
Molecular Characterization and Genetic Variation in Ceratocystis fimbriata Ell. and Halst. on Pomegranate
 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Fungal Isolation and Molecular Characterization
2.2. Koch’s Postulate
2.3. Morphological Characterization of the Pathogen
2.4. Fungal Genomic DNA Extraction
2.5. PCR Amplification of Internal Transcribed Spacer (ITS) Region
2.6. Sequencing of rDNA ITS Region and Sequence Analysis of C. fimbriata Isolates
2.7. Genetic Variability Studies of the Pathogenic Isolates
2.8. Band Scoring and Data Analysis
3. Results
3.1. Morphological Characterization of the Pathogen
3.2. Molecular Characterization of Ceratocystis fimbriata Using ITS Primers
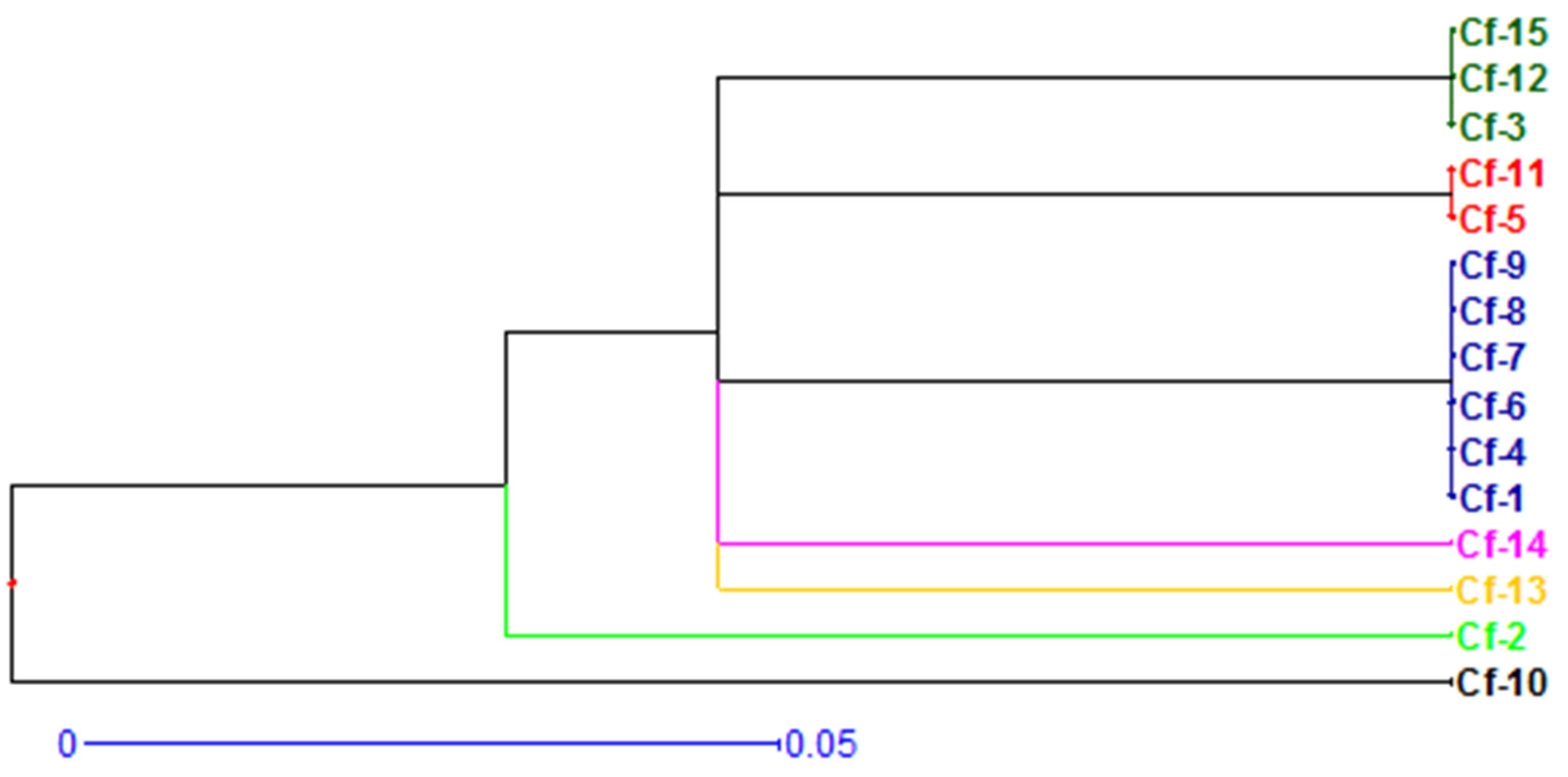
3.3. Genetic Variability Studies of Collected Isolates of Ceratocystis fimbriata
4. Discussion
4.1. Isolation and Identification
4.2. Molecular Characterization of Ceratocystis fimbriata Using ITS Primers
4.3. Genetic Variability Studies of Collected Isolates of Ceratocystis fimbriata
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anonymous. Indian Horticultural Database. 2018. Available online: http://nhb.gov.in (accessed on 30 March 2022).
- Somasekhara, Y.M. Spacious distribution of wilt (Ceratocystis fimbriata Halst. and Halt.) of pomegranate (Punica granatum L.) in India. Res. Crops 2006, 7, 844–853. [Google Scholar]
- Somasekhara, Y.M. New record of Ceratocystis fimbriata causing wilt of Pomegranate in India. Dis. Notes 1999, 83, 400. [Google Scholar] [CrossRef] [PubMed]
- Moller, W.J.; Devay, J.E. Carrot as a species selective isolation medium for Ceratocystis fimbriata. Phytopathology 1968, 58, 123–124. [Google Scholar]
- Decenzo, R.A.; Harrington, T.C. Use of (CAT)5 as a DNA fingerprinting probe for fungi. Phytopathology 1994, 84, 534–540. [Google Scholar] [CrossRef]
- Harrington, T.C.; McNew, D.M.; Steimel, J.; Hofstra, D.; Farrell, R. Phylogeny and taxonomy of the Ophiostoma piceae complex and the Dutch elm disease fungi. Mycologia 2001, 93, 110–135. [Google Scholar] [CrossRef] [Green Version]
- Steimel, J.; Engelbrecht, C.J.B.; Harrington, T.C. Development and characterization of microsatellite markers for the fungus Ceratocystis fimbriata. Mol. Ecol. Notes 2004, 4, 215–218. [Google Scholar] [CrossRef]
- Huang, Q.; Zhu, Y.Y.; Chen, H.R.; Wang, Y.Y.; Lie, Y.L.; Lu, W.J.; Ruan, X.Y. First report of pomegranate wilt caused by Ceratocystis fimbriata in Yunnan, China. Plant Dis. 2003, 87, 1150. [Google Scholar] [CrossRef] [PubMed]
- Fateh, F.S.; Kazmi, M.R.; Ahmad, I.; Ashraf, M. Ceratocystis fimbriata isolated from vascular bundles of declining mango trees in Sindh, Pakistan. Pak. J. Bot. 2006, 38, 1257–1259. [Google Scholar]
- Jadhav, V.T.; Sharma, K.K. Integrated management of diseases in pomegranate. In Proceedings of the 2nd International Symposium on Pomegranate and Minor, Including Mediterranean Fruits, Dharwad, India, 23–27 June 2009; University of Agricultural Sciences: Dharwad, India, 2009; pp. 48–52. [Google Scholar]
- Xu, B.; Zheng, X.H.; Guo, W.X.; Zhou, X.P.; He, P. First report of pomegranate wilt caused by Ceratocystis fimbriata in Sichuan Province. Plant Dis. 2011, 95, 776. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, V.G.; Kshirsagar, P.; Tirmali, A.M. Studies on wilt complex disease of pomegranate (Punica granatum L.). Adv. Life Sci. 2016, 5, 747–755. [Google Scholar]
- Raja, S.G.; Amaresh, Y.S.; Yenjerappa, S.T.; Amaregouda, A.; Shreenivas, A.G. Morphological characteristics of Ceratocystis fimbriata Ell. and Halst. causing wilt in pomegranate. Int. J. Pure Appl. Biosci. 2017, 5, 285–289. [Google Scholar]
- Raja. Investigations on wilt of pomegranate caused by Ceratocystis fimbriata Ell. and Halst. in Karnataka. Ph.D. Thesis, University of Agricultural Sciences, Raichur, India, 2017. [Google Scholar]
- Somu, R. Etiology, Epidemiology and Management of Wilt of Pomegranate Caused by Ceratocystis fimbriata Ellis and Halst. Ph.D. Thesis, University of Horticultural Sciences Bagalkot, Bagalkot, India, 2017. [Google Scholar]
- Anonymous. Annual Report (2016–2017); NRCP: Solapur, India, 2016; p. 39. [Google Scholar]
- Alam, M.W.; Gleason, M.L.; Mehboob, S.; Riaz, K.; Rehman, A. First report of Ceratocystis fimbriata causing pomegranate wilt in Pakistan. Plant Dis. 2016, 101, 251. [Google Scholar] [CrossRef]
- Singh, A.; Singh, U.S.; Willocquet, L.; Savary, S. Relationship among cultural morphological characteristics, anastomosis behavior and pathogenicity of Rhizoctonia solani Khun on rice. J. Mycol. Plant Pathol. 1999, 29, 306–316. [Google Scholar]
- Santini, A.; Capretti, P. Analysis of the Italian population of Ceratocystis fimbriata f. sp. platani using RAPD and minisatellite markers. Plant Pathol. 2000, 49, 461–467. [Google Scholar] [CrossRef] [Green Version]
- Scruggs, A.C.; Isaiah, T.; Adams, M.L.; Quesada-Ocampo, L.M. Genetic diversity, fungicide sensitivity and host resistance to Ceratocystis fimbriata infecting sweet potato in North Carolina. Plant Dis. 2017, 101, 994–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrington, T.C.; McNew, D.L. Self-fertility and uni-directional mating-type switching in Ceratocystis coerulescens, a filamentous ascomycete. Curr. Genet. 1997, 32, 9–52. [Google Scholar] [CrossRef] [PubMed]
- Harrington, T.C.; Steimel, J.; Kile, G. Genetic variation in three Ceratocystis species with outcrossing, selfing and asexual reproductive strategies. Eur. J. Forest Pathol. 1998, 28, 217–226. [Google Scholar] [CrossRef]
- Engelbrecht, C.J.B.; Harrington, T.C.; Steimel, J.; Capretti, P. Genetic variation in eastern North American and putatively introduced populations of Ceratocystis fimbriata f. sp. platani. Mol. Ecol. 2004, 13, 2995–3005. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Sl. No. | Isolate | Name of the Place | ||
|---|---|---|---|---|
| Village | Taluk | District | ||
| 1 | Cf-1 | Pura | Devanahalli | Rural Bengaluru |
| 2 | Cf-2 | Harohalli | ||
| 3 | Cf-3 | Vijayapura | ||
| 4 | Cf-4 | Yaluvahalli | ||
| 5 | Cf-5 | Kalenahalli | North Bengaluru | Bengaluru |
| 6 | Cf-6 | GKVK, Hebbal | ||
| 7 | Cf-7 | Uyyalappanahalli | Kanakapura | Ramanagara |
| 8 | Cf-8 | Jakkalli | Kollegala | Chamarajanagara |
| 9 | Cf-9 | Rangapura | Sira | Tumakuru |
| 10 | Cf-10 | Thogaragunte | ||
| 11 | Cf-11 | Dharmapura | Hiriyur | Chitradurga |
| 12 | Cf-12 | Javagondanahalli | ||
| 13 | Cf-13 | Maskal | ||
| 14 | Cf-14 | Ramajjanahalli | Hosadurga | |
| 15 | Cf-15 | Kurubarahalli | ||
| Sl. No. | Isolate | Colony Colour | Type of Colony Growth | Type of Margin | Growth Rate (cm) | Days Taken for Complete Growth | |||
|---|---|---|---|---|---|---|---|---|---|
| Growth at 10 DAIs | Growth at 15 DAIs | Growth at 20 DAIs | Growth at 25 DAIs | ||||||
| 1 | Cf-1 | Whitish grey | Flat | Uniform | 3.63 | 6.13 | 7.13 | 7.83 | 30 |
| 2 | Cf-2 | Light grey | Flat | Uniform | 5.76 | 8.60 | 9.00 | 9.00 | 19 |
| 3 | Cf-3 | Grey | Flat | Uniform | 4.06 | 6.67 | 9.00 | 9.00 | 20 |
| 4 | Cf-4 | Greyish | Flat | Uniform | 5.23 | 8.30 | 8.73 | 9.00 | 23 |
| 5 | Cf-5 | Grey | Flat | Uniform | 6.80 | 9.00 | 9.00 | 9.00 | 15 |
| 6 | Cf-6 | Dark grey | Flat | Uniform | 4.33 | 7.26 | 8.32 | 9.00 | 24 |
| 7 | Cf-7 | Greyish | Flat | Irregular | 4.53 | 7.36 | 7.98 | 9.00 | 26 |
| 8 | Cf-8 | Grey | Flat | Uniform | 4.93 | 8.23 | 9.00 | 9.00 | 18 |
| 9 | Cf-9 | Light grey | Flat | Uniform | 6.20 | 8.83 | 8.92 | 9.00 | 23 |
| 10 | Cf-10 | Dark grey | Flat | Uniform | 4.20 | 7.50 | 9.00 | 9.00 | 19 |
| 11 | Cf-11 | Grey | Flat | Uniform | 5.20 | 7.00 | 8.60 | 9.00 | 22 |
| 12 | Cf-12 | Grey | Flat | Uniform | 4.86 | 7.13 | 8.86 | 9.00 | 21 |
| 13 | Cf-13 | Dark grey | Flat | Uniform | 2.63 | 3.83 | 5.33 | 6.46 | 30 |
| 14 | Cf-14 | Greenish grey | Flat | Uniform | 4.30 | 5.26 | 5.91 | 6.56 | 35 |
| 15 | Cf-15 | Grey | Flat | Uniform | 5.96 | 8.80 | 9.00 | 9.00 | 17 |
| S.Em ± | 1.64 | ||||||||
| CD @ 1% | 4.81 | ||||||||
| Sl. No. | Isolate | Closest Match | Country | Query Cover (%) | Percent Similarity (%) | Query Length (bp) | Accession No. |
|---|---|---|---|---|---|---|---|
| 1 | Cf-4 | C. fimbriata isolate C2174 | China | 99 | 98.43 | 656 | KY580844.1 |
| C. fimbriata | China | 99 | 98.42 | 675 | KY440175.1 | ||
| C. fimbriata HgK5 | China | 98 | 98.88 | 769 | MH535909.1 | ||
| C. fimbriata isolate 4-8SYWPGYP | China | 97 | 99.03 | 759 | MH535923.1 | ||
| C. fimbriata isolate C2870 | China | 99 | 98.42 | 648 | KY580869.1 | ||
| 2 | Cf-5 | C. fimbriata voucher MG-2-10 | China | 96 | 90.10 | 660 | KX101051.1 |
| C. fimbriata voucher MG-2-8 | China | 96 | 90.10 | 640 | KX101050.1 | ||
| C. fimbriata voucher MG-2-4 | China | 96 | 90.10 | 651 | KX101049.1 | ||
| C. fimbriata voucher MG-2-1 | China | 96 | 90.10 | 652 | KX101048.1 | ||
| C. fimbriata YM1-2 | China | 96 | 89.95 | 635 | MH793674.1 | ||
| 3 | Cf-7 | C. fimbriata isolate C2866 | China | 98 | 98.42 | 656 | KY580865.1 |
| C. fimbriata isolate C2864 | China | 98 | 98.43 | 665 | KY580863.1 | ||
| C. fimbriata isolate C2847 | China | 98 | 98.43 | 666 | KY580857.1 | ||
| C. fimbriata isolate C2862 | China | 98 | 98.42 | 653 | KY580861.1 | ||
| C. fimbriata isolate HF-28 | China | 98 | 98.43 | 665 | KT963162.1 | ||
| 4 | Cf-8 | C. fimbriata isolate AM6-2SYP-GYP | China | 96 | 99.52 | 759 | MH535926.1 |
| C. fimbriata isolate YW2-3 | China | 96 | 99.52 | 635 | MH793676.1 | ||
| C. fimbriata isolate YW3-1 | China | 95 | 99.51 | 629 | MH793677.1 | ||
| C. fimbriata isolate C3-1GYP | China | 95 | 99.51 | 627 | MH540139.1 | ||
| C. fimbriata isolate NRCP-CF22 | Bagalkote, India | 97 | 98.72 | 623 | KU877210.1 | ||
| C. fimbriata isolate NRCP-CF24 | Bagalkote, India | 96 | 99.03 | 623 | KU877211.1 | ||
| 5 | Cf-9 | C. fimbriata isolate C2174 | China | 99 | 99.06 | 656 | KY580844.1 |
| C. fimbriata isolate C2864 | China | 98 | 99.06 | 665 | KY580863.1 | ||
| C. fimbriata isolate C2847 | China | 98 | 99.06 | 666 | KY580857.1 | ||
| C. fimbriata isolate C2866 | China | 99 | 98.90 | 656 | KY580865.1 | ||
| C. fimbriata isolate C2864 | China | 99 | 98.90 | 665 | KY580863.1 | ||
| 6 | Cf-11 | C. fimbriata voucher MG-2-10 | China | 97 | 99.52 | 660 | KX101051.1 |
| C. fimbriata voucher MG-2-8 | China | 97 | 99.68 | 640 | KX101050.1 | ||
| C. fimbriata voucher MG-2-4 | China | 97 | 99.52 | 651 | KX101049.1 | ||
| C. fimbriata voucher MG-2-1 | China | 97 | 99.52 | 652 | KX101048.1 | ||
| C. fimbriata isolate C2868 | China | 97 | 99.36 | 653 | KY580867.1 |
| Sl. No. | Isolates | Districts | Accession Numbers |
|---|---|---|---|
| 1 | Cf-4 (ITS 1) | Bengaluru Rural | MZ749649 |
| Cf-4 (ITS 4) | MZ749650 | ||
| 2 | Cf-5 (ITS 1) | Bengaluru Urban | MZ749663 |
| Cf-5 (ITS 4) | MZ749664 | ||
| 3 | Cf-7 (ITS 1) | Ramanagara | MZ749665 |
| Cf-7 (ITS 4) | MZ749666 | ||
| 4 | Cf-8 (ITS 1) | Chamarajanagara | MZ749671 |
| Cf-8 (ITS 4) | MZ749672 | ||
| 5 | Cf-9 (ITS 1) | Tumakuru | MZ749741 |
| Cf-9 (ITS 4) | MZ749742 | ||
| 6 | Cf-11 (ITS 1) | Chitradurga | MZ749730 |
| Cf-11 (ITS 4) | MZ749731 |
| Cf-1 | Cf-2 | Cf-3 | Cf-4 | Cf-5 | Cf-6 | Cf-7 | Cf-8 | Cf-9 | Cf-10 | Cf-11 | Cf-12 | Cf-13 | Cf-14 | Cf-15 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cf-1 | 1.00 | ||||||||||||||
| Cf-2 | 0.13 | 1.00 | |||||||||||||
| Cf-3 | 0.10 | 0.13 | 1.00 | ||||||||||||
| Cf-4 | 0.00 | 0.13 | 0.10 | 1.00 | |||||||||||
| Cf-5 | 0.10 | 0.13 | 0.10 | 0.10 | 1.00 | ||||||||||
| Cf-6 | 0.00 | 0.13 | 0.10 | 0.00 | 0.10 | 1.00 | |||||||||
| Cf-7 | 0.00 | 0.13 | 0.10 | 0.00 | 0.10 | 0.00 | 1.00 | ||||||||
| Cf-8 | 0.00 | 0.13 | 0.10 | 0.00 | 0.10 | 0.00 | 0.00 | 1.00 | |||||||
| Cf-9 | 0.00 | 0.13 | 0.10 | 0.00 | 0.10 | 0.00 | 0.00 | 0.00 | 1.00 | ||||||
| Cf-10 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 1.00 | |||||
| Cf-11 | 0.10 | 0.13 | 0.10 | 0.10 | 0.00 | 0.10 | 0.10 | 0.10 | 0.10 | 0.20 | 1.00 | ||||
| Cf-12 | 0.10 | 0.13 | 0.00 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | 0.20 | 0.10 | 1.00 | |||
| Cf-13 | 0.10 | 0.13 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | 0.20 | 0.10 | 0.10 | 1.00 | ||
| Cf-14 | 0.10 | 0.13 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | 0.20 | 0.10 | 0.10 | 0.10 | 1.00 | |
| Cf-15 | 0.10 | 0.13 | 0.00 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | 0.20 | 0.10 | 0.00 | 0.10 | 0.10 | 1.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Erakyathappa, N.S.; Venkatappa, D.; Gopal, S.C.; Sridhara, S.; Kalaiah, H.M.; Ugalat, J. Molecular Characterization and Genetic Variation in Ceratocystis fimbriata Ell. and Halst. on Pomegranate. J. Fungi 2022, 8, 1276. https://doi.org/10.3390/jof8121276
Erakyathappa NS, Venkatappa D, Gopal SC, Sridhara S, Kalaiah HM, Ugalat J. Molecular Characterization and Genetic Variation in Ceratocystis fimbriata Ell. and Halst. on Pomegranate. Journal of Fungi. 2022; 8(12):1276. https://doi.org/10.3390/jof8121276
Chicago/Turabian StyleErakyathappa, Navyashree Sailapura, Devappa Venkatappa, Sangeetha Chittarada Gopal, Shankarappa Sridhara, Honnabyraiah Madhapura Kalaiah, and Jayashree Ugalat. 2022. "Molecular Characterization and Genetic Variation in Ceratocystis fimbriata Ell. and Halst. on Pomegranate" Journal of Fungi 8, no. 12: 1276. https://doi.org/10.3390/jof8121276