
Utilization of Corn Steep Liquor for the Production of Fairy Chemicals by Lepista sordida Mycelia

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Material
2.2. Compositions of the Media
2.3. Culture Conditions
2.4. Quantification of the FCs
2.5. Statistical Analysis
3. Results
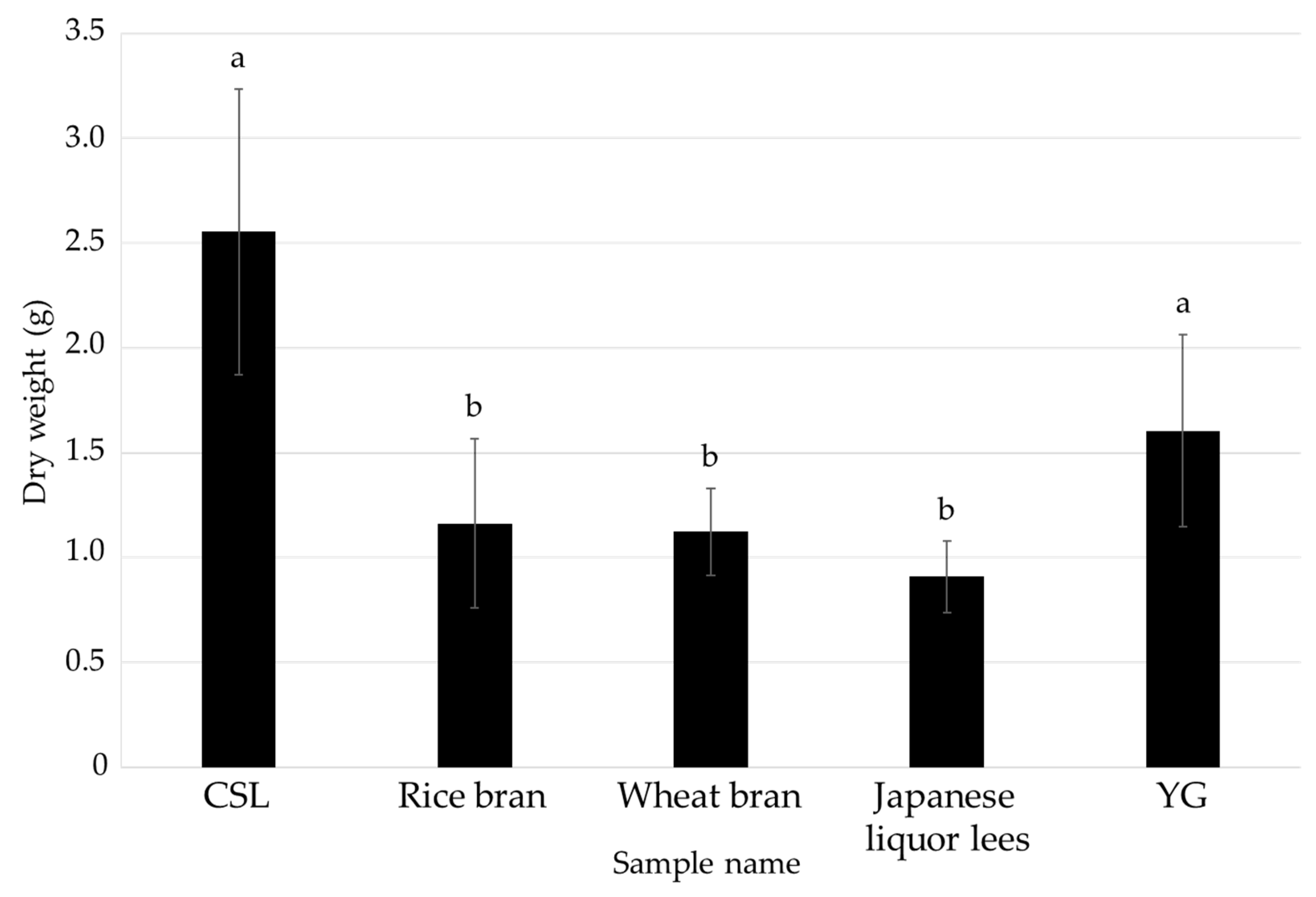
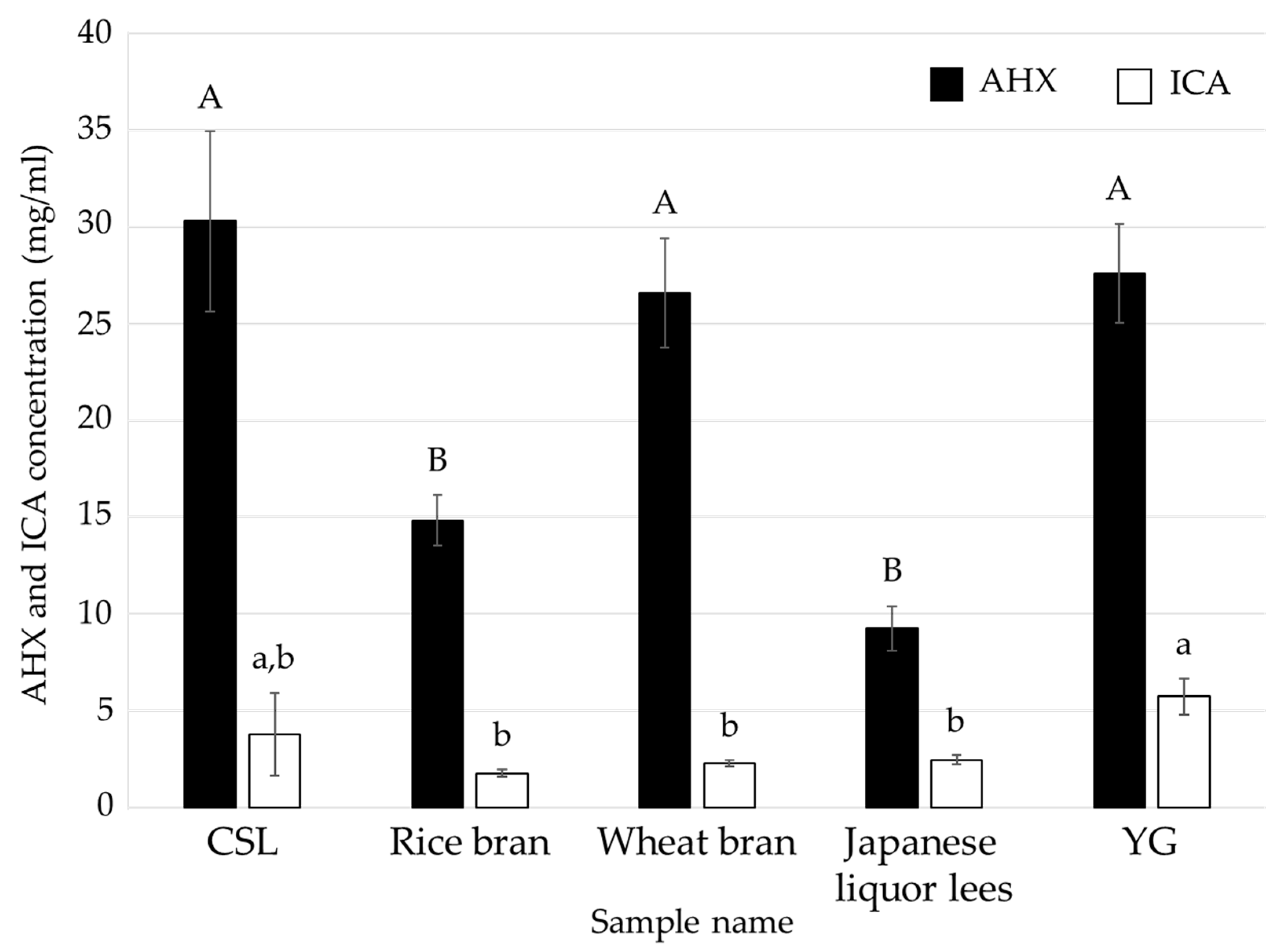
3.1. Effects of Food Industrial By-Products as Nutritional Additives on the Mycelial Cultivation of L. sordida
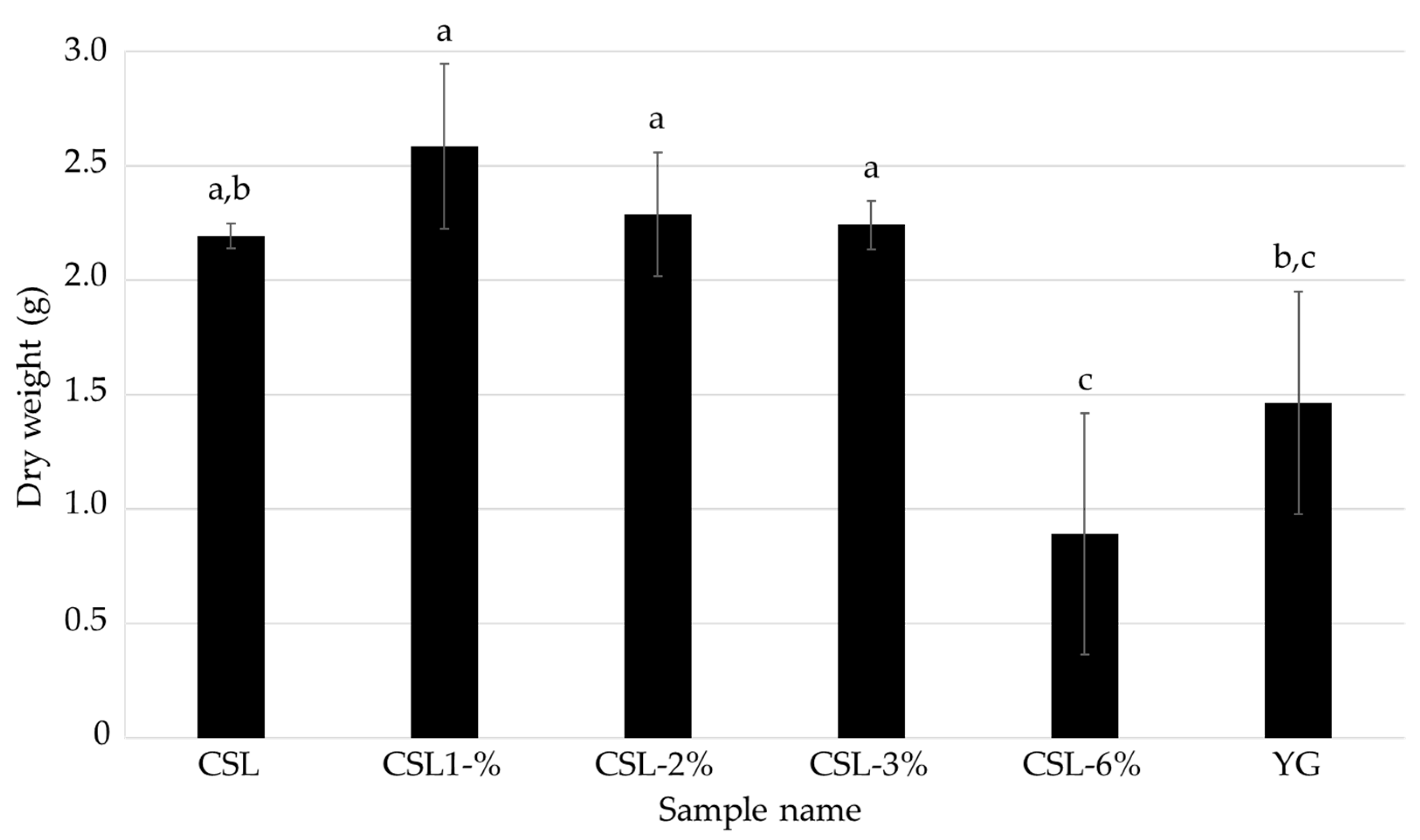
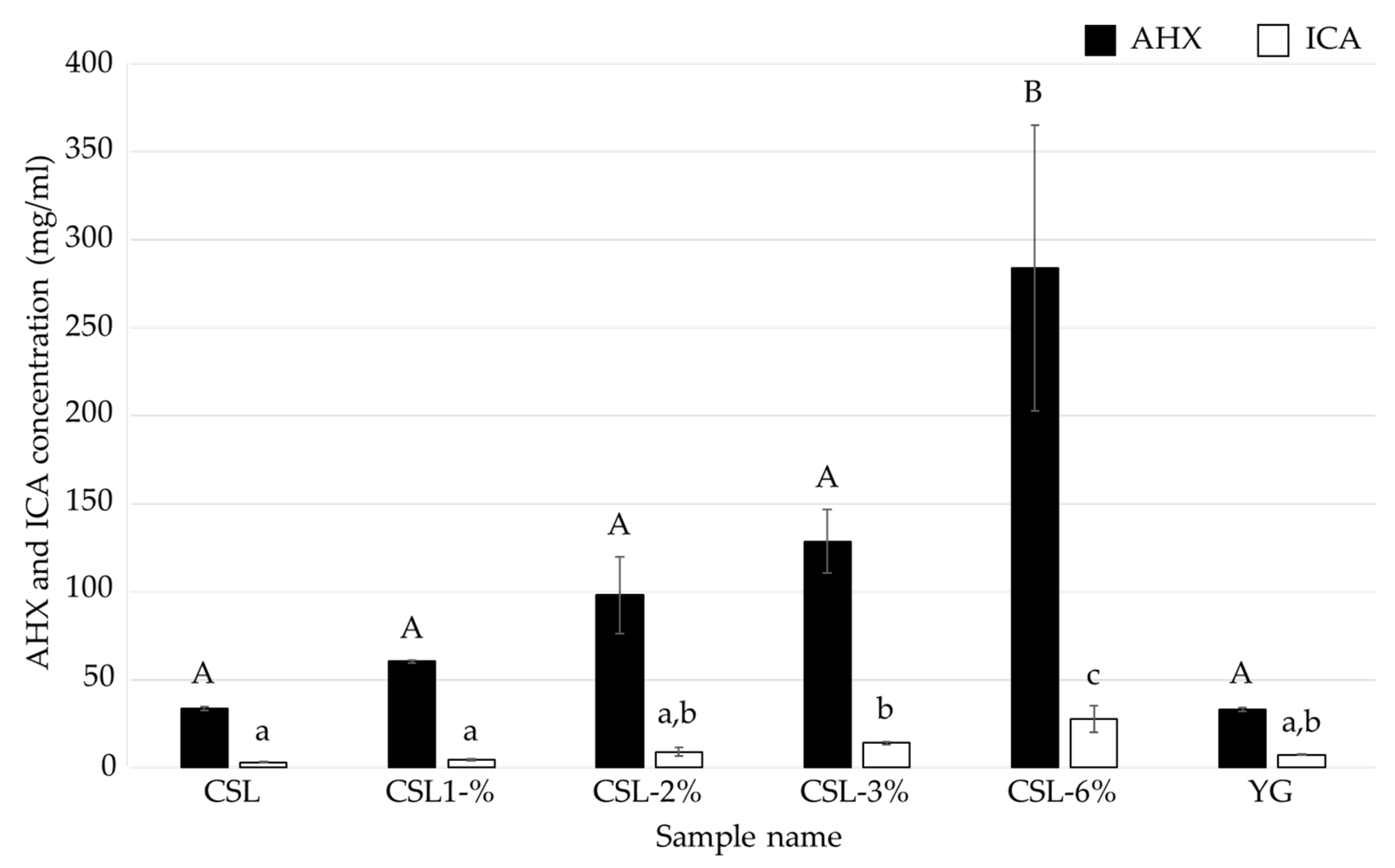
3.2. Investigation of the Optimal Concentration of CSL as a Nutritional Additive for the Mycelial Cultivation of L. sordida
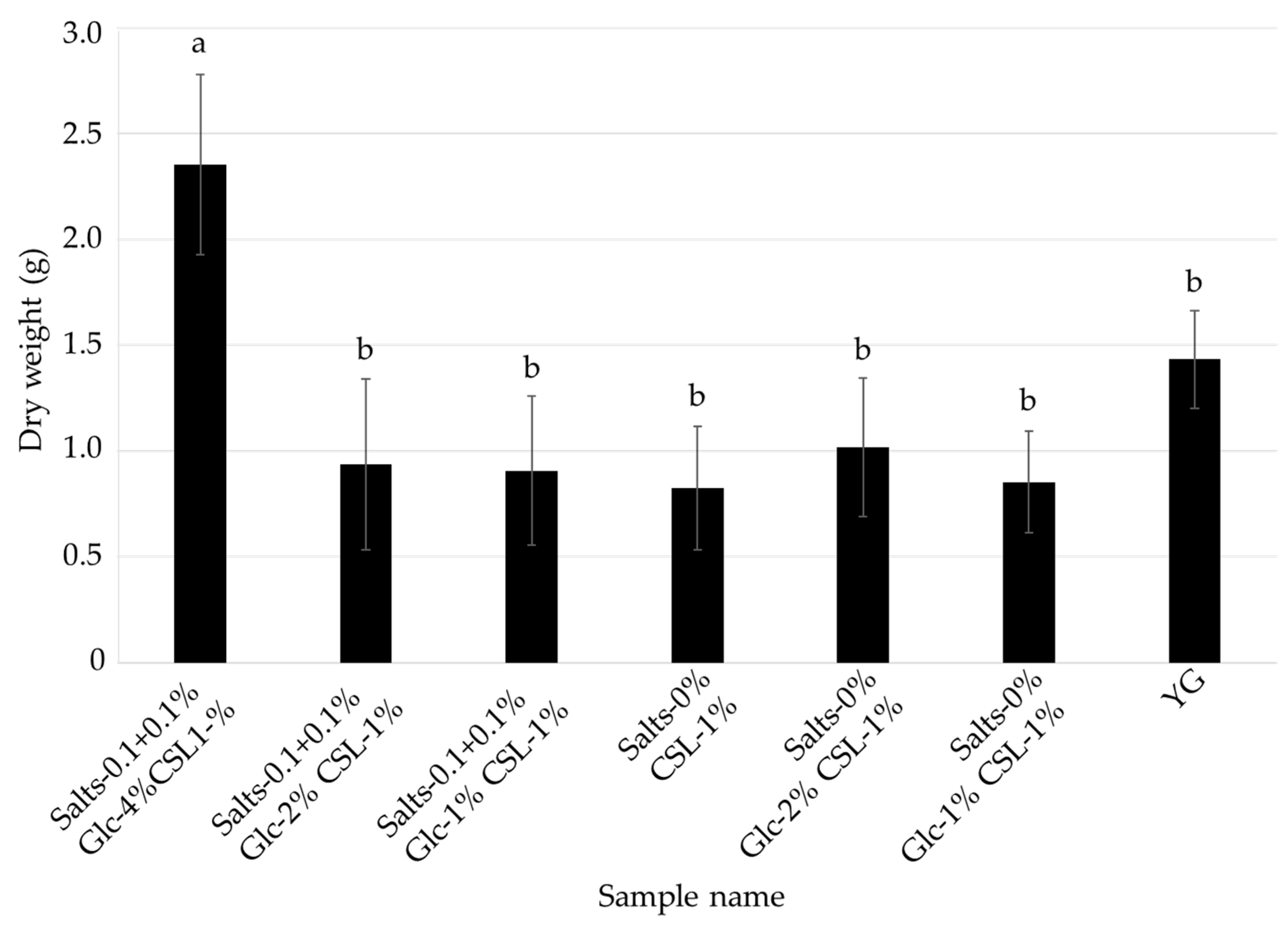
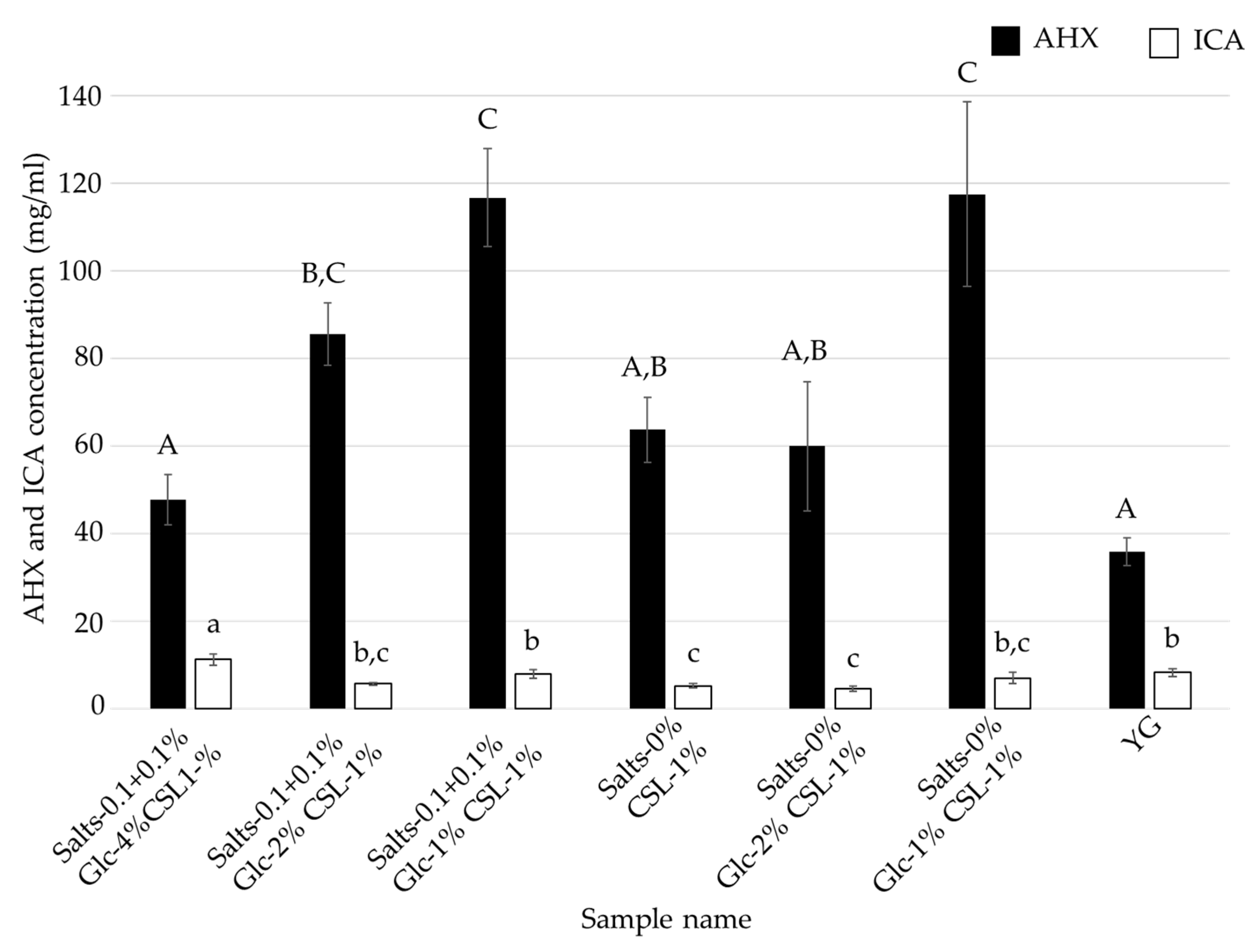
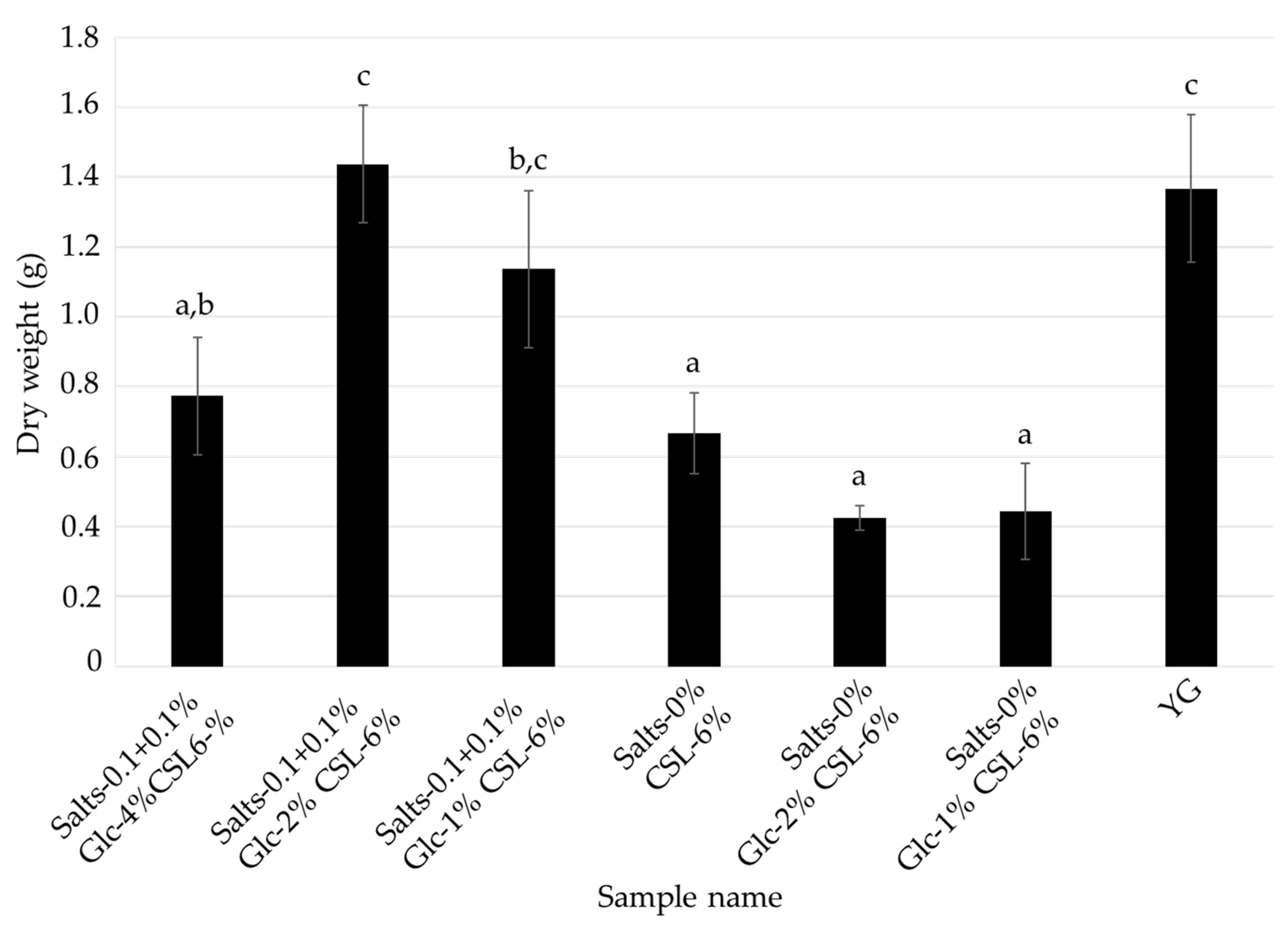
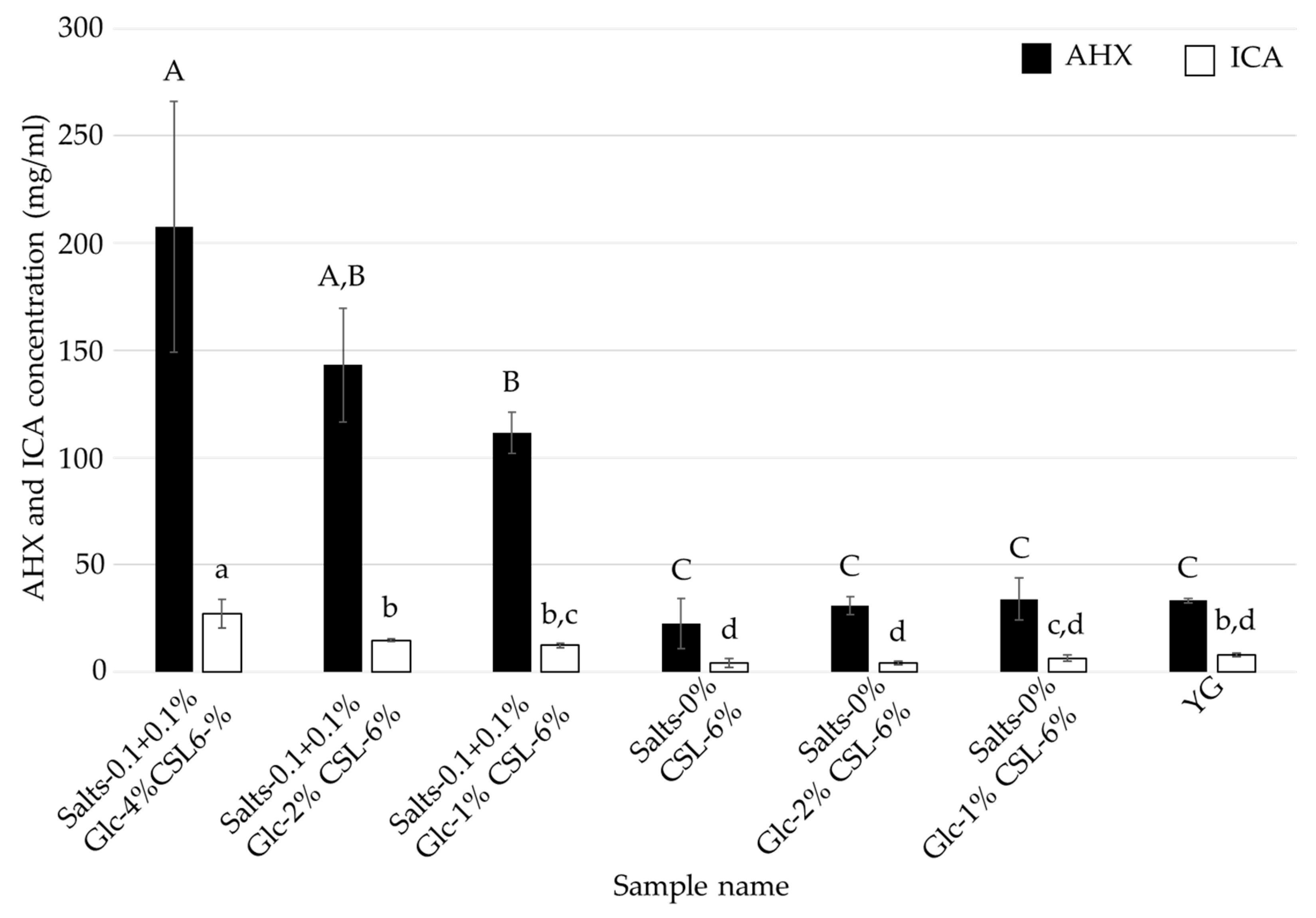
3.3. Effects of Glucose and Inorganic Salts on the Mycelial Cultivation of L. sordida
4. Discussion
4.1. CSL as a Nutritional Additive in Liquid Media Is Effective for the Culturing of L. sordida Mycelia
4.2. Speculation as to Why CSL Increased the Mycelial Yield, and AHX and ICA Contents
4.3. Efficient Mycelial Cultivation Method for L. sordida According to the Purpose
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shiferaw, B.; Prasanna, B.M.; Hellin, J.; Bänziger, M. Crops that feed the world 6. Past successes and future challenges to the role played by maize in global food security. Food Secur. 2011, 3, 307–327. [Google Scholar] [CrossRef] [Green Version]
- Iwade, I. Culture Methods of Mushrooms; Chikyusha: Tokyo, Japan, 1959; pp. 116–117. (In Japanese) [Google Scholar]
- Harada, E.; Nagatomi, F.; Kijidani, Y.; Meguro, S.; Kammera, T. Utilization of soba-shochu distillery waste for benzaldehyde and ergothioneine production from liquid culture of the edible basidiomycete Grifola gargal. Mushroom Sci. Biotechnol. 2014, 21, 165–171. [Google Scholar] [CrossRef]
- Mitchinson, A. Fairy chemicals. Nature 2014, 505, 298. [Google Scholar] [CrossRef]
- Choi, J.-H.; Fushimi, K.; Abe, N.; Tanaka, H.; Maeda, S.; Morita, A.; Hara, M.; Motohashi, R.; Matsuaga, J.; Eguchi, Y.; et al. Disclosure of the “fairy” of fairy-ring-forming fungus Lepista sordida. ChemBioChem 2010, 11, 1373–1377. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.-H.; Abe, N.; Tanaka, H.; Fushimi, K.; Nishina, Y.; Morita, A.; Kiriiwa, Y.; Motohashi, R.; Hashizume, D.; Koshino, H.; et al. Plant-growth regulator, imidazole-4-carboxamide, produced by the fairy ring forming fungus Lepista sordida. J. Agric. Food Chem. 2010, 58, 9956–9959. [Google Scholar] [CrossRef] [PubMed]
- Beal, D.D.; Skibba, J.L.; Croft, W.A.; Cohen, S.M.; Bryan, G.T. Carcinogenicity of the antineoplastic agent, 5-(3,3-dimethyl-1-triazeno)-imidazole-4-carboxamide, and its metabolites in rats. J. Natl. Cancer Inst. 1975, 54, 951–957. [Google Scholar] [CrossRef]
- Vaughan, K.; Wilman, D.E.V.; Wheelhouse, R.T.; Stevens, M.F.G. A 15N NMR investigation of a series of benzotriazinones and related antitumour heterocycles. Magn. Reson. Chem. 2002, 40, 300–302. [Google Scholar] [CrossRef]
- Leone-Bay, A.; Glaser, L. An efficient method for the preparation of 4(5)-cyanoimidazole. Synth. Commun. 1987, 17, 1409–1412. [Google Scholar] [CrossRef]
- Kanzawa, F.; Hoshi, A.; Kuretani, K. Inhibition of guanine deaminase with derivatives of imidazole. Chem. Pharm. Bull. 1971, 19, 1737–1738. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.-H.; Ohnishi, T.; Yamakawa, Y.; Takeda, S.; Sekiguchi, S.; Maruyama, W.; Yamashita, K.; Suzuki, T.; Morita, A.; Ikka, T.; et al. The source of “fairy rings”: 2-azahypoxanthine and its metabolite found in a novel purine metabolic pathway in plants. Angew. Chem. Int. Ed. 2014, 53, 1552–1555. [Google Scholar] [CrossRef]
- Kawagishi, H. Fairy chemicals—A candidate for a new family of plant hormones and possibility of practical use in agriculture. Biosci. Biotechnol. Biochem. 2018, 82, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Kawagishi, H. Are fairy chemicals a new family of plant hormones? Proc. Jpn. Acad. Ser. B 2019, 95, 29–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asai, T.; Choi, J.-H.; Ikka, T.; Fushimi, K.; Abe, N.; Tanaka, H.; Yamakawa, Y.; Kobori, H.; Kiriiwa, Y.; Motohashi, R.; et al. Effect of 2-azahypoxanthine (AHX) produced by the fairy-ring-forming fungus on the growth and the grain yield of rice. Jpn. Agric. Res. Q. 2015, 49, 45–49. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.; Miwa, Y.; Wu, J.; Shoda, C.; Jeong, H.; Kawagishi, H.; Tsubota, K.; Kurihara, T. A fairy chemical suppresses retinal angiogenesis as a HIF inhibitor. Biomolecules 2020, 10, 1405. [Google Scholar] [CrossRef] [PubMed]
- Inoue, C.; Yasuma, T.; D’Alessandro-Gabazza, C.N.; Toda, M.; D’Alessandro, V.F.; Inoue, R.; Fujimoto, H.; Kobori, H.; Tharavecharak, S.; Takeshita, A.; et al. The fairy chemical imidazole-4-carboxamide inhibits the expression of Axl, PD-L1, and PD-L2 and improves response to cisplatin in melanoma. Cells 2022, 11, 374. [Google Scholar] [CrossRef] [PubMed]
- Aoshima, H.; Ito, M.; Ibuki, R.; Kawagishi, H. Potential of fairy chemicals as functional cosmetic ingredients: Effect of 2-aza-8-oxohypoxanthine (AOH) on skin lightness. Int. J. Med. Mushrooms 2022, 24, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Aoshima, H.; Ito, M.; Ibuki, R.; Kawagishi, H. The potential of 2-aza-8-oxohypoxanthine as a cosmetic ingredient. Cosmetics 2021, 8, 60. [Google Scholar] [CrossRef]
- Aoshima, H.; Ibuki, R.; Ito, M.; Kawagishi, H. Clinical evaluation of topical lotion containing 2-aza-8-oxohypoxanthine on skin barrier function against water loss. Cosmetics 2021, 8, 83. [Google Scholar] [CrossRef]
- Aoshima, H.; Matsumoto, T.; Ibuki, R.; Kawagishi, H. Safety evaluation of 2-aza-8-oxohypoxanthine by in vitro skin sensitization and human tests. Fundam. Toxicol. Sci. 2021, 8, 123–133. [Google Scholar] [CrossRef]
- Aoshima, H.; Hyodo, S.; Ibuki, R.; Wu, J.; Choi, J.-H.; Kawagishi, H. Safety evaluation of 2-aza-8-oxohypoxanthine based on in vitro and human patch tests. Fundam. Toxicol. Sci. 2020, 7, 207–214. [Google Scholar] [CrossRef]
- Choi, J.-H.; Kikuchi, A.; Pumkaeo, P.; Hirai, H.; Tokuyama, S.; Kawagishi, H. Bioconversion of AHX to AOH by resting cells of Burkholderia contaminans CH-1. Biosci. Biotechnol. Biochem. 2016, 80, 2045–2050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takemura, H.; Choi, J.-H.; Matsuzaki, N.; Taniguchi, Y.; Wu, J.; Hirai, H.; Motohashi, R.; Asakawa, T.; Ikeuchi, K.; Inai, M.; et al. A fairy chemical, imidazole-4-carboxamide, is produced on a novel purine metabolic pathway in rice. Sci. Rep. 2019, 9, 9899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taiwo, A.E.; Madzimbamuto, T.N.; Ojumu, T.V. Optimization of corn steep liquor dosage and other fermentation parameters for ethanol production by Saccharomyces cerevisiae type 1 and anchor instant yeast. Energies 2018, 11, 1740. [Google Scholar] [CrossRef] [Green Version]
- Perlman, D. The evolution of penicillin manufacturing processes. In the history of penicillin production. Chem. Eng. Prog. Symp. Ser. 1970, 60, 25–30. [Google Scholar]
- Harned, R.L. Riboflavine Production. U.S. Patent 3,475,274, 28 October 1969. [Google Scholar]
- Lee, P.; Lee, W.; Lee, S.; Chang, H.; Chang, Y. Fermentative production of succinic acid from glucose and corn steep liquor by Anaerobiospirillum succiniciproducens. Biotechnol. Bioprocess Eng. 2000, 5, 379–381. [Google Scholar] [CrossRef]
- Abe, M.; Iida, S.; Ohga, S. Supplemental effect of corn steep liquor in a sawdust−based substrate of Shiitake (Lentinuta edodes). Jpn. Soc. Mushroom Sci. Biotechnol. 2002, 10, 29–34. [Google Scholar] [CrossRef]
- Maleki-Kakelar, M.; Azarhoosh, M.J.; Senji, S.J.; Aghaeinejad-Meybodi, A. Urease production using corn steep liquor as a low-cost nutrient source by Sporosarcina pasteurii: Biocementation and process optimization via artificial intelligence approaches. Environ. Sci. Pollut. Res. 2022, 29, 13767–13781. [Google Scholar] [CrossRef]
- Nakano, A.; Uehara, Y.; Yamauchi, A. Effects of organic liquid fertilizer (corn steep liquor) on initial growth and rhizosphere of tomato plant. Environ. Control. Biol. 2000, 38, 211–219. [Google Scholar] [CrossRef] [Green Version]
- Matsushima, H.; Fukumi, H.; Arima, K. Isolation of zeanic acid, a natural plant growth-regulator from corn steep liquor and its chemical structure. Agric. Biol. Chem. 1973, 37, 1865–1871. [Google Scholar] [CrossRef]
- Matsushima, H.; Arima, K. Physiological activities of zeanic acid, a new plant-growth promotor from corn steep liquor. Agric. Biol. Chem. 1973, 37, 1873–1880. [Google Scholar] [CrossRef]
- Adio, A.M.; Casteel, C.L.; Vos, M.D.; Kim, J.-H.; Joshi, V.; Li, B.; Juéry, C.; Daron, J.; Kliebenstein, D.J.; Jander, G. Biosynthesis and defensive function of N Δ-acetylornithine, a jasmonate-induced Arabidopsis Metabolite. Plant Cell 2011, 23, 3303–3318. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Aboshi, T.; Teraishi, M.; Strickler, S.R.; Spindel, J.E.; Tung, C.-W.; Takata, R.; Matsumoto, F.; Maesaka, Y.; McCouch, S.R.; et al. The tyrosine aminomutase TAM1 is required for b-tyrosine biosynthesis in rice. Plant Cell 2015, 27, 1265–1278. [Google Scholar] [CrossRef] [Green Version]
- Alamgir, K.M.; Hojo, Y.; Christeller, J.T.; Fukumoto, K.; Isshiki, R.; Shinya, T.; Baldwin, I.T.; Galis, I. Systematic analysis of rice (Oryza sativa) metabolic responses to herbivory. Plant Cell Environ. 2016, 39, 453–466. [Google Scholar] [CrossRef] [PubMed]
- Schmelz, E.A.; Kaplan, F.; Huffaker, A.; Dafoe, N.J.; Vaughan, M.M.; Ni, X.; Rocca, J.R.; Alborn, H.T.; Teal, P.E. Identity, regulation, and activity of inducible diterpenoid phytoalexins in maize. Proc. Natl. Acad. Sci. USA 2011, 108, 5455–5460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huffaker, A.; Kaplan, F.; Vaughan, M.M.; Dafoe, N.J.; Ni, X.; Rocca, J.R.; Alborn, H.T.; Teal, P.E.; Schmelz, E.A. Novel acidic sesquiterpenoids constitute a dominant class of pathogen-induced phytoalexins in maize. Plant Physiol. 2011, 156, 2082–2097. [Google Scholar] [CrossRef] [Green Version]
- Christensen, S.A.; Huffaker, A.; Kaplan, F.; Sims, J.; Ziemann, S.; Doehlemann, G.; Ji, L.; Schmitz, R.J.; Kolomiets, M.V.; Alborn, H.T.; et al. Maize death acids, 9-lipoxygenase–derived cyclopente(a)nones, display activity as cytotoxic phytoalexins and transcriptional mediators. Proc. Natl. Acad. Sci. USA 2015, 112, 11407–11412. [Google Scholar] [CrossRef] [Green Version]
- Ito, A.; Choi, J.-H.; Takemura, H.; Kotajima, M.; Wu, J.; Tokuyama, S.; Hirai, H.; Asakawa, T.; Ouchi, H.; Inai, M.; et al. Biosynthesis of the fairy chemicals, 2-azahypoxanthine and imidazole-4-carboxamide, in the fairy ring-forming fungus Lepista sordida. J. Nat. Prod. 2020, 83, 2469–2476. [Google Scholar] [CrossRef]
- Chinta, Y.D.; Kano, K.; Widiastuti, A.; Fukahori, M.; Kawasaki, S.; Eguchi, Y.; Misu, H.; Odani, H.; Zhou, S.; Narisawa, K.; et al. Effect of corn steep liquor on lettuce root rot (Fusarium oxysporum f.sp. lactucae) in hydroponic cultures. J. Sci. Food Agric. 2014, 94, 2317–2323. [Google Scholar] [CrossRef]
- Sharma, R.K.; Kothari, R.M. Innovative application of corn steep liquor for the increased production of food grains. Technovation 1992, 12, 213–221. [Google Scholar] [CrossRef]
- Fox, J.E.; Miller, C. Factors in corn steep water promoting growth of plant tissues. Plant Physiol. 1959, 34, 577–579. [Google Scholar] [CrossRef] [Green Version]
- Zhou, K.; Yu, J.; Ma, Y.; Cai, L.; Zheng, L.; Gong, W.; Liu, Q.-A. Corn steep liquor: Green biological resources for bioindustry. Appl. Biochem. Biotechnol. 2022, 194, 3280–3295. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CSL | Rice | Wheat | Japanese | Control | |
|---|---|---|---|---|---|
| Bran | Bran | Liquor Lees | (YG) | ||
| Glucose | 4% | 4% | 4% | 4% | 4% |
| CSL | 0.3% | - | - | - | - |
| Rice bran | - | 0.3% | - | - | - |
| Wheat bran | - | - | 0.3% | - | - |
| Japanese liquor lees | - | - | - | 0.3% | - |
| Yeast extract | - | - | - | - | 0.3% |
| MgSO4·7H2O | 0.1% | 0.1% | 0.1% | 0.1% | 0.1% |
| KH2PO4 | 0.1% | 0.1% | 0.1% | 0.1% | 0.1% |
| CSL | CSL-1% | CSL-2% | CSL-3% | CSL-6% | Control | |
|---|---|---|---|---|---|---|
| (YG) | ||||||
| Glucose | 4% | 4% | 4% | 4% | 4% | 4% |
| CSL | 0.3% | 1% | 2% | 3% | 6% | - |
| Yeast extract | - | - | - | - | - | 0.3% |
| MgSO4·7H2O | 0.1% | 0.1% | 0.1% | 0.1% | 0.1% | 0.1% |
| KH2PO4 | 0.1% | 0.1% | 0.1% | 0.1% | 0.1% | 0.1% |
| Salts- 0.1 + 0.1% Glc-4% CSL-1% | Salts- 0.1 + 0.1% Glc-2% CSL-1% | Salts- 0.1 + 0.1% Glc-1% CSL-1% | Salts-0% Glc-4% CSL-1% | Salts-0% Glc-2% CSL1% | Salts-0% Glc-1% CSL-1% | Control (YG) | |
|---|---|---|---|---|---|---|---|
| Glucose | 4% | 2% | 1% | 4% | 2% | 1% | 4% |
| CSL | 1% | 1% | 1% | 1% | 1% | 1% | - |
| Yeast extract | - | - | - | - | - | - | 0.3% |
| MgSO4·7H2O | 0.1% | 0.1% | 0.1% | - | - | - | 0.1% |
| KH2PO4 | 0.1% | 0.1% | 0.1% | - | - | - | 0.1% |
| Salts- 0.1 + 0.1% Glc-4% CSL-6% | Salts- 0.1 + 0.1% Glc-2% CSL-6% | Salts- 0.1 + 0.1% Glc-1% CSL-6% | Salts-0% Glc-4% CSL-6% | Salts-0% Glc-2% CSL-6% | Salts-0% Glc-1% CSL-6% | Control (YG) | |
|---|---|---|---|---|---|---|---|
| Glucose | 4% | 2% | 1% | 4% | 2% | 1% | 4% |
| CSL | 6% | 6% | 6% | 6% | 6% | 6% | - |
| Yeast extract | - | - | - | - | - | - | 0.3% |
| MgSO4·7H2O | 0.1% | 0.1% | 0.1% | - | - | - | 0.1% |
| KH2PO4 | 0.1% | 0.1% | 0.1% | - | - | - | 0.1% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kobori, H.; Wu, J.; Takemura, H.; Choi, J.-H.; Tada, N.; Kawagishi, H. Utilization of Corn Steep Liquor for the Production of Fairy Chemicals by Lepista sordida Mycelia. J. Fungi 2022, 8, 1269. https://doi.org/10.3390/jof8121269
Kobori H, Wu J, Takemura H, Choi J-H, Tada N, Kawagishi H. Utilization of Corn Steep Liquor for the Production of Fairy Chemicals by Lepista sordida Mycelia. Journal of Fungi. 2022; 8(12):1269. https://doi.org/10.3390/jof8121269
Chicago/Turabian StyleKobori, Hajime, Jing Wu, Hirohide Takemura, Jae-Hoon Choi, Naoto Tada, and Hirokazu Kawagishi. 2022. "Utilization of Corn Steep Liquor for the Production of Fairy Chemicals by Lepista sordida Mycelia" Journal of Fungi 8, no. 12: 1269. https://doi.org/10.3390/jof8121269