
Improved Itaconate Production with Ustilago cynodontis via Co-Metabolism of CO2-Derived Formate

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Analytical Methods
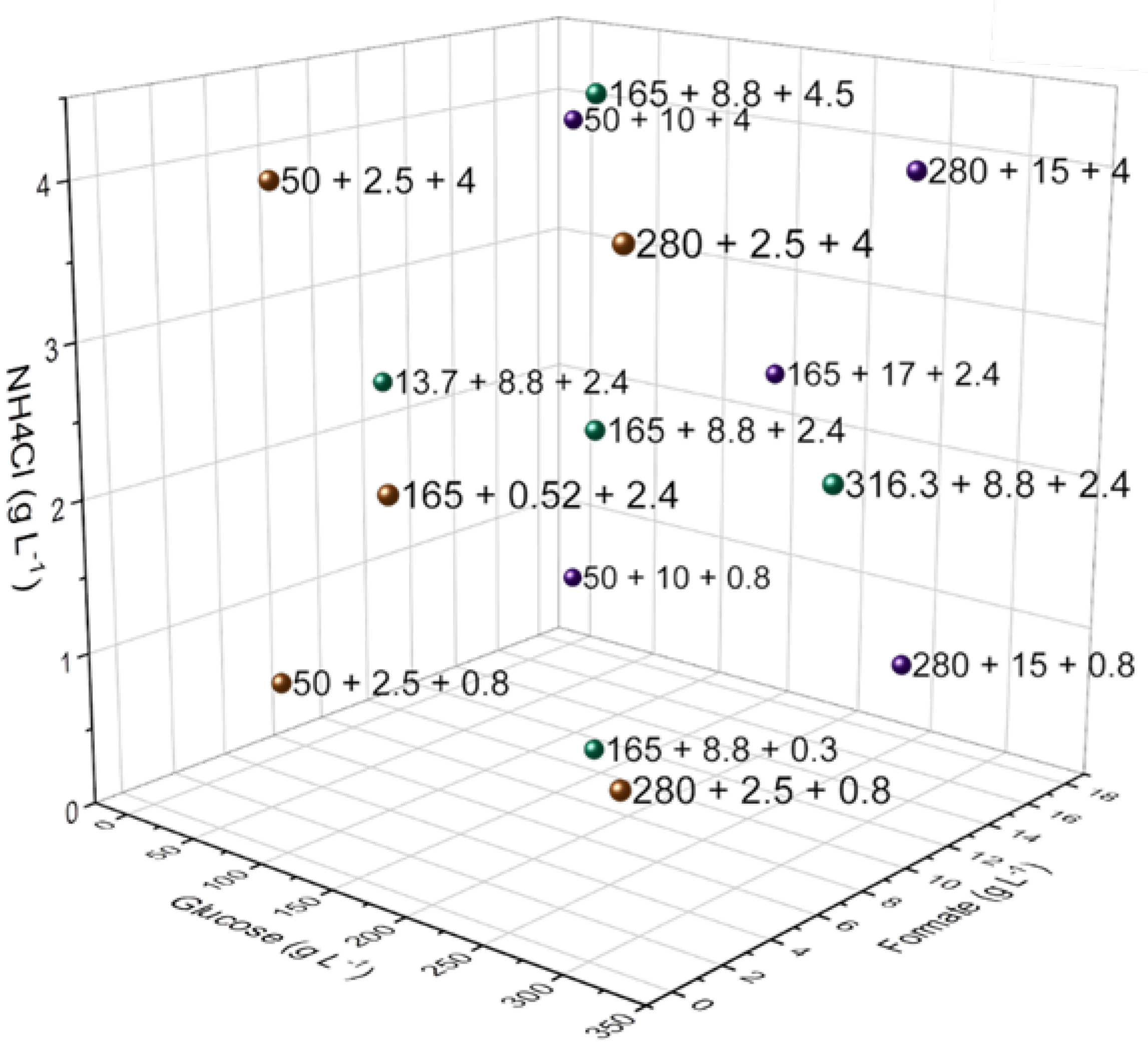
2.3. Design of Experiments (DoE)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Glucose [g L−1] | Formate [g L−1] | NH4Cl [g L−1] | CaCO3 [g L−1] |
|---|---|---|---|---|
| 1 | 13.7 | 8.8 | 2.4 | 33 |
| 2 | 50 | 2.5 | 0.8 | |
| 3 | 50 | 2.5 | 4 | |
| 4 | 50 | 15 | 0.8 | |
| 5 | 50 | 15 | 4 | |
| 6 | 165 | 0.5 | 2.4 | 66 |
| 7 | 165 | 8.8 | 0.3 | |
| 8 | 165 | 8.8 | 2.4 | |
| 9 | 165 | 8.8 | 4.5 | |
| 10 | 165 | 17 | 2.4 | |
| 11 | 280 | 2.5 | 0.8 | |
| 12 | 280 | 2.5 | 4 | |
| 13 | 280 | 15 | 0.8 | |
| 14 | 280 | 15 | 4 | |
| 15 | 316.3 | 8.8 | 2.4 |
2.4. CO2 Hydrogenation in Presence of NaOH and Cis-[RuCl2(C12-dppm)2] in Apolar Solvent/H2O
3. Results and Discussion
3.1. Formate Co-Metabolization with Metabolically Engineered U. cynodontis Strains
3.2. Optimization of Formate Co-Metabolization via Design of Experiment (DoE) Approach
3.3. Implementing CO2-Derived Formate
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ullmann, L.; Phan, A.N.; Kaplan, D.K.; Blank, L.M. Ustilaginaceae Biocatalyst for Co-Metabolism of CO2-Derived Substrates toward Carbon-Neutral Itaconate Production. J. Fungi 2021, 7, 98. [Google Scholar] [CrossRef] [PubMed]
- Gopaliya, D.; Kumar, V.; Khare, S.K. Recent advances in itaconic acid production from microbial cell factories. Biocatal. Agric. Biotechnol. 2021, 36, 102130. [Google Scholar] [CrossRef]
- Karaffa, L.; Díaz, R.; Papp, B.; Fekete, E.; Sándor, E.; Kubicek, C.P. A deficiency of manganese ions in the presence of high sugar concentrations is the critical parameter for achieving high yields of itaconic acid by Aspergillus terreus. Appl. Microbiol. Biotechnol. 2015, 99, 7937–7944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bafana, R.; Pandey, R.A. New approaches for itaconic acid production: Bottlenecks and possible remedies. Crit. Rev. Biotechnol. 2018, 38, 68–82. [Google Scholar] [CrossRef]
- Hosseinpour Tehrani, H.; Becker, J.; Bator, I.; Saur, K.; Meyer, S.; Rodrigues Lóia, A.C.; Blank, L.M.; Wierckx, N. Integrated strain- And process design enable production of 220 g L−1 itaconic acid with Ustilago maydis. Biotechnol. Biofuels 2019, 12, 263. [Google Scholar] [CrossRef]
- Hosseinpour Tehrani, H. Engineering the Morphology and Metabolism of Ustilago to Expand the Process Window for Itaconic Acid Production. 2019, Volume 13. Available online: http://publications.rwth-aachen.de/record/765515 (accessed on 29 October 2022).
- Hosseinpour Tehrani, H.; Geiser, E.; Engel, M.; Hartmann, S.K.; Hossain, A.H.; Punt, P.J.; Blank, L.M.; Wierckx, N. The interplay between transport and metabolism in fungal itaconic acid production. Fungal Genet. Biol. 2019, 125, 45–52. [Google Scholar] [CrossRef]
- Tehrani, H.H.; Tharmasothirajan, A.; Track, E.; Blank, L.M.; Wierckx, N. Engineering the morphology and metabolism of pH tolerant Ustilago cynodontis for efficient itaconic acid production. Metab. Eng. 2019, 54, 293–300. [Google Scholar] [CrossRef]
- Phan, A.N.; Blank, L.M. GC-MS-Based Metabolomics for the Smut Fungus Ustilago maydis: A Comprehensive Method Optimization to Quantify Intracellular Metabolites. Front. Mol. Biosci. 2020, 7, 211. [Google Scholar] [CrossRef]
- Becker, J.; Tehrani, H.H.; Ernst, P.; Blank, L.M.; Wierckx, N. An optimized ustilago maydis for itaconic acid production at maximal theoretical yield. J. Fungi 2021, 7, 20. [Google Scholar] [CrossRef]
- Wu, J.; Huang, Y.; Ye, W.; Li, Y. CO2 Reduction: From the Electrochemical to Photochemical Approach. Advanced Science 2017, 4, 1700194. [Google Scholar] [CrossRef]
- Liu, X.; Inagaki, S.; Gong, J. Heterogeneous Molecular Systems for Photocatalytic CO2 Reduction with Water Oxidation. Angew. Chem.—Int. Ed. 2016, 55, 14924–14950. [Google Scholar] [CrossRef]
- Francke, R.; Schille, B.; Roemelt, M. Homogeneously Catalyzed Electroreduction of Carbon Dioxide—Methods, Mechanisms, and Catalysts. Chem. Rev. 2018, 118, 4631–4701. [Google Scholar] [CrossRef]
- Artz, J.; Müller, T.E.; Thenert, K.M.; Kleinekorte, J.; Meys, R.; Sternberg, A.; Bardow, A.; Leitner, W. Sustainable Conversion of Carbon Dioxide: An Integrated Review of Catalysis and Life Cycle Assessment. Chem. Rev. 2018, 118, 434–504. [Google Scholar] [CrossRef]
- Klankermayer, J.; Wesselbaum, S.; Beydoun, K.; Leitner, W. Selective Catalytic Synthesis Using the Combination of Carbon Dioxide and Hydrogen: Catalytic Chess at the Interface of Energy and Chemistry. Angew. Chem.—Int. Ed. 2016, 55, 7296–7343. [Google Scholar] [CrossRef]
- Kinzel, N.W.; Werlé, C.; Leitner, W. Transition Metal Complexes as Catalysts for the Electroconversion of CO2: An Organometallic Perspective. Angew. Chem.—Int. Ed. 2021, 60, 11628–11686. [Google Scholar] [CrossRef]
- Cauwenbergh, R.; Das, S. Photochemical reduction of carbon dioxide to formic acid. Green Chem. 2021, 23, 2553–2574. [Google Scholar] [CrossRef]
- Leitner, W. Carbon Dioxide as a Raw Material: The Synthesis of Formic Acid and Its Derivatives from CO2. Angew. Chem. Int. Ed. Engl. 1995, 34, 2207–2221. [Google Scholar] [CrossRef]
- Navarrete, C.; Jacobsen, I.H.; Martínez, J.L.; Procentese, A. Cell factories for industrial production processes: Current issues and emerging solutions. Processes 2020, 8, 768. [Google Scholar] [CrossRef]
- Babel, W. The Auxiliary Substrate Concept: From simple considerations to heuristically valuable knowledge. Eng. Life Sci. 2009, 9, 285–290. [Google Scholar] [CrossRef]
- Babel, W.; Müller, R.H.; Markuske, K.D. Improvement of growth yield of yeast on glucose to the maximum by using an additional energy source. Arch. Microbiol. 1983, 136, 203–208. [Google Scholar] [CrossRef]
- Scott, M.; Molinos, B.B.; Westhues, C.; Franciò, G.; Leitner, W. Aqueous Biphasic Systems for the Synthesis of Formates by Catalytic CO2 Hydrogenation: Integrated Reaction and Catalyst Separation for CO2-Scrubbing Solutions. ChemSusChem 2017, 10, 1085–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leitner, W.; Franciò, G.; Scott, M.; Westhues, C.; Langanke, J.; Lansing, M.; Hussong, C.; Erdkamp, E. Carbon2Polymer—Chemical Utilization of CO2 in the Production of Isocyanates. Chem. Ing. Tech. 2018, 90, 1504–1512. [Google Scholar] [CrossRef]
- Jens, C.M.; Scott, M.; Liebergesell, B.; Westhues, C.G.; Schäfer, P.; Franciò, G.; Leonhard, K.; Leitner, W.; Bardow, A. Rh-Catalyzed Hydrogenation of CO 2 to Formic Acid in DMSO-Based Reaction Media. Adv. Synth. Catal. 2018, 361, 7903. [Google Scholar] [CrossRef] [Green Version]
- Guntermann, N.; Mengers, H.G.; Franciò, G.; Blank, L.M.; Leitner, W. Bio-energy conversion with carbon capture and utilization (BECCU): Integrated biomass fermentation and chemo-catalytic CO2 hydrogenation for bioethanol and formic acid co-production. Green Chem. 2021, 23, 9860–9864. [Google Scholar] [CrossRef]
- Scott, M.; Westhues, C.G.; Kaiser, T.; Baums, J.C.; Jupke, A.; Franciò, G.; Leitner, W. Methylformate from CO2: An integrated process combining catalytic hydrogenation and reactive distillation. Green Chem. 2019, 21, 6307–6317. [Google Scholar] [CrossRef] [Green Version]
- Park, D.S.; Abdelrahman, O.A.; Vinter, K.P.; Howe, P.M.; Bond, J.Q.; Reineke, T.M.; Zhang, K.; Dauenhauer, P.J. Multifunctional Cascade Catalysis of Itaconic Acid Hydrodeoxygenation to 3-Methyl-tetrahydrofuran. ACS Sustain. Chem. Eng. 2018, 6, 9394–9402. [Google Scholar] [CrossRef]
- Geilen, F.M.A.; Engendahl, B.; Harwardt, A.; Marquardt, W.; Klankermayer, J.; Leitner, W. Selective and flexible transformation of biomass-derived platform chemicals by a multifunctional catalytic system. Angew. Chem.—Int. Ed. 2010, 49, 5510–5514. [Google Scholar] [CrossRef]
- Geiser, E.; Przybilla, S.K.; Engel, M.; Kleineberg, W.; Büttner, L.; Sarikaya, E.; Hartog, T.D.; Klankermayer, J.; Leitner, W.; Bölker, M.; et al. Genetic and biochemical insights into the itaconate pathway of Ustilago maydis enable enhanced production. Metab. Eng. 2016, 38, 427–435. [Google Scholar] [CrossRef]
- Duetz, W.A.; Rüedi, L.; Hermann, R.; O’Connor, K.; Büchs, J.; Witholt, B. Methods for intense aeration, growth, storage, and replication of bacterial strains in microtiter plates. Appl. Environ. Microbiol. 2000, 66, 2641–2646. [Google Scholar] [CrossRef] [Green Version]
- Maassen, N.; Panakova, M.; Wierckx, N.; Geiser, E.; Zimmermann, M.; Bölker, M.; Klinner, U.; Blank, L.M. Influence of carbon and nitrogen concentration on itaconic acid production by the smut fungus Ustilago maydis. Eng. Life Sci. 2014, 14, 129–134. [Google Scholar] [CrossRef]
- Zambanini, T.; Hartmann, S.K.; Schmitz, L.M.; Büttner, L.; Hosseinpour Tehrani, H.; Geiser, E.; Beudels, M.; Venc, D.; Wandrey, G.; Büchs, J.; et al. Promoters from the itaconate cluster of ustilago maydis are induced by nitrogen depletion. Fungal Biol. Biotechnol. 2017, 4, 11. [Google Scholar] [CrossRef] [Green Version]
- Karaffa, L.; Kubicek, C.P. Production of organic acids by Fungi. In Encyclopedia of Mycology; Springer: Berlin/Heidelberg, Germany, 2021; pp. 406–419. [Google Scholar] [CrossRef]
- Tehrani, H.H.; Saur, K.; Tharmasothirajan, A.; Blank, L.M.; Wierckx, N. Process engineering of pH tolerant Ustilago cynodontis for efficient itaconic acid production. Microb. Cell Fact. 2019, 18, 213. [Google Scholar] [CrossRef] [Green Version]
- Babel, W.; Brinkmann, U.; Müller, R.H. The auxiliary substrate concept—An approach for overcoming limits of microbial performances. Acta Biotechnol. 1993, 13, 211–242. [Google Scholar] [CrossRef]




 g L−1 itaconate) and yield (
g L−1 itaconate) and yield ( g g−1).
g L−1 itaconate) and yield ( g g−1).
g g−1).
g L−1 itaconate) and yield ( g g−1).




| Strain Number | Strain Designation | Resistance | Description | Reference Number |
|---|---|---|---|---|
| #2705 | Ustilago cynodontis | wildtype | NBRC 7530 | |
| #2706 | Ustilago cynodontis | wildtype | NBRC 9727 | |
| #4852 | Ustilago cynodontis | carboxin | Δfuz7 Δcyp3 ΔPria1::Petef | |
| #4853 | Ustilago cynodontis | carboxin | Δfuz7 Δcyp3 ΔPria1::Petef + mttA |
| Condition | Strain | Titermax [g L−1] | YP/Smax [g g−1] | qp, max [g L−1 h−1] |
|---|---|---|---|---|
| 100 g L−1 glucose 0 g L−1 sodium formate 0.8 g L−1 NH4Cl | U. cynodontis Δfuz7 Δcyp3 ↑Pria1 | 50.9 ± 0.3 * | 0.51 ± 0.003 | 0.12 |
| 100 g L−1 glucose 2 g L−1 sodium formate 0.8 g L−1 NH4Cl | 53.9 ± 0.7 * | 0.52 ± 0.007 | 0.14 | |
| 100 g L−1 glucose 0 g L−1 sodium formate 0.8 g L−1 NH4Cl | U. cynodontis Δfuz7 Δcyp3 PetefmttA ↑Pria1 | 50.9 ± 0.4 ** | 0.51 ± 0.004 | 0.12 |
| 100 g L−1 glucose 2 g L−1 sodium formate 0.8 g L−1 NH4Cl | 56.1 ± 0.2 ** | 0.55 ± 0.001 | 0.14 |
| Factor | −1 | 0 | +1 |
|---|---|---|---|
| Glucose [g L−1] | 50 | 165 | 280 |
| Sodium formate [g L−1] | 2.5 | 8.8 | 15 |
| Ammonium chloride [g L−1] | 0.8 | 2.4 | 4 |
| Initial Concentrations | Symbol | Titermax, predicted | Titermax | YP/S,max, predicted | YP/S, max |
|---|---|---|---|---|---|
| [g L−1] | [g L−1] | [g g−1] | [g g−1] | ||
| 238.8 g L−1 glucose, 9.35 g L−1 sodium formate, | ■ | 141.9 ± 8.6 | 138.2 ± 7.0 | 0.57 ± 0.1 | 0.55 ± 0.1 |
| 3.5 g L−1 NH4Cl | |||||
| 95.2 g L−1 glucose, | ▲ | 50.1 ± 5.1 | 52 ± 3.3 | 0.56 ± 0.1 | 0.55 ± 0.1 |
| 2.5 g L−1 sodium formate, | |||||
| 0.8 g L−1 NH4Cl |
| Initial Concentrations | Solvent | Symbol | Titermax [g L−1] | YP/S,max [gITA gsub−1] |
|---|---|---|---|---|
| [gITA gglu−1] | ||||
| 50 g L−1 glucose, 2 g L−1 sodium formate, 0.8 g L−1 NH4Cl | w/o solvent + commercial HCOONa | ▼ | 28.4 ± 0.5 | 0.54 ± 0.1 0.56 ± 0.1 |
| catalyst in tetradecane | ■ | 36.7 ± 0.1 | 0.69 ± 0.1 0.71 ± 0.1 | |
| catalyst in octylacetate | ● | 29.8 ± 0.1 | 0.57 ± 0.1 0.59 ± 0.1 | |
| catalyst in anisole | ♦ | 34.3 ± 0.1 | 0.66 ± 0.1 0.68 ± 0.1 | |
| 100 g L−1 glucose, 4 g L−1 sodium formate, 3.5 g L−1 NH4Cl | w/o solvent + commercial HCOONa | ■ | 63.3 | 0.57 0.59 |
| catalyst in tetradecane | ● | 64.2 | 0.66 0.68 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ullmann, L.; Guntermann, N.; Kohl, P.; Schröders, G.; Müsgens, A.; Franciò, G.; Leitner, W.; Blank, L.M. Improved Itaconate Production with Ustilago cynodontis via Co-Metabolism of CO2-Derived Formate. J. Fungi 2022, 8, 1277. https://doi.org/10.3390/jof8121277
Ullmann L, Guntermann N, Kohl P, Schröders G, Müsgens A, Franciò G, Leitner W, Blank LM. Improved Itaconate Production with Ustilago cynodontis via Co-Metabolism of CO2-Derived Formate. Journal of Fungi. 2022; 8(12):1277. https://doi.org/10.3390/jof8121277
Chicago/Turabian StyleUllmann, Lena, Nils Guntermann, Philipp Kohl, Gereon Schröders, Andreas Müsgens, Giancarlo Franciò, Walter Leitner, and Lars M. Blank. 2022. "Improved Itaconate Production with Ustilago cynodontis via Co-Metabolism of CO2-Derived Formate" Journal of Fungi 8, no. 12: 1277. https://doi.org/10.3390/jof8121277