
Antifungal Mechanism of Metabolites from Newly Isolated Streptomyces sp. Y1-14 against Banana Fusarium Wilt Disease Using Metabolomics

Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Actinomycetes
2.2. Screening of Antifungal Actinomycetes
2.3. Identification of Antifungal Actinomycetes
2.4. Assay of Abroad-Spectrum Antifungal Activity of Strain Y1-14
2.5. Preparation of Ethanol Extracts
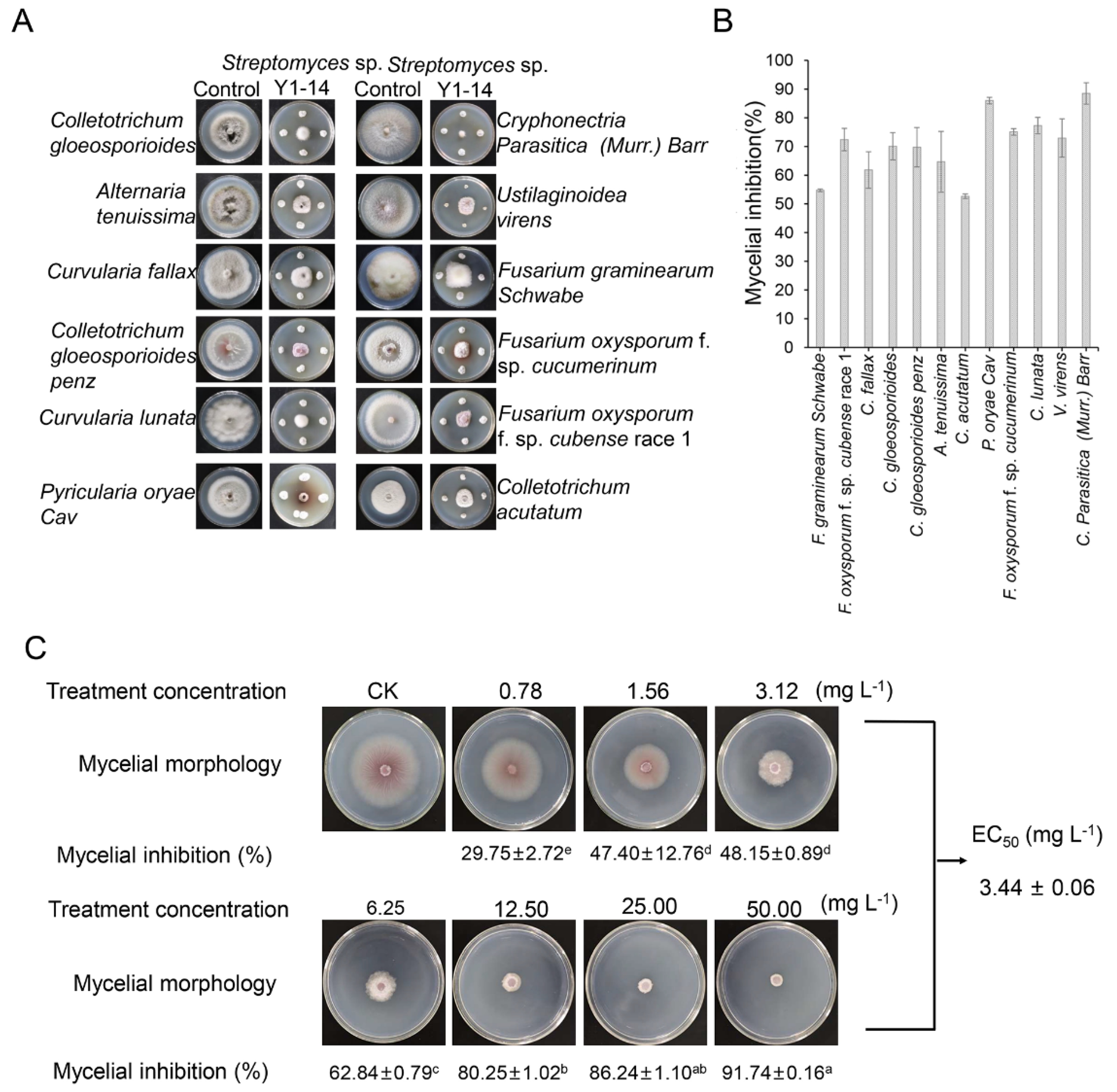
2.6. Inhibition Percentage of Strain Y1-14 Extracts on Mycelial Growth
2.7. Effect of Strain Y1-14 Extracts on Morphological Characteristics of Foc TR4 Mycelia
2.8. Effect of Strain Y1-14 Extracts on Spore Germination of Foc TR4
2.9. Effects of Strain Y1-14 Extracts on Ultrastructure of Foc TR4
2.10. Determination of Antifungal Activity of Volatile Organic Compounds from Strain Y1-14
2.11. Component Analysis of VOCs by Head-Space Gas Chromatography–Mass Spectrometry
2.12. Effects of Strain Y1-14 Extracts on Metabolism of Foc TR4
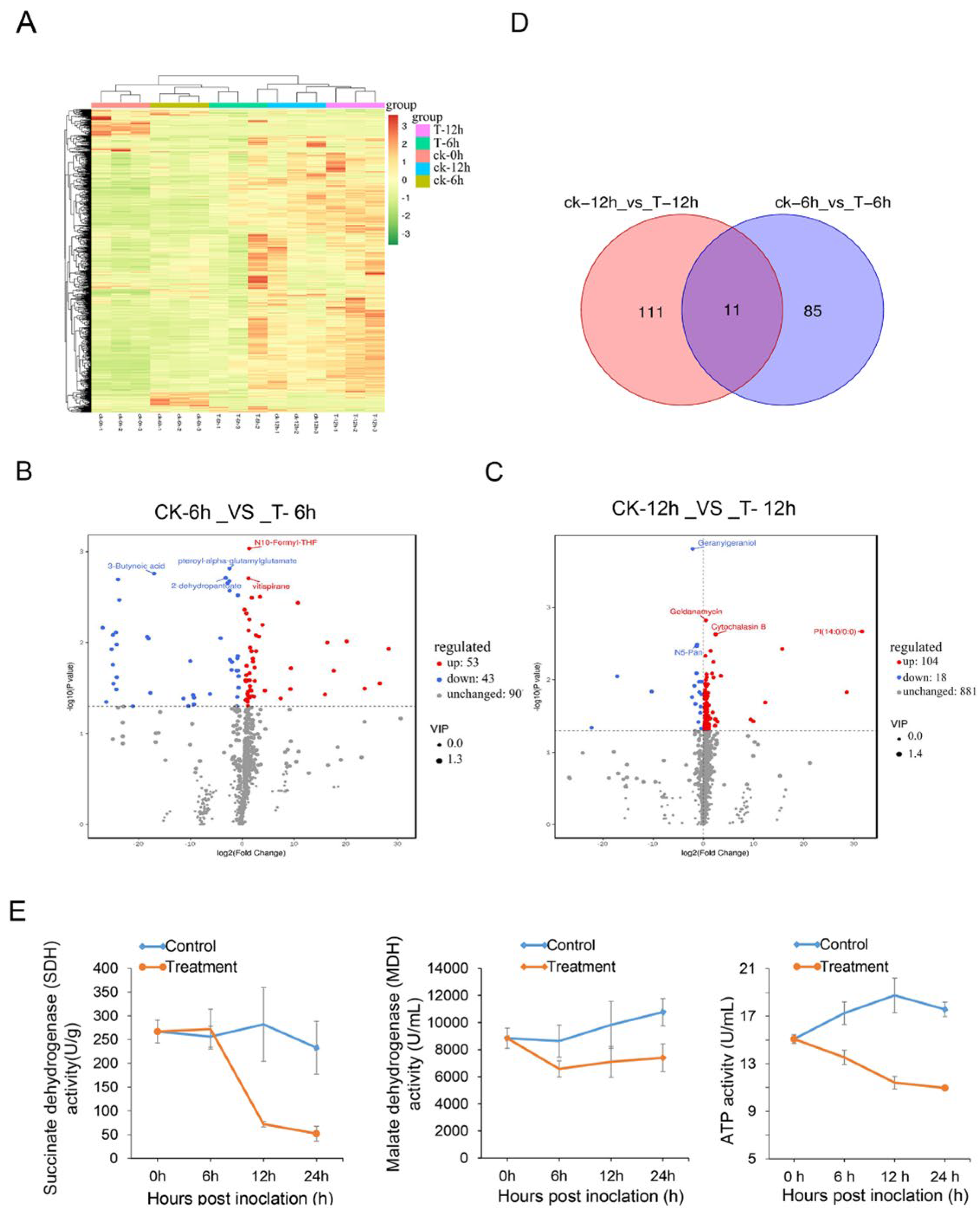
2.13. Determination of Mitochondrial Enzyme Activities
2.14. Data Analysis
3. Results
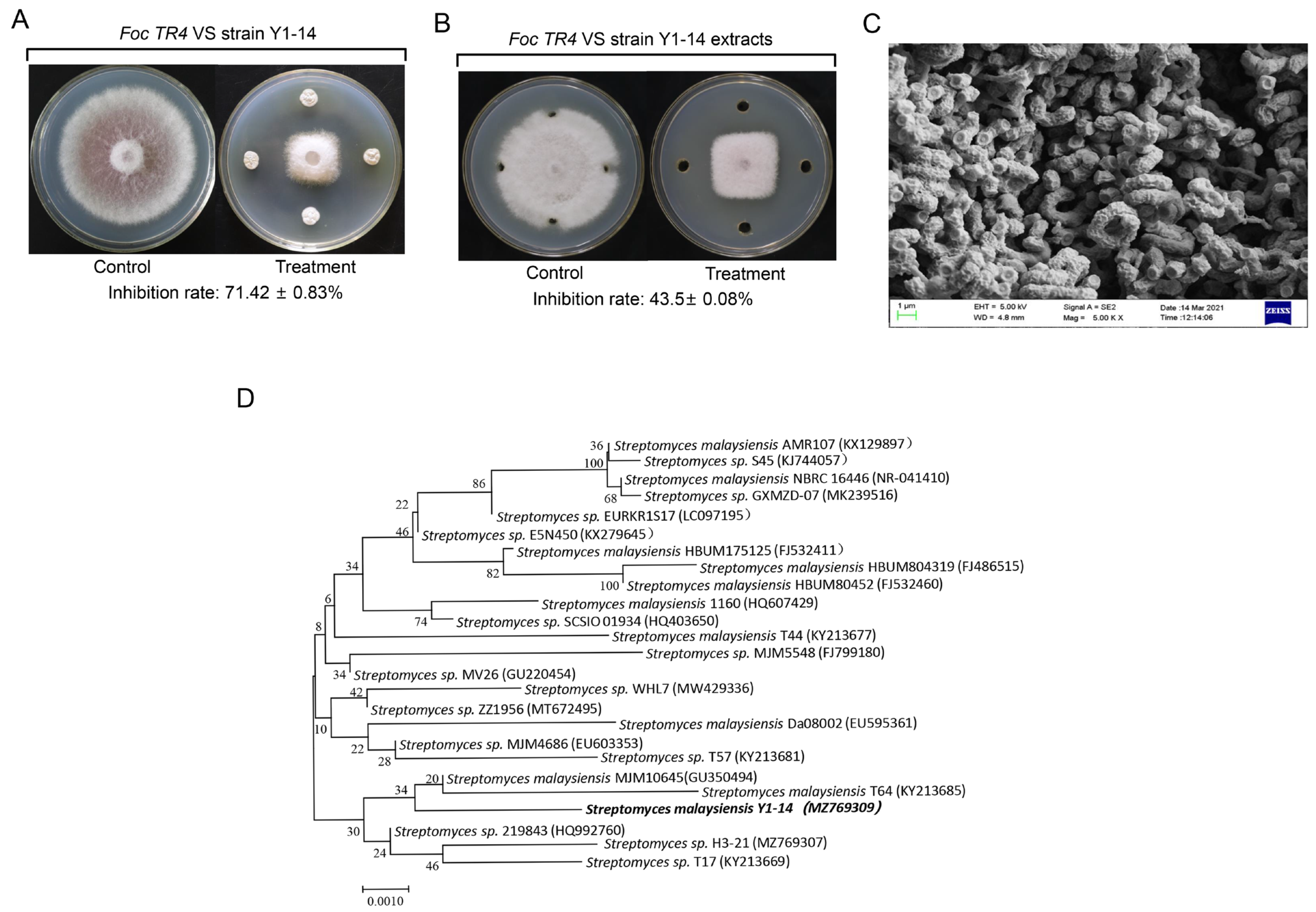
3.1. Screening and Identification of Actinomycetes with Antifungal Activity
3.2. Assay of Broad-Spectrum Antifungal Activity
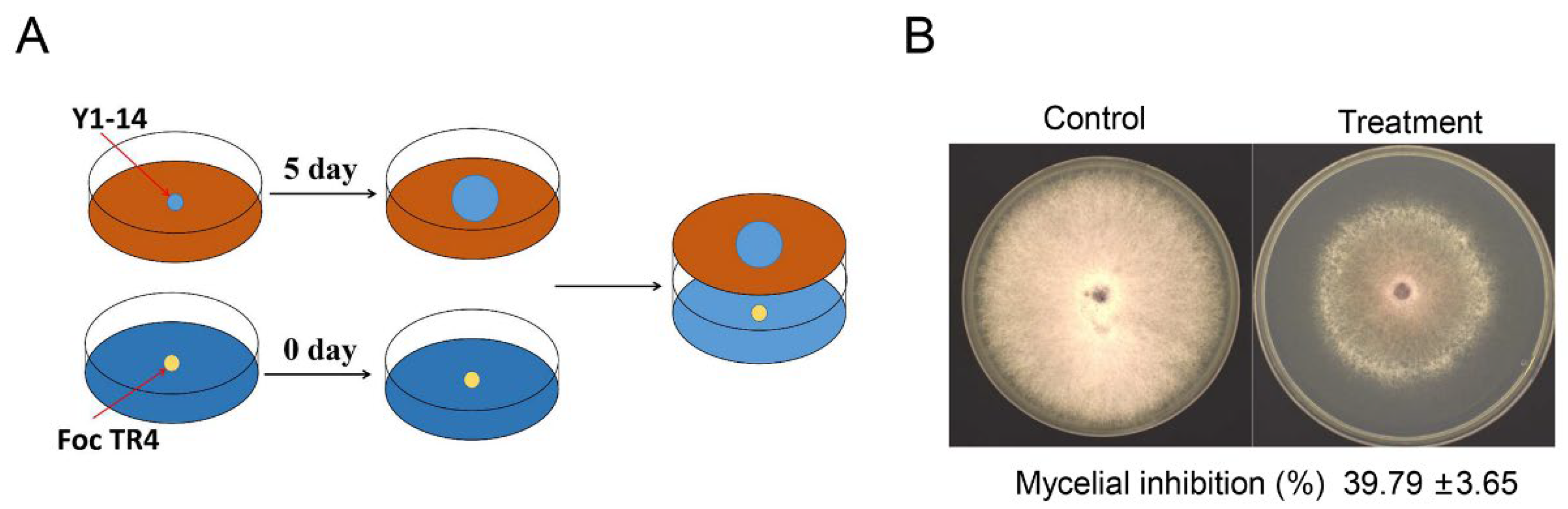
3.3. Inhibition Activity of Streptomyces sp. Y1-14 against Foc TR4
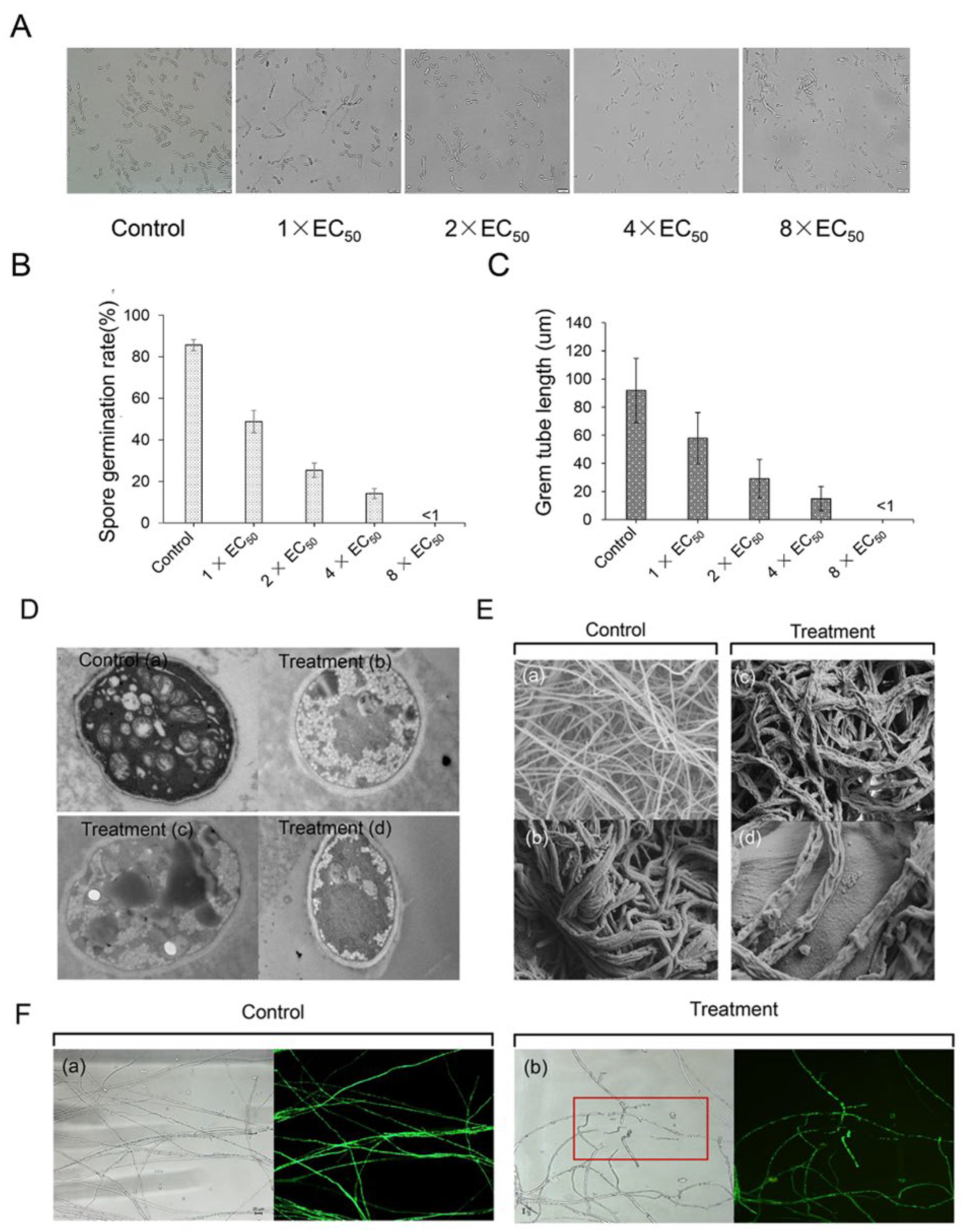
3.4. Effect of Streptomyces sp. Y1-14 Extracts on Spore Germination and Germ Tube Elongation of Foc TR4 In Vitro
3.5. Effect of Streptomyces sp. Y1-14 Extracts on Mycelial Morphology and Ultrastructure of Foc TR4
3.6. Chemical Constituent Analysis of VOCs
3.7. Effects of Streptomyces sp. Y1-14 Extracts on Foc TR4 Metabolites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, H.; Hu, C.; Xie, A.; Wu, S.; Dou, T. Overexpression of MpbHLH transcription factor, an encoding ICE1-like protein, enhances Foc TR4-resistance of Cavendish banana. Sci. Hortic. 2022, 291, 110590. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations: FAOSTAT Crops. 2019. Available online: https://www.fao.org/faostat/en/#data/QC (accessed on 13 November 2019).
- Rebouas, T.A.; de Jesus Rocha, A.; Cerqueira, T.S.; Adorno, P.R.; Barreto, R.Q.; Ferreira, M.; Lino, L.S.M.; Amorim, V.B.D.; Santos-Serejo, J.A.D.; Haddad, F.; et al. Pre-selection of banana somaclones resistant to Fusarium oxysporum f. sp. cubense, subtropical race 4. Crop Prot. 2021, 147, 105692. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Jv, H.; Lu, M.; Wang, B.; Ruan, Y. Significant decline in banana Fusarium wilt disease is associated with soil microbiome reconstruction under chilli pepper-banana rotation. Eur. J. Soil Biol. 2020, 97, 103154. [Google Scholar] [CrossRef]
- Tripathi, L.; Ntui, V.O.; Tripathi, J.N. CRISPR/Cas9-based genome editing of banana for disease resistance. Curr. Opin. Plant Biol. 2020, 56, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Zapata-Sarmientoa, D.H.; Palacios-Palaa, E.F.; Rodríguez-Hernándezb, A.A.; Melchora, D.L.M.; Rodríguez-Monroya, M.; Sepúlveda-Jiménez, G. Trichoderma asperellum, a potential biological control agent of Stemphylium vesicarium, on onion (Allium cepa L.). Biol. Control. 2019, 140, 104105. [Google Scholar] [CrossRef]
- Bubici, G.; Kaushal, M.; Prigigallo, M.I.; Gómez-Lama Cabanás, C.; Mercado-Blanco, J. Biological control agents against Fusarium wilt of banana. Front. Microbiol. 2019, 10, 616. [Google Scholar] [CrossRef] [Green Version]
- Thangavelu, R.; Gopi, M. Combined application of native Trichoderma isolates possessing multiple functions for the control of Fusarium wilt disease in banana cv. grand naine. Biocontrol Sci. Technol. 2015, 25, 1147–1164. [Google Scholar] [CrossRef]
- Li, C.; Cheng, P.; Zheng, L.; Li, Y.; Yu, G. Comparative genomics analysis of two banana Fusarium wilt biocontrol endophytes Bacillus subtilis R31 and TR21 provides insights into their differences on phytobeneficial trait. Genomics 2021, 113, 900–909. [Google Scholar] [CrossRef]
- Xu, Z.; Wang, M.; Du, J.; Huang, T.; Chen, Y. Isolation of Burkholderia sp. HQB-1, a promising biocontrol bacteria to protect banana against Fusarium wilt through phenazine-1-carboxylic acid secretion. Front. Microbiol. 2020, 11, 605152. [Google Scholar] [CrossRef]
- Bérdy, J. Bioactive microbial metabolites. J. Antibiot. 2005, 58, 1–26. [Google Scholar] [CrossRef]
- Bhatti, A.A.; Haq, S.; Bhat, R.A. Actinomycetes benefaction role in soil and plant health. Microb. Pathog. 2017, 111, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Bu, Q.T.; Li, Y.P.; Xie, H.; Li, J.F.; Li, Y.Q. Rational engineering strategies for achieving high-yield, high-quality and high-stability of natural product production in actinomycetes. Metab. Eng. 2021, 67, 198–215. [Google Scholar] [CrossRef] [PubMed]
- Dede, A.; Güven, K.; Ahn, N. Isolation, plant growth-promoting traits, antagonistic effects on clinical and plant pathogenic organisms and identification of actinomycetes from olive rhizosphere. Microb. Pathog. 2020, 143, 104134. [Google Scholar] [CrossRef]
- Shan, W.; Zhou, Y.; Liu, H.; Yu, X. Endophytic actinomycetes from tea plants (Camellia sinensis): Isolation, abundance, antimicrobial, and plant-growth-promoting activities. Biomed Res. Int. 2018, 1470305. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Li, K.; Zhou, D.; Zhang, M.; Qi, D.; Jing, T.; Zang, X.; Qia, C.; Wang, W.; Xie, J. Biological control of banana wilt disease caused by Fusarium oxyspoum f. sp. cubense using Streptomyces sp. H4. Biol. Control 2021, 155, 104524. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, H.; Huang, Y.; Peng, J.; Xie, J.; Wang, W. Isolation and evaluation of rhizosphere actinomycetes with potential application for biocontrolling Fusarium wilt of banana caused by Fusarium oxysporum f. sp. cubense tropical race 4. Front. Microbiol. 2021, 12, 763038. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cai, B.; Li, K.; Zhao, Y.; Li, C.; Liu, S.; Xiang, D.; Zhang, L.; Xie, J.; Wang, W. Biological control of Fusarium oxysporum f. sp. cubense tropical race 4 in banana plantlets using newly isolated Streptomyces sp. WHL7 from marine soft coral. Plant Dis. 2022, 106, 254–259. [Google Scholar] [CrossRef]
- Ou, Y.; Penton, C.R.; Geisen, S.; Shen, Z.; Shen, Q. Deciphering underlying drivers of disease suppressiveness against pathogenic Fusarium oxysporum. Front. Microbiol. 2019, 10, 2535. [Google Scholar] [CrossRef] [Green Version]
- Williams, S.T.; Goodfellow, M.; Alderson, G.; Wellington, E.; Sackin, M.J. Numerical classification of Streptomyces and related genera. J. Gen. Microbiol. 1983, 129, 1743–1813. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.; Zhao, Y.; Zhou, D.; Qi, D.; Wang, W. A newly isolated Streptomyces sp. YYS-7 with a broad-spectrum antifungal activity improves the banana plant resistance to Fusarium oxysporum f. sp. cubense tropical race 4. Front. Microbiol. 2020, 11, 1712. [Google Scholar] [CrossRef]
- Zou, N.; Zhou, D.; Chen, Y.; Lin, P.; Chen, Y.; Wang, W.; Xie, J.; Wang, M. A novel antifungal actinomycete Streptomyces sp. strain h3-2 effectively controls banana Fusarium wilt. Front. Microbiol. 2021, 12, 706647. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.S.; EI-Gendy, A.O.; Ahmed, R.R.; Hassan, H.M.; EI-Kabbany, H.M.; Merdash, A.G. Exploring the antimicrobial and antitumor potentials of Streptomyces sp. AGM12-1 isolated from Egyptian soil. Front. Microbiol. 2017, 8, 438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, D.; Zou, L.; Zhou, D.; Chen, Y.; Wang, W. Taxonomy and broad-spectrum antifungal activity of Streptomyces sp. SCA3-4 isolated from rhizosphere soil of Opuntia stricta. Front. Microbiol. 2019, 10, 1390. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. Mega7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Jing, T.; Zhou, D.; Zhang, M.; Yun, T.; Qi, D.; Wei, Y.; Chen, Y.; Zang, X.; Wang, W.; Xie, J. Newly isolated Streptomyces sp. JBS5-6 as a potential biocontrol agent to control banana Fusarium wilt: Genome sequencing and secondary metabolite cluster profiles. Front. Microbiol. 2020, 11, 602591. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jing, T.; Zhou, D.; Zhang, M.; Qi, D.; Zang, X.; Zhao, Y.; Li, K.; Tang, W.; Chen, Y.; et al. Biocontrol efficacy and possible mechanism of Streptomyces sp. H4 against postharvest anthracnose caused by Colletotrichum fragariae on strawberry fruit. Postharvest Biol. Technol. 2021, 175, 111401. [Google Scholar] [CrossRef]
- Vanewijk, P.; Hoekstra, J. Calculation of the EC50 and its confidence interval when subtoxic stimulus is present. Ecotoxicol. Environ. Saf. 1993, 25, 25–32. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, D.; Qi, D.; Gao, Z.; Luo, Y. Growth promotion and disease suppression ability of a Streptomyces sp. cb-75 from banana rhizosphere soil. Front. Microbiol. 2017, 8, 2704. [Google Scholar] [CrossRef]
- Dennis, C.; Webster, J. Antagonistic properties of species-groups of Trichoderma: I. Production of non-volatile antibiotics. Trans. Brit. Mycol. Soc. 1971, 57, 25–39. [Google Scholar] [CrossRef]
- Rajani, P.; Rajasekaran, C.; Vasanthakumari, M.M.; Olsson, S.B.; Shaanker, R.U. Inhibition of plant pathogenic fungi by endophytic Trichoderma spp. through mycoparasitism and volatile organic compounds. Microbiol. Res. 2021, 242, 126595. [Google Scholar] [CrossRef]
- Francesco, A.D.; Ugolini, L.; Lazzeri, L.; Mari, M. Production of volatile organic compounds by Aureobasidium pullulans as a potential mechanism of action against postharvest fruit pathogens. Biol. Control. 2015, 81, 8–14. [Google Scholar] [CrossRef]
- Mizutani, A.; Yukioka, H.; Tamura, H.; Miki, N.; Masuko, M.; Takeda, R. Respiratory characteristics in Pyricularia oryzae exposed to a novel alkoxyiminoacetamide fungicide. Phytopathology 1995, 85, 306–311. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, C.; Li, F.; Li, Z.; Chen, M.; Wang, Y.; Qiao, X.; Zhang, H. Fumigant activity of volatiles from Streptomyces alboflavus TD-1 against Fusarium moniliforme Sheldon. J. Microbiol. 2013, 51, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Gerber, N.N. A volatile metabolite of actinomycetes, 2-methylisoborneol. J. Antibiot. 1969, 22, 508–509. [Google Scholar] [CrossRef] [Green Version]
- Ogura, T.; Sunairi, M.; Nakajima, M. 2-methylisoborneol and geosmin, the main sources of soil odor, inhibit the germination of brassicaceae seeds. Soil Sci. Plant Nutr. 2000, 46, 217–227. [Google Scholar] [CrossRef]
- Rădulescu, M.; Jianu, C.; Lukinich-Gruia, A.T.; Mioc, M.; Mioc, A.; Șoica, C.; Stana, L.G. Chemical composition, in vitro and in silico antioxidant potential of Melissa officinalis subsp. officinalis essential oil. Antioxidants 2021, 10, 1081. [Google Scholar] [CrossRef]
- Dalli, M.; Azizi, S.E.; Benouda, H.; Azghar, A.; Tahri, M.; Bouammali, B.; Maleb, A.; Gseyra, N. Molecular composition and antibacterial effect of five essential oils extracted from Nigella sativa L. seeds against multidrug-resistant bacteria: A comparative study. Evid. Based Complement. Altern. Med. 2021, 2021, 6643765. [Google Scholar] [CrossRef]
- Schmidt, R.; Cordovez, V.; de Boer, W.; Raaijmakers, J.; Garbeva, P. Volatile affairs in microbial interactions. ISME J. 2015, 9, 2329–2335. [Google Scholar] [CrossRef] [Green Version]
- Audrain, B.; Farag, M.A.; Ryu, C.M.; Ghigo, J.M. Role of bacterial volatile compounds in bacterial biology. FEMS Microbiol. Rev. 2015, 39, 222–233. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Huang, C.; Xue, Y.; Li, S.; Lu, L.; Wang, C. Biofumigation with volatile organic compounds from Streptomyces alboflavus TD-1 and pure chemicals to control Aspergillus ochraceus. Ann. Appl. Biol. 2018, 173, 313–322. [Google Scholar] [CrossRef]
- Gouda, S.; Das, G.; Sen, S.K.; Shin, H.S.; Patra, J.K. A treasure house of bioactive compounds of medicinal importance. Front. Microbiol. 2016, 7, 1538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, P.C.; Liu, S.C.; Ho, M.C.; Huang, T.W.; Huang, C.H. A soil-isolated Streptomyces spororaveus species produces a high-molecular-weight antibiotic AF1 against fungi and Gram-positive bacteria. Antibiotics 2022, 11, 679. [Google Scholar] [CrossRef] [PubMed]
- Kopp, D.; Sunna, A. Alternative carbohydrate pathways-enzymes, functions and engineering. Crit. Rev. Biotechnol. 2020, 40, 895–912. [Google Scholar] [CrossRef]
- Noor, E.; Eden, E.; Milo, R.; Alon, U. Central carbon metabolism as a minimal biochemical walk between precursors for biomass and energy. Mol Cell. 2010, 39, 809–820. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.L.; Cox, M.M. Lehninger Principles of Biochemistry, 4th ed.; W.H. Freeman and Company: New York, NY, USA, 2005. [Google Scholar]
- Flamholz, A.; Noor, E.; Bar-Even, A.; Liebermeister, W.; Milo, R. Glycolytic strategy as a tradeoff between energy yield and protein cost. Proc. Natl. Acad. Sci. USA 2013, 110, 10039–10044. [Google Scholar] [CrossRef] [Green Version]
- Bertels, L.K.; Fernández, M.L.; Heinisch, J.J. The pentose phosphate pathway in yeasts-more than a poor cousin of glycolysis. Biomolecules 2021, 11, 725. [Google Scholar] [CrossRef]
- Wood, T. The Pentose Phosphate Pathway; Academic Press: Orlando, FL, USA, 1985. [Google Scholar]
- Sprenger, G.A. Genetics of pentose-phosphate pathway enzymes of Escherichia coli K-12. Arch. Microbiol. 1995, 164, 324–330. [Google Scholar] [CrossRef]
- Yin, X.; Li, J.; Shin, H.D.; Du, G.; Liu, L.; Chen, J. Metabolic engineering in the biotechnological production of organic acids in the tricarboxylic acid cycle of microorganisms: Advances and prospects. Biotechnol. Adv. 2015, 33, 830–841. [Google Scholar] [CrossRef]
- Fernie, A.R.; Carrari, F.; Sweetlove, L.J. Respiratory metabolism: Glycolysis, the TCA cycle and mitochondrial electron transport. Curr. Opin. Plant Biol. 2004, 7, 254–261. [Google Scholar] [CrossRef]
- Eniafe, J.; Jiang, S. The functional roles of TCA cycle metabolites in cancer. Oncogene 2021, 40, 3351–3363. [Google Scholar] [CrossRef]
- Jia, Y.; Wong, D.C.; Sweetman, C.; Bruning, J.B.; Ford, C.M. New insights into the evolutionary history of plant sorbitol dehydrogenase. BMC Plant Biol. 2015, 15, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, X.; Liu, W.; Su, Y.; Liu, X.; Liu, J.; Liu, N.; Wang, G.; Jiao, X.; Fan, X.; Xue, C.; et al. Studies on the novel pyridine sulfide containing SDH based heterocyclic amide fungicide. Pest Manag. Sci. 2020, 76, 2368–2378. [Google Scholar] [CrossRef] [PubMed]
- Venkat, S.; Gregory, C.; Sturges, J.; Gan, Q.; Fan, C. Studying the lysine acetylation of malate dehydrogenase. J. Mol. Biol. 2017, 429, 1396–1405. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RT a (min) | Possible Compound b,c | Relative Peak Area (%) | Classification |
|---|---|---|---|
| 1.343 | Ala-Gly | 1.4163 | Others |
| 2.081 | Ethyl Acetate | 1.0644 | |
| 4.089 | Silanediol, dimethyl- | 1.0444 | |
| 11.026 | 3-Furancarboxylic acid, methyl ester | 1.1225 | |
| 30.66 | trans-1,10-Dimethyl-trans-9-decalinol | 21.05 | |
| 12.949 | p-Menth-8-ene, 3-methylene- | 3.9783 f | Hydrocarbons |
| 22.687 | Naphthalene, 1,2,3,4,4a,5,6,8a-octahydro-4a,8-dimethyl- | 9.5415 | |
| 22.894 | Cyclohexane, 1,1,4,4-tetramethyl-2,5-dimethylene- | 4.0992 | |
| 23.551 | Bicyclo [4.3.0] non-3-ene, 3,4,7-trimethyl- | 1.1359 | |
| 23.74 | (2Z)-2-ethylidene-1,7,7-trimethylbicyclo [2.2.1] heptane | 2.2966 | |
| 30.34 | Tetradecane | 1.1357 | |
| 20.836 | 2-Methylisoborneol | 11.9192 | Alcohols |
| 32.681 | Octahydro-2,2,5a,9-tetramethyl-2H-3,9a-methano-1-benzoxepin | 3.8177 | |
| 35.365 | Palustrol | 3.2699 | |
| 39.452 | Nerolidol | 1.4601 | |
| 28.035 | β-Chamigrene | 1.3231 | Terpene |
| 29.732 | Cyclohexane, 1-ethenyl-1-methyl-2,4-bis(1-methylethenyl)-, [1S-(1à,2á,4á)]- | 1.0102 | |
| 30.759 | (−)-α-Cedrene | 2.179 | |
| 32.357 | Guaia-10(14),11-diene | 2.7849 | |
| 33.415 | β-Cubebene | 4.2486 | |
| 34.095 | γ-Gurjunene | 1.6816 | |
| 34.685 | γ-Cadinene | 1.0292 | |
| 34.941 | Cadina-1(10),4-diene | 2.3372 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, M.; Cheng, Q.; Cai, B.; Chen, Y.; Wei, Y.; Qi, D.; Li, Y.; Yan, L.; Li, X.; Long, W.; et al. Antifungal Mechanism of Metabolites from Newly Isolated Streptomyces sp. Y1-14 against Banana Fusarium Wilt Disease Using Metabolomics. J. Fungi 2022, 8, 1291. https://doi.org/10.3390/jof8121291
Cao M, Cheng Q, Cai B, Chen Y, Wei Y, Qi D, Li Y, Yan L, Li X, Long W, et al. Antifungal Mechanism of Metabolites from Newly Isolated Streptomyces sp. Y1-14 against Banana Fusarium Wilt Disease Using Metabolomics. Journal of Fungi. 2022; 8(12):1291. https://doi.org/10.3390/jof8121291
Chicago/Turabian StyleCao, Miaomiao, Qifeng Cheng, Bingyu Cai, Yufeng Chen, Yongzan Wei, Dengfeng Qi, Yuqi Li, Liu Yan, Xiaojuan Li, Weiqiang Long, and et al. 2022. "Antifungal Mechanism of Metabolites from Newly Isolated Streptomyces sp. Y1-14 against Banana Fusarium Wilt Disease Using Metabolomics" Journal of Fungi 8, no. 12: 1291. https://doi.org/10.3390/jof8121291