
Heat Shock Transcription Factor CgHSF1 Is Required for Melanin Biosynthesis, Appressorium Formation, and Pathogenicity in Colletotrichum gloeosporioides


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Plant Material
2.2. Bioinformatics Analysis
2.3. Construction of the Gene Knock-Out Strain, PniiA-Cghsf1 Inducible Strain, and the GFP or FLAG Tagged Strains
2.4. Pathogenicity Assay
2.5. Quantitative RT-PCR Analysis
2.6. Appressorium Formation Assay
2.7. Melanin Content Measurement
2.8. Chromatin Immunoprecipitation (ChIP) and Quantitative PCR Analysis
2.9. Recombinant Protein Preparation and Electrophoretic Mobility Shift Assay (EMSA)
2.10. Statistical Analysis
3. Results
3.1. Identification and Bioinformatic Analysis of CgHSF1
3.2. Subcellular Localization of CgHSF1
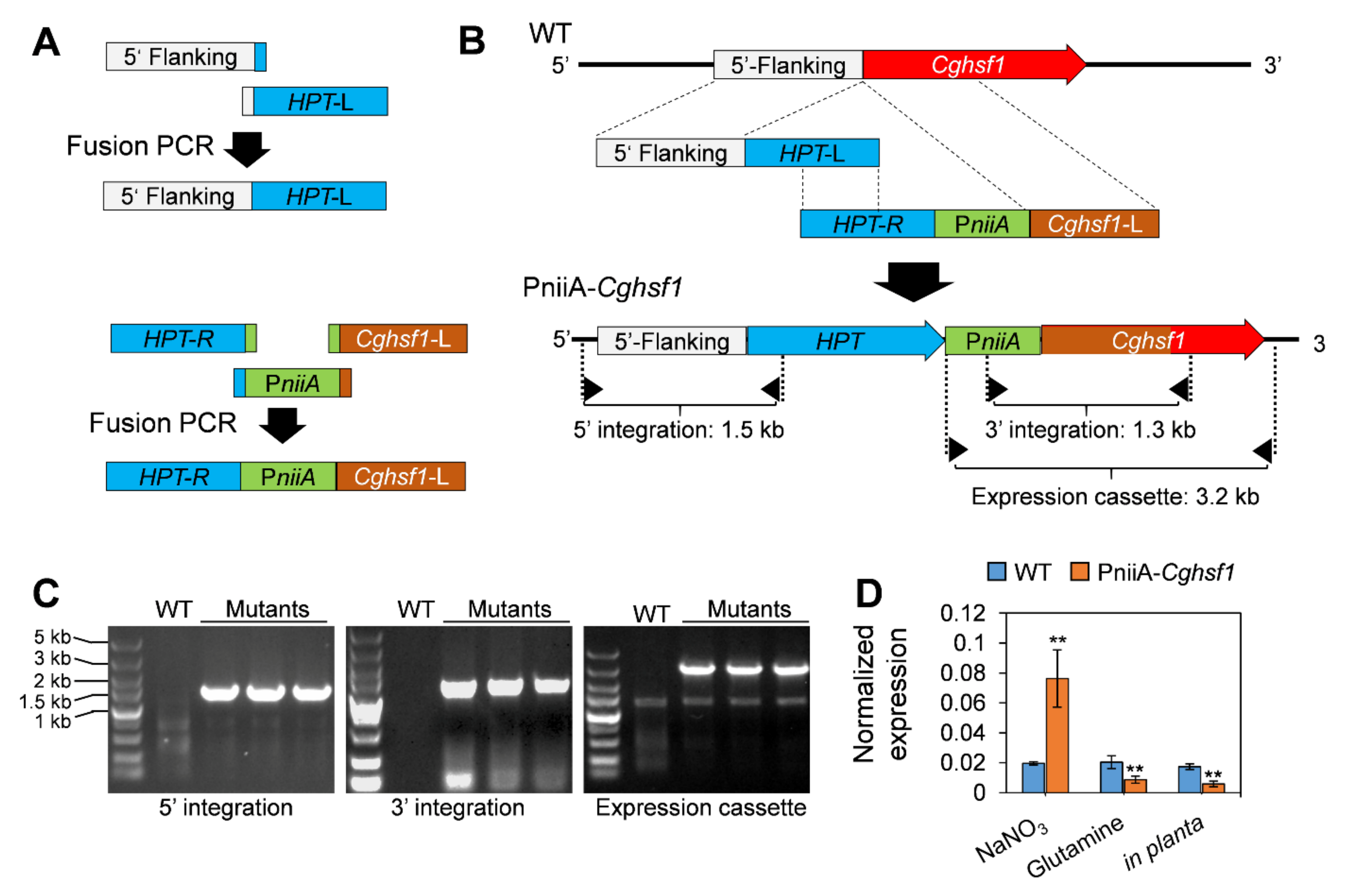
3.3. Generation of the PniiA-Cghsf1 Strain
3.4. CgHSF1 Is Required for Pathogenicity
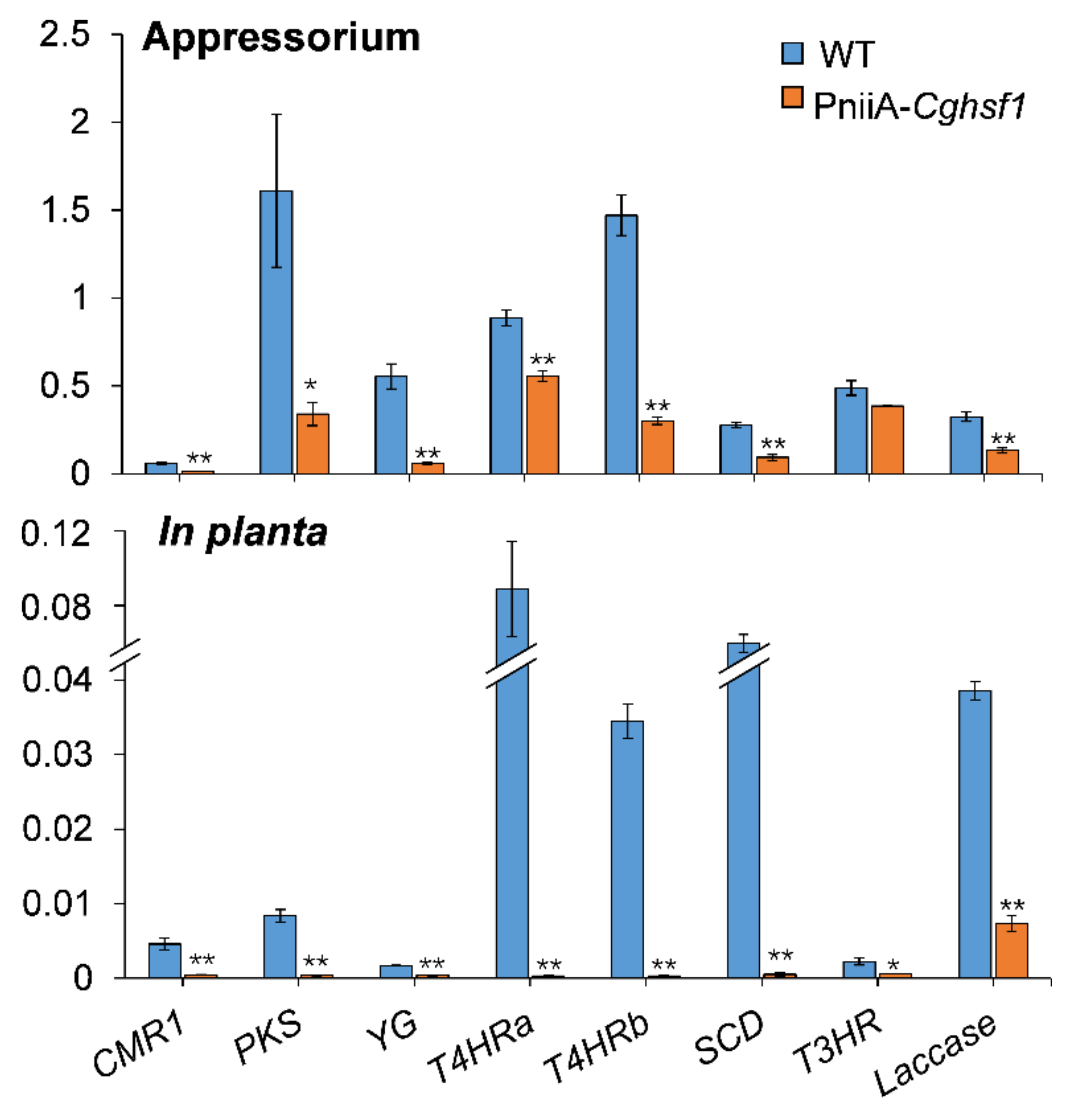
3.5. CgHSF1 Plays a Role in Appressorium Formation
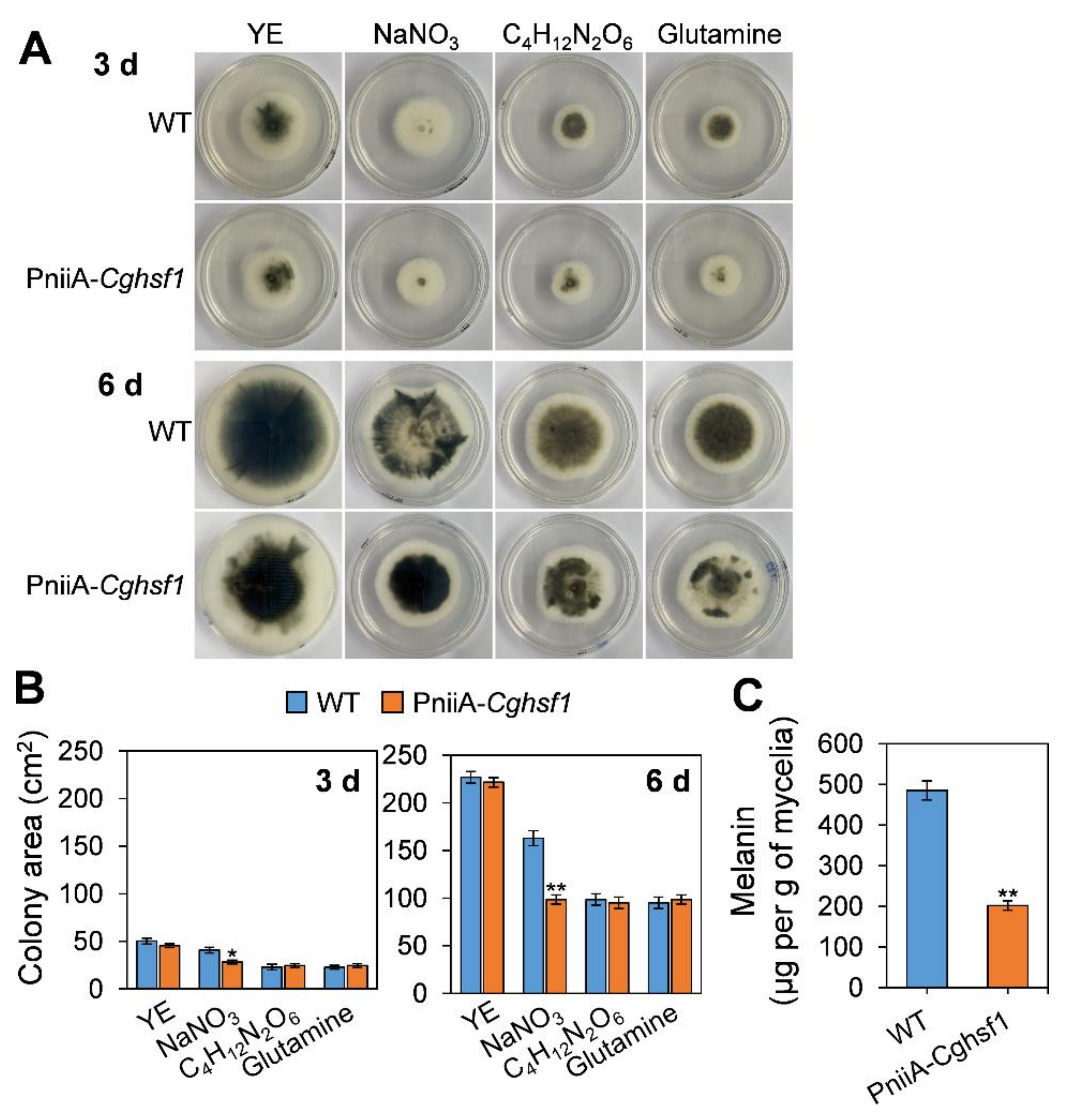
3.6. CgHSF1 Plays a Role in Melanin Biosynthesis
3.7. CgHSF1 Regulates Transcription of Melanin Biosynthesis Genes
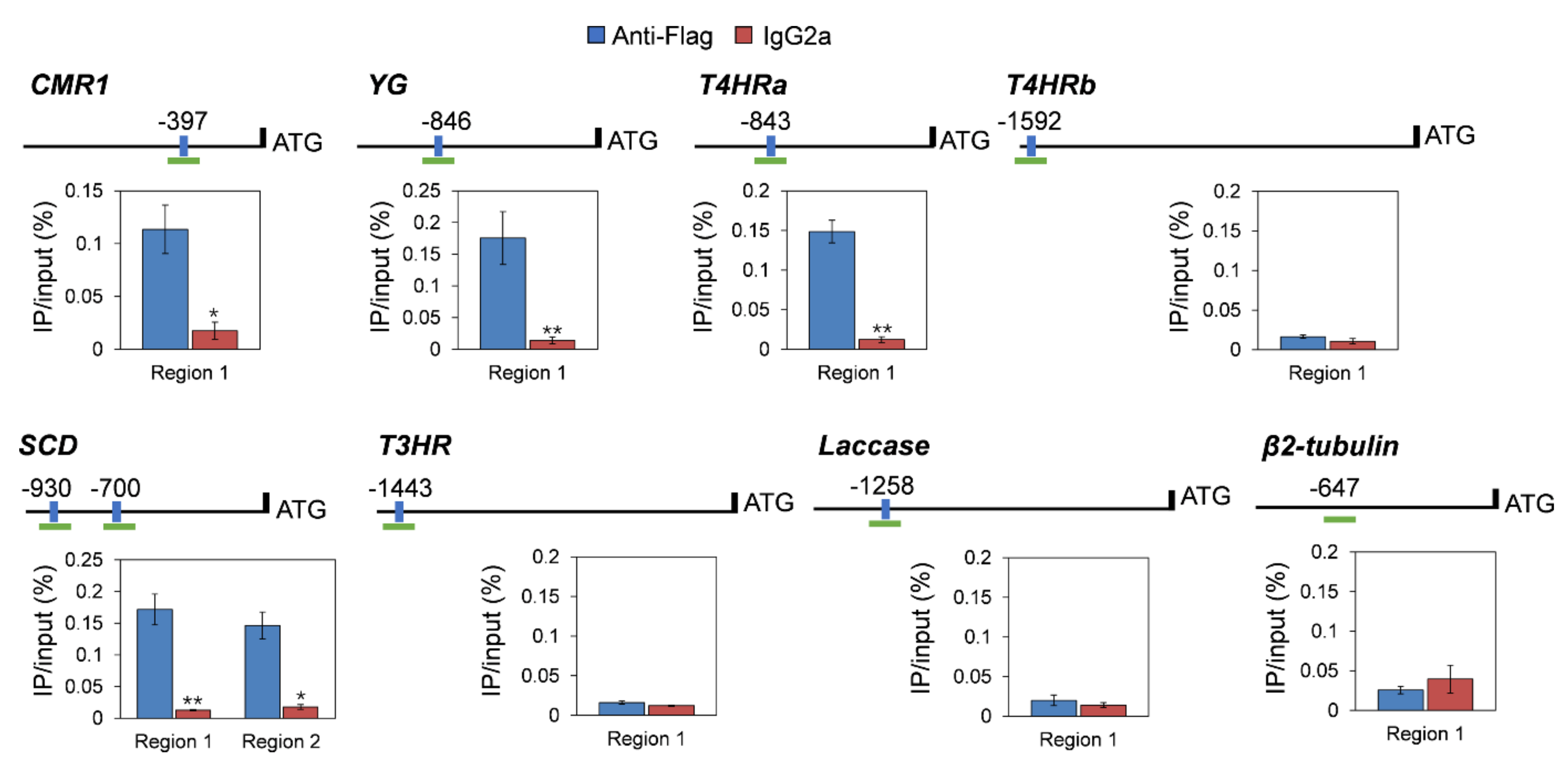
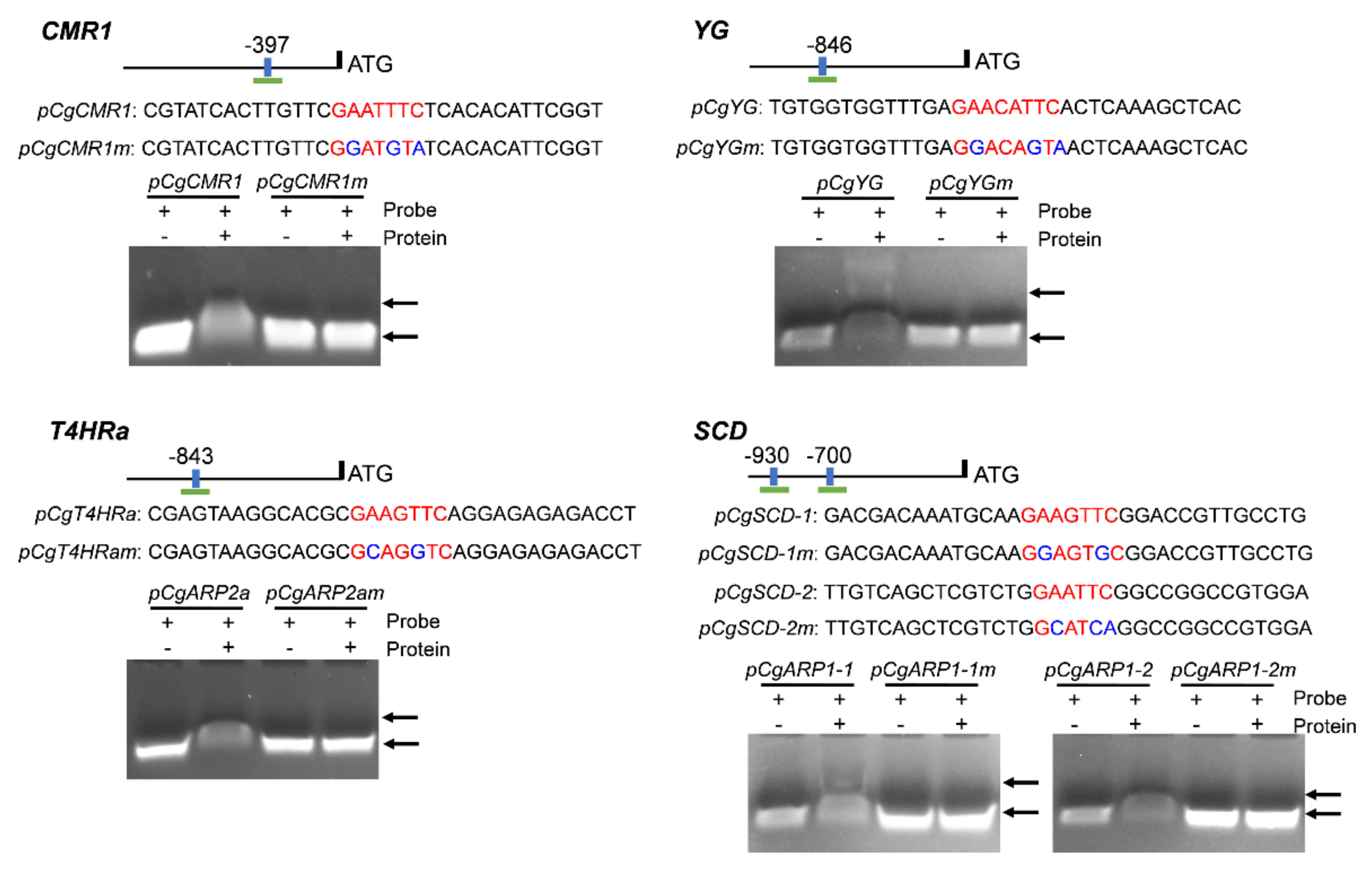
3.8. CgHSF1 Directly Binds to the Promoters of Melanin Biosynthesis Genes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 Fungal Pathogens in Molecular Plant Pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phoulivong, S.; Cai, L.; Chen, H.; McKenzie, E.H.; Abdelsalam, K.; Chukeatirote, E.; Hyde, K.D. Colletotrichum gloeosporioides Is Not a Common Pathogen on Tropical Fruits. Fungal Divers. 2010, 44, 33–43. [Google Scholar] [CrossRef]
- O’Connell, R.J.; Thon, M.R.; Hacquard, S.; Amyotte, S.G.; Kleemann, J.; Torres, M.F.; Damm, U.; Buiate, E.A.; Epstein, L.; Alkan, N.; et al. Lifestyle Transitions in Plant Pathogenic Colletotrichum Fungi Deciphered by Genome and Transcriptome Analyses. Nat. Genet. 2012, 44, 1060–1065. [Google Scholar] [CrossRef] [PubMed]
- Gan, P.; Ikeda, K.; Irieda, H.; Narusaka, M.; O'Connell, R.J.; Narusaka, Y.; Takano, Y.; Kubo, Y.; Shirasu, K. Comparative Genomic and Transcriptomic Analyses Reveal the Hemibiotrophic Stage Shift of Colletotrichum Fungi. New Phytol. 2013, 197, 1236–1249. [Google Scholar] [CrossRef]
- Huang, G.; Wang, H.; Chou, S.; Nie, X.; Chen, J.; Liu, H. Bistable Expression of WOR1, a Master Regulator of White-Opaque Switching in Candida albicans. Proc. Natl. Acad. Sci. USA 2006, 103, 12813–12818. [Google Scholar] [CrossRef] [Green Version]
- Cain, C.W.; Lohse, M.B.; Homann, O.R.; Sil, A.; Johnson, A.D. A Conserved Transcriptional Regulator Governs Fungal Morphology in Widely Diverged Species. Genetics 2012, 190, 511–521. [Google Scholar] [CrossRef] [Green Version]
- Jonkers, W.; Dong, Y.; Broz, K.; Kistler, H.C. The Wor1-Like Protein Fgp1 Regulates Pathogenicity, Toxin Synthesis and Reproduction in the Phytopathogenic Fungus Fusarium graminearum. PLoS Pathog. 2012, 8, e1002724. [Google Scholar] [CrossRef] [Green Version]
- Brown, D.W.; Busman, M.; Proctor, R.H. Fusarium verticillioides SGE1 Is Required for Full Virulence and Regulates Expression of Protein Effector and Secondary Metabolite Biosynthetic Genes. Mol. Plant Microbe Interact. 2014, 27, 809–823. [Google Scholar] [CrossRef] [Green Version]
- Michielse, C.B.; Studt, L.; Janevska, S.; Sieber, C.M.; Arndt, B.; Espino, J.J.; Humpf, H.U.; Güldener, U.; Tudzynski, B. The Global Regulator FfSge1 Is Required for Expression of Secondary Metabolite Gene Clusters but Not for Pathogenicity in Fusarium fujikuroi. Environ. Microbiol. 2015, 17, 2690–2708. [Google Scholar] [CrossRef]
- Van der Does, H.C.; Fokkens, L.; Yang, A.; Schmidt, S.M.; Langereis, L.; Lukasiewicz, J.M.; Hughes, T.R.; Rep, M. Transcription Factors Encoded on Core and Accessory Chromosomes of Fusarium oxysporum Induce Expression of Effector Genes. PLoS Genet. 2016, 12, e1006401. [Google Scholar] [CrossRef] [Green Version]
- Michielse, C.B.; van Wijk, R.; Reijnen, L.; Manders, E.M.; Boas, S.; Olivain, C.; Alabouvette, C.; Rep, M. The Nuclear Protein Sge1 of Fusarium oxysporum Is Required for Parasitic Growth. PLoS Pathog. 2009, 5, e1000637. [Google Scholar] [CrossRef] [Green Version]
- Michielse, C.B.; Becker, M.; Heller, J.; Moraga, J.; Collado, I.G.; Tudzynski, P. The Botrytis cinerea Reg1 Protein, a Putative Transcriptional Regulator, Is Required for Pathogenicity, Conidiogenesis, and the Production of Secondary Metabolites. Mol. Plant Microbe Interact. 2011, 24, 1074–1085. [Google Scholar] [CrossRef] [Green Version]
- Santhanam, P.; Thomma, B.P. Verticillium dahliae Sge1 Differentially Regulates Expression of Candidate Effector Genes. Mol. Plant Microbe Interact. 2013, 26, 249–256. [Google Scholar] [CrossRef] [Green Version]
- Mirzadi Gohari, A.; Mehrabi, R.; Robert, O.; Ince, I.A.; Boeren, S.; Schuster, M.; Steinberg, G.; de Wit, P.J.; Kema, G.H. Molecular characterization and functional analyses of ZtWor1, a transcriptional regulator of the fungal wheat pathogen Zymoseptoria tritici. Mol. Plant Pathol. 2014, 15, 394–405. [Google Scholar] [CrossRef]
- Tollot, M.; Assmann, D.; Becker, C.; Altmüller, J.; Dutheil, J.Y.; Wegner, C.E.; Kahmann, R. The WOPR Protein Ros1 Is a Master Regulator of Sporogenesis and Late Effector Gene Expression in the Maize Pathogen Ustilago maydis. PLoS Pathog. 2016, 12, e1005697. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zhai, S.; Zhang, H.; Zuo, R.; Wang, J.; Guo, M.; Zheng, X.; Wang, P.; Zhang, Z. Shared and Distinct Functions of Two Gti1/Pac2 Family Proteins in Growth, Morphogenesis and Pathogenicity of Magnaporthe oryzae. Environ. Microbiol. 2014, 16, 788–801. [Google Scholar] [CrossRef]
- Zahiri, A.; Heimel, K.; Wahl, R.; Rath, M.; Kämper, J. The Ustilago maydis Forkhead Transcription Factor Fox1 Is Involved in the Regulation of Genes Required for the Attenuation of Plant Defenses during Pathogenic Development. Mol. Plant Microbe Interact. 2010, 23, 1118–1129. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Kief, J.; Auffarth, K.; Farfsing, J.W.; Mahlert, M.; Nieto, F.; Basse, C.W. The Ustilago maydis Cys2His2-Type Zinc Finger Transcription Factor Mzr1 Regulates Fungal Gene Expression during the Biotrophic Growth stage. Mol. Microbiol. 2008, 68, 1450–1470. [Google Scholar] [CrossRef]
- Akerfelt, M.; Morimoto, R.I.; Sistonen, L. Heat Shock Factors: Integrators of Cell Stress, Development and Lifespan. Nat. Rev. Mol. Cell Biol. 2010, 11, 545–555. [Google Scholar] [CrossRef]
- Anckar, J.; Sistonen, L. Regulation of HSF1 Function in the Heat Stress Response: Implications in Aging and Disease. Annu. Rev. Biochem. 2011, 80, 1089–1115. [Google Scholar] [CrossRef]
- Parker, C.S.; Topol, J. A Drosophila RNA Polymerase II Transcription Factor Binds to the Regulatory Site of an hsp 70 Gene. Cell 1984, 37, 273–283. [Google Scholar] [CrossRef]
- Wiederrecht, G.; Shuey, D.J.; Kibbe, W.A.; Parker, C.S. The Saccharomyces and Drosophila Heat Shock Transcription Factors Are Identical in Size and DNA Binding Properties. Cell 1987, 48, 507–515. [Google Scholar] [CrossRef]
- Gomez-Pastor, R.; Burchfiel, E.T.; Thiele, D.J. Regulation of Heat Shock Transcription Factors and Their Roles in Physiology and Disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 4–19. [Google Scholar] [CrossRef]
- Nover, L.; Bharti, K.; Döring, P.; Mishra, S.K.; Ganguli, A.; Scharf, K.D. Arabidopsis and the Heat Stress Transcription Factor World: How Many Heat Stress Transcription Factors Do We Need? Cell Stress Chaperones 2001, 6, 177–189. [Google Scholar] [CrossRef]
- Jedlicka, P.; Mortin, M.A.; Wu, C. Multiple Functions of Drosophila Heat Shock Transcription Factor In Vivo. EMBO J. 1997, 16, 2452–2462. [Google Scholar] [CrossRef] [Green Version]
- Shopland, L.S.; Hirayoshi, K.; Fernandes, M.; Lis, J.T. HSF Access to Heat Shock Elements In Vivo Depends Critically on Promoter Architecture Defined by GAGA Factor, TFIID, and RNA Polymerase II Binding Sites. Genes Dev. 1995, 9, 2756–2769. [Google Scholar] [CrossRef] [Green Version]
- Moye-Rowley, W.S. Regulation of the Transcriptional Response to Oxidative Stress in Fungi: Similarities and Differences. Eukaryot. Cell 2003, 2, 381–389. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.J.; Christians, E.S.; Liu, L.; Xiao, X.; Sohal, R.S.; Benjamin, I.J. Mouse Heat Shock Transcription Factor 1 Deficiency Alters Cardiac Redox Homeostasis and Increases Mitochondrial Oxidative Damage. EMBO. J. 2002, 21, 5164–5172. [Google Scholar] [CrossRef] [Green Version]
- Von Koskull-Döring, P.; Scharf, K.D.; Nover, L. The Diversity of Plant Heat Stress Transcription Factors. Trends Plant Sci. 2007, 12, 452–457. [Google Scholar] [CrossRef]
- Guo, M.; Liu, J.H.; Ma, X.; Luo, D.X.; Gong, Z.H.; Lu, M.H. The Plant Heat Stress Transcription Factors (HSFs): Structure, Regulation, and Function in Response to Abiotic Stresses. Front. Plant Sci. 2016, 7, 114. [Google Scholar] [CrossRef] [Green Version]
- Akerfelt, M.; Trouillet, D.; Mezger, V.; Sistonen, L. Heat Shock Factors at a Crossroad between Stress and Development. Ann. N. Y. Acad. Sci. 2007, 1113, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Wunderlich, M.; Gross-Hardt, R.; Schöffl, F. Heat Shock Factor HSFB2a Involved in Gametophyte Development of Arabidopsis thaliana and Its Expression is Controlled by a Heat-Inducible Long Non-Coding Antisense RNA. Plant Mol. Biol. 2014, 85, 541–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimoto, M.; Oshima, K.; Shinkawa, T.; Wang, B.B.; Inouye, S.; Hayashida, N.; Takii, R.; Nakai, A. Analysis of HSF4 Binding Regions Reveals Its Necessity for Gene Regulation during Development and Heat Shock Response in Mouse Lenses. J. Biol. Chem. 2008, 283, 29961–29970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veri, A.O.; Miao, Z.; Shapiro, R.S.; Tebbji, F.; O'Meara, T.R.; Kim, S.H.; Colazo, J.; Tan, K.; Vyas, V.K.; Whiteway, M.; et al. Tuning Hsf1 Levels Drives Distinct Fungal Morphogenetic Programs with Depletion Impairing Hsp90 Function and Overexpression Expanding the Target Space. PLoS Genet. 2018, 14, e1007270. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Wang, Q.; An, B.; Hou, X.; Guo, Y.; Luo, H.; He, C. Dicer-Like Proteins Regulate the Growth, Conidiation, and Pathogenicity of Colletotrichum gloeosporioides from Hevea brasiliensis. Front. Microbiol. 2018, 8, 2621. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Wang, Q.; Luo, H.; He, C.; An, B. HbWRKY40 Plays an Important Role in the Regulation of Pathogen Resistance in Hevea brasiliensis. Plant Cell Rep. 2020, 39, 1095–1107. [Google Scholar] [CrossRef]
- Ream, J.A.; Lewis, L.K.; Lewis, K.A. Rapid Agarose Gel Electrophoretic Mobility Shift Assay for Quantitating Protein: RNA Interactions. Anal. Biochem. 2016, 511, 36–41. [Google Scholar] [CrossRef] [Green Version]
- Nicholls, S.; Leach, M.D.; Priest, C.L.; Brown, A.J. Role of the Heat Shock Transcription Factor, Hsf1, in a Major Fungal Pathogen That Is Obligately Associated with Warm-Blooded Animals. Mol. Microbiol. 2009, 74, 844–861. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Sillaots, S.; Lemieux, S.; Davison, J.; Kauffman, S.; Breton, A.; Linteau, A.; Xin, C.; Bowman, J.; Becker, J.; et al. Essential Gene Identification and Drug Target Prioritization in Aspergillus fumigatus. PLoS Pathog. 2007, 3, e24. [Google Scholar] [CrossRef]
- Pinchai, N.; Perfect, B.Z.; Juvvadi, P.R.; Fortwendel, J.R.; Cramer, R.A.; Asfaw, Y.G.; Heitman, J.; Perfect, J.R.; Steinbach, W.J. Aspergillus fumigatus Calcipressin CbpA Is Involved in Hyphal Growth and Calcium Homeostasis. Eukaryot. Cell 2009, 8, 511–519. [Google Scholar] [CrossRef] [Green Version]
- Lamoth, F.; Juvvadi, P.R.; Fortwendel, J.R.; Steinbach, W.J. Heat Shock Protein 90 Is Required for Conidiation and Cell Wall Integrity in Aspergillus fumigatus. Eukaryot. Cell 2012, 11, 1324–1332. [Google Scholar] [CrossRef] [Green Version]
- Apostol, I.; Heinstein, P.F.; Low, P.S. Rapid Stimulation of an Oxidative Burst during Elicitation of Cultured Plant Cells: Role in Defense and Signal Transduction. Plant Physiol. 1989, 90, 109–116. [Google Scholar] [CrossRef] [Green Version]
- Apel, K.; Hirt, H. Reactive Oxygen Species: Metabolism, Oxidative Stress, and Signal Transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Leach, M.D.; Farrer, R.A.; Tan, K.; Miao, Z.; Walker, L.A.; Cuomo, C.A.; Wheeler, R.T.; Brown, A.J.; Wong, K.H.; Cowen, L.E. Hsf1 and Hsp90 Orchestrate Temperature-Dependent Global Transcriptional Remodelling and Chromatin Architecture in Candida albicans. Nat. Commun. 2016, 7, 11704. [Google Scholar] [CrossRef] [Green Version]
- Sheng, Y.; Wang, Y.; Meijer, H.J.; Yang, X.; Hua, C.; Ye, W.; Tao, K.; Liu, X.; Govers, F.; Wang, Y. The Heat Shock Transcription Factor PsHSF1 of Phytophthora sojae Is Required for Oxidative Stress Tolerance and Detoxifying the Plant Oxidative Burst. Environ. Microbiol. 2015, 17, 1351–1364. [Google Scholar] [CrossRef]
- Saunders, D.G.; Aves, S.J.; Talbot, N.J. Cell Cycle-Mediated Regulation of Plant Infection by the Rice Blast Fungus. Plant Cell 2010, 22, 497–507. [Google Scholar] [CrossRef] [Green Version]
- Veneault-Fourrey, C.; Barooah, M.; Egan, M.; Wakley, G.; Talbot, N.J. Autophagic Fungal Cell Death Is Necessary for Infection by the Rice Blast Fungus. Science 2006, 312, 580–583. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.R.; Hamer, J.E. MAP Kinase and cAMP Signaling Regulate Infection Structure Formation and Pathogenic Growth in the Rice Blast Fungus Magnaporthe grisea. Genes Dev. 1996, 10, 2696–2706. [Google Scholar] [CrossRef] [Green Version]
- Adachi, K.; Hamer, J.E. Divergent cAMP Signaling Pathways Regulate Growth and Pathogenesis in the Rice Blast Fungus Magnaporthe grisea. Plant Cell 1998, 10, 1361–1374. [Google Scholar] [CrossRef] [Green Version]
- Ryder, L.S.; Talbot, N.J. Regulation of Appressorium Development in Pathogenic Fungi. Curr. Opin. Plant Biol. 2015, 26, 8–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenman, H.C.; Casadevall, A. Synthesis and Assembly of Fungal Melanin. Appl. Microbiol. Biotechnol. 2012, 93, 931–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, A.K.; Gajjar, D.U.; Vasavada, A.R. DOPA and DHN Pathway Orchestrate Melanin Synthesis in Aspergillus Species. Med. Mycol. 2014, 52, 10–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nosanchuk, J.D.; Casadevall, A. The Contribution of Melanin to Microbial Pathogenesis. Cell Microbiol. 2003, 5, 203–223. [Google Scholar] [CrossRef]
- Toledo, A.V.; Franco, M.E.E.; Yanil Lopez, S.M.; Troncozo, M.I.; Saparrat, M.C.N.; Balatti, P.A. Melanins in Fungi: Types, Localization and Putative Biological Roles. Physiol. Mol. Plant Pathol. 2017, 99, 2–6. [Google Scholar] [CrossRef]
- Money, N.P.; Howard, R.J. Confirmation of a Link between Fungal Pigmentation, Turgor Pressure, and Pathogenicity Using a New Method of Turgor Measurement. Fungal Genet. Biol. 1996, 20, 217–227. [Google Scholar] [CrossRef]
- Ludwig, N.; Löhrer, M.; Hempel, M.; Mathea, S.; Schliebner, I.; Menzel, M.; Kiesow, A.; Schaffrath, U.; Deising, H.B.; Horbach, R. Melanin Is Not Required for Turgor Generation but Enhances Cell-Wall Rigidity in Appressoria of the Corn Pathogen Colletotrichum graminicola. Mol. Plant Microbe Interact. 2014, 27, 315–327. [Google Scholar] [CrossRef] [Green Version]
- Slot, J.C.; Rokas, A. Horizontal Transfer of a Large and Highly Toxic Secondary Metabolic Gene Cluster between Fungi. Curr. Biol. 2011, 21, 134–139. [Google Scholar] [CrossRef] [Green Version]
- Tsuji, G.; Kenmochi, Y.; Takano, Y.; Sweigard, J.; Farrall, L.; Furusawa, I.; Horino, O.; Kubo, Y. Novel Fungal Transcriptional Activators, Cmr1p of Colletotrichum lagenarium and pig1p of Magnaporthe grisea, Contain Cys2His2 Zinc Finger and Zn(II)2Cys6 Binuclear Cluster DNA-Binding Motifs and Regulate Transcription of Melanin Biosynthesis Genes in a Developmentally Specific Manner. Mol. Microbiol. 2000, 38, 940–954. [Google Scholar] [CrossRef]
- Cho, Y.; Srivastava, A.; Ohm, R.A.; Lawrence, C.B.; Wang, K.H.; Grigoriev, I.V.; Marahatta, S.P. Transcription Factor Amr1 Induces Melanin Biosynthesis and Suppresses Virulence in Alternaria brassicicola. PLoS Pathog. 2012, 8, e1002974. [Google Scholar] [CrossRef] [Green Version]
- Wu, C. Heat Shock Transcription Factors: Structure and Regulation. Annu. Rev. Cell Dev. Biol. 1995, 11, 441–469. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, X.; Wang, Q.; Feng, Q.; Zhang, B.; He, C.; Luo, H.; An, B. Heat Shock Transcription Factor CgHSF1 Is Required for Melanin Biosynthesis, Appressorium Formation, and Pathogenicity in Colletotrichum gloeosporioides. J. Fungi 2022, 8, 175. https://doi.org/10.3390/jof8020175
Gao X, Wang Q, Feng Q, Zhang B, He C, Luo H, An B. Heat Shock Transcription Factor CgHSF1 Is Required for Melanin Biosynthesis, Appressorium Formation, and Pathogenicity in Colletotrichum gloeosporioides. Journal of Fungi. 2022; 8(2):175. https://doi.org/10.3390/jof8020175
Chicago/Turabian StyleGao, Xuesheng, Qiannan Wang, Qingdeng Feng, Bei Zhang, Chaozu He, Hongli Luo, and Bang An. 2022. "Heat Shock Transcription Factor CgHSF1 Is Required for Melanin Biosynthesis, Appressorium Formation, and Pathogenicity in Colletotrichum gloeosporioides" Journal of Fungi 8, no. 2: 175. https://doi.org/10.3390/jof8020175