed parameters of each analyte were optimized, as summarized in
Table 2. FB
1+B
2 and AFB1 were measured positively. For DON, the negative mode was applied.
For calculations, all the sample solutions were measured against a calibration curve for the analyte of interest. Raw results were calculated using the Analyst® 1.6.2 software package. The peak concentration was calculated as cspike = cstockˑ(Vspike÷Vfinal). The results were corrected with the recovery of the standard addition, i.e., the weighed sample, and the extraction solvent was added using the following formula: csample = (crawˑVextr ÷ msample)/((crawspiked/craw) ÷ cspike).
2.6. Statistical Methods
For the artificial ear rot data, the four- and two-way ANOVA models were used in three replicates. For the toxin data, the two-way ANOVA model was used without replicates from the Excel Analytical Tools. As such, the year and isolate effects could be balanced to some extent. For stability, the variance from the one-way ANOVA (Excel) was used. Additionally, regression and correlation tests were used to compare ear rot and toxin data using the built-in Excel program. For the four-way ANOVA, first a two-way ANOVA was made by Excel from the sums of replicates, and the four-way ANOVA was then conducted by the functions presented by Sváb [
65] and Weber [
66]. Their statistical tables were also used to evaluate significance levels. The calculation of the overproduction of toxins is demonstrated in Figure 1, following the procedure of Mesterhazy et al. [
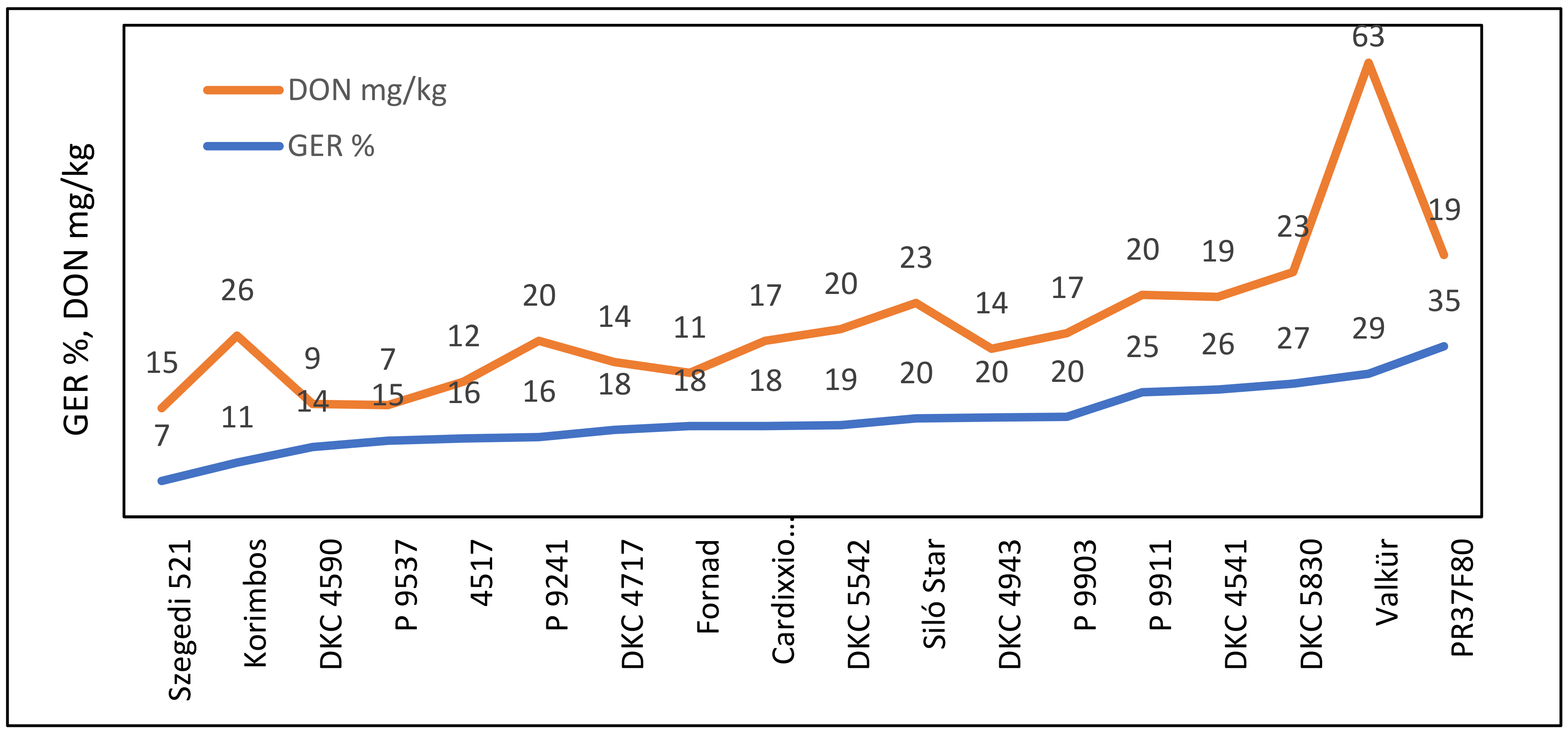
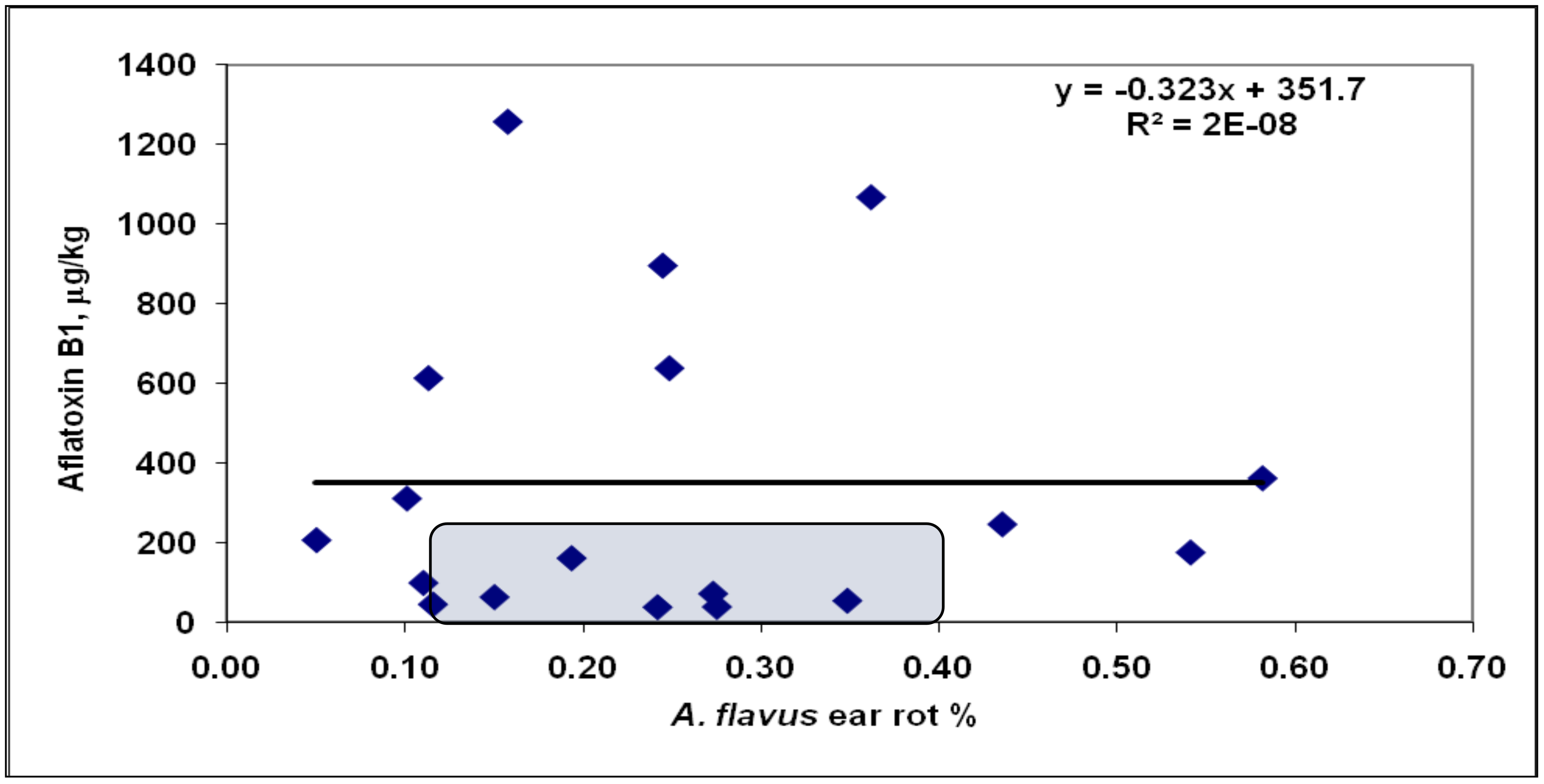
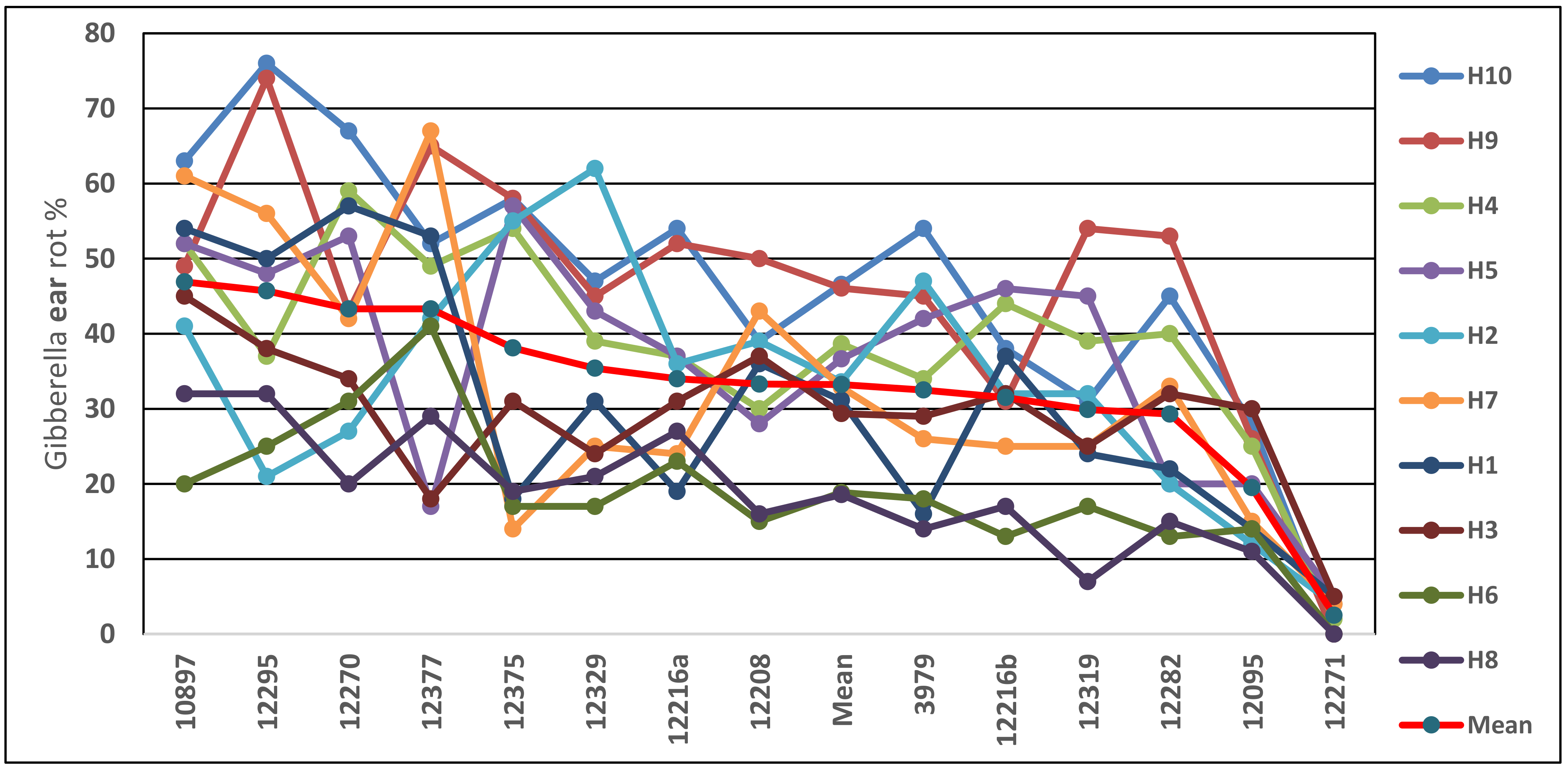
25]. This was followed for all further figures of this type.


 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}