

Effect of Flagellin Pre-Exposure on the Inflammatory and Antifungal Response of Bronchial Epithelial Cells to Fungal Pathogens
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains
2.2. Cell Culture and In Vitro Model
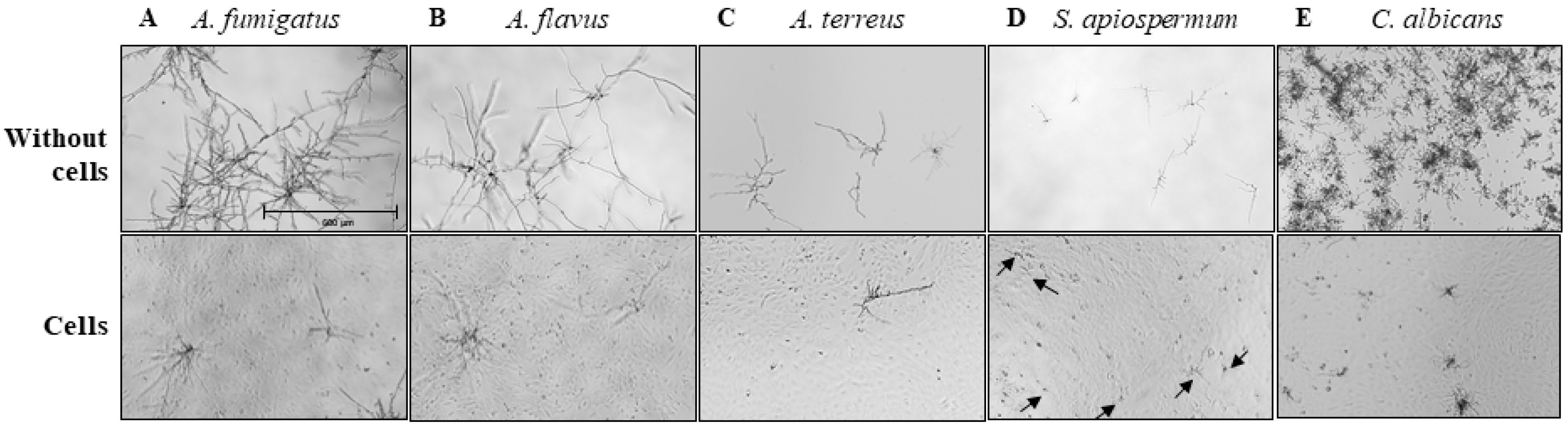
2.3. Antifungal Activity
2.4. Inflammatory Response
2.5. Cytotoxicity
2.6. Statistical Analysis
3. Results
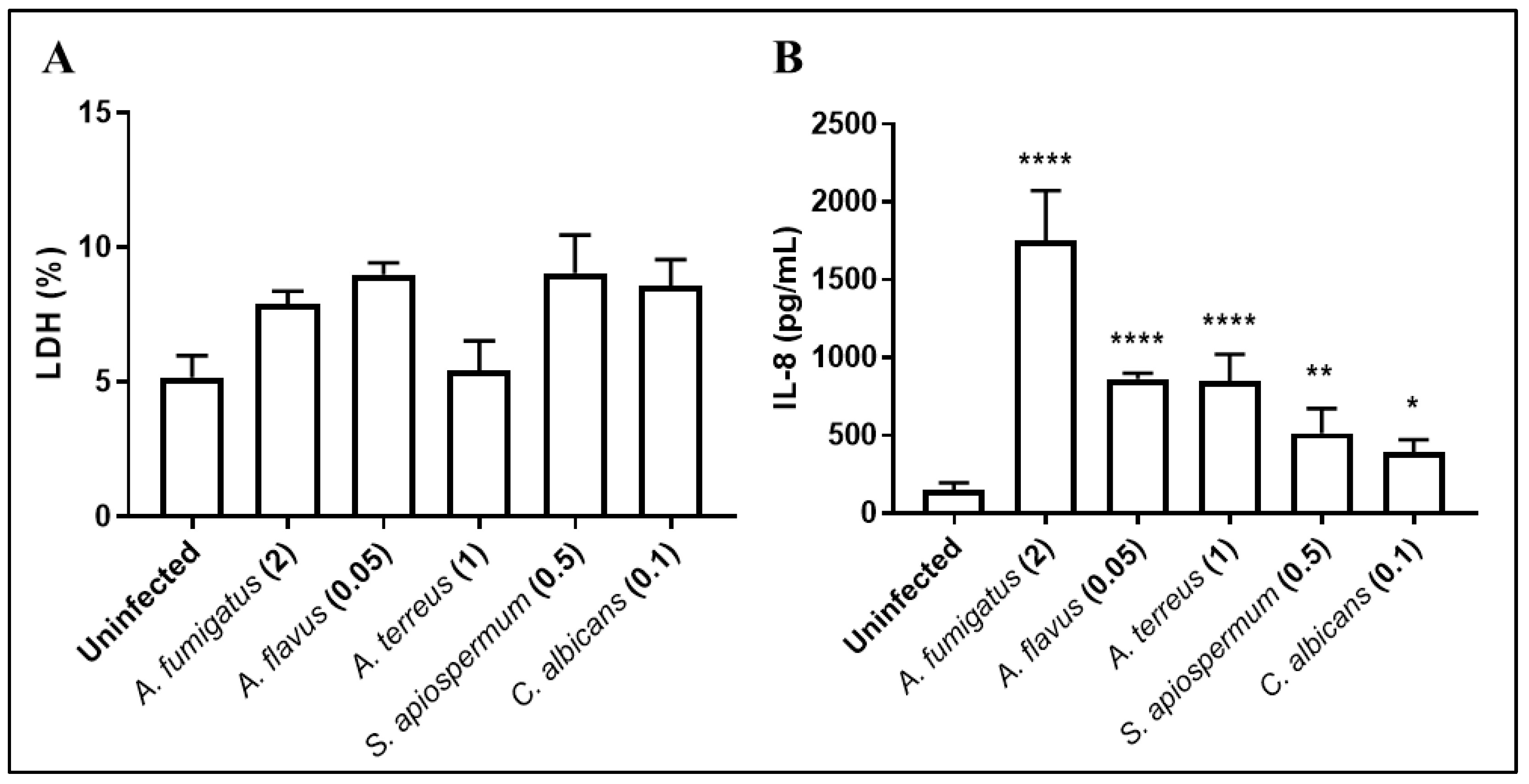
3.1. Inflammatory and Antifungal Response of BEC to Fungal Infections
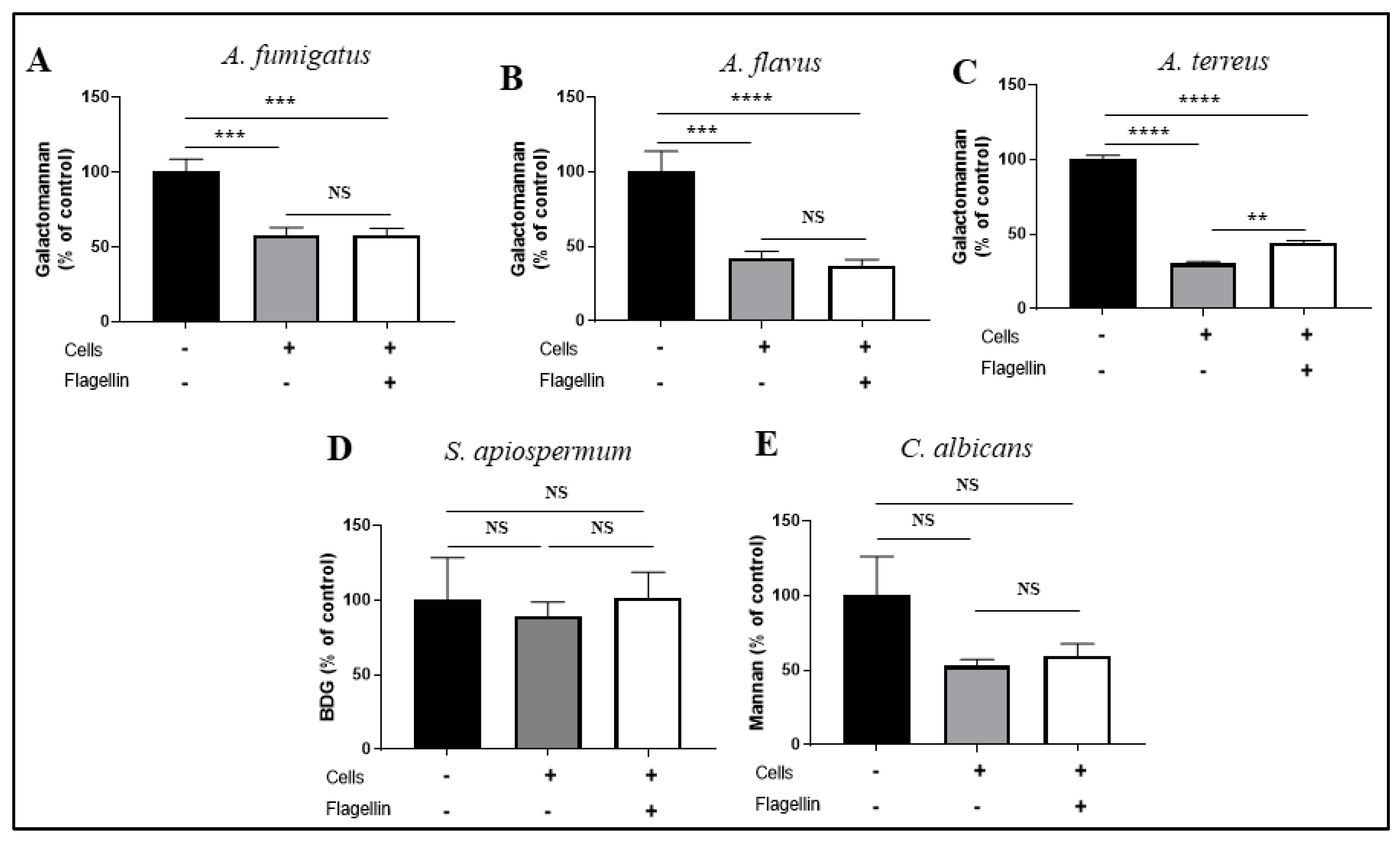
3.2. Impact of IIM on the BEC Responses toward Fungi
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ren, C.L.; Rosenfeld, M.; Mayer, O.H.; Davis, S.D.; Kloster, M.; Castile, R.G.; Hiatt, P.W.; Hart, M.; Johnson, R.; Jones, P.; et al. Analysis of the associations between lung function and clinical features in preschool children with Cystic Fibrosis. Pediatr. Pulmonol. 2012, 47, 574–581. [Google Scholar] [CrossRef] [PubMed]
- Flora, M.; Perrotta, F.; Nicolai, A.; Maffucci, R.; Pratillo, A.; Mollica, M.; Bianco, A.; Calabrese, C. Staphylococcus Aureus in chronic airway diseases: An overview. Respir. Med. 2019, 155, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Clemente, M.; de la Rosa, D.; Máiz, L.; Girón, R.; Blanco, M.; Olveira, C.; Canton, R.; Martinez-García, M.A. Impact of Pseudomonas aeruginosa Infection on Patients with Chronic Inflammatory Airway Diseases. J. Clin. Med. 2020, 9, 3800. [Google Scholar] [CrossRef] [PubMed]
- Brandt, C.; Roehmel, J.; Rickerts, V.; Melichar, V.; Niemann, N.; Schwarz, C. Aspergillus Bronchitis in Patients with Cystic Fibrosis. Mycopathologia 2018, 183, 61–69. [Google Scholar] [CrossRef]
- Tiew, P.Y.; Mac Aogáin, M.; Ter, S.K.; Aliberti, S.; Chalmers, J.D.; Chotirmall, S.H. Respiratory Mycoses in COPD and Bronchiectasis. Mycopathologia 2021, 186, 623–638. [Google Scholar] [CrossRef]
- Valenza, G.; Tappe, D.; Turnwald, D.; Frosch, M.; König, C.; Hebestreit, H.; Abele-Horn, M. Prevalence and antimicrobial susceptibility of microorganisms isolated from sputa of patients with cystic fibrosis. J. Cyst. Fibros. 2008, 7, 123–127. [Google Scholar] [CrossRef]
- Armstead, J.; Morris, J.; Denning, D.W. Multi-Country Estimate of Different Manifestations of Aspergillosis in Cystic Fibrosis. PLoS ONE 2014, 9, e98502. [Google Scholar] [CrossRef]
- Gago, S.; Denning, D.W.; Bowyer, P. Pathophysiological aspects of Aspergillus colonization in disease. Med. Mycol. 2019, 57, S219–S227. [Google Scholar] [CrossRef]
- Hatziagorou, E.; Walsh, T.J.; Tsanakas, J.N.; Roilides, E. Aspergillus and the paediatric lung. Paediatr. Respir. Rev. 2009, 10, 178–185. [Google Scholar] [CrossRef]
- Singh, A.; Ralhan, A.; Schwarz, C.; Hartl, D.; Hector, A. Fungal Pathogens in CF Airways: Leave or Treat? Mycopathologia 2018, 183, 119–137. [Google Scholar] [CrossRef]
- Engel, T.G.P.; Slabbers, L.; de Jong, C.; Melchers, W.J.G.; Hagen, F.; Verweij, P.E.; Merkus, P.; Meis, J.F. Prevalence and diversity of filamentous fungi in the airways of cystic fibrosis patients—A Dutch, multicentre study. J. Cyst. Fibros. 2019, 18, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Coron, N.; Pihet, M.; Fréalle, E.; Lemeille, Y.; Pinel, C.; Pelloux, H.; Gargala, G.; Favennec, L.; Accoceberry, I.; Durand-Joly, I.; et al. Toward the Standardization of Mycological Examination of Sputum Samples in Cystic Fibrosis: Results from a French Multicenter Prospective Study. Mycopathologia 2018, 183, 101–117. [Google Scholar] [CrossRef] [PubMed]
- Muthig, M.; Hebestreit, A.; Ziegler, U.; Seidler, M.; Müller, F.-M.C. Persistence of Candida species in the respiratory tract of cystic fibrosis patients. Med. Mycol. 2010, 48, 56–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Máiz, L.; Vendrell, M.; Olveira, C.; Girón, R.; Nieto, R.; Martínez-García, M.Á. Prevalence and Factors Associated with Isolation of Aspergillus and Candida from Sputum in Patients with Non-Cystic Fibrosis Bronchiectasis. Respiration 2015, 89, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Gileles-Hillel, A.; Shoseyov, D.; Polacheck, I.; Korem, M.; Kerem, E.; Cohen-Cymberknoh, M. Association of chronic Candida albicans respiratory infection with a more severe lung disease in patients with cystic fibrosis: Candida albicans in CF Lung Disease. Pediatric Pulmonol. 2015, 50, 1082–1089. [Google Scholar] [CrossRef]
- Baxter, C.G.; Moore, C.B.; Jones, A.M.; Webb, A.K.; Denning, D.W. IgE-Mediated Immune Responses and Airway Detection of Aspergillus and Candida in Adult Cystic Fibrosis. Chest 2013, 143, 1351–1357. [Google Scholar] [CrossRef]
- Roach, J.C.; Glusman, G.; Rowen, L.; Kaur, A.; Purcell, M.K.; Smith, K.D.; Hood, L.E.; Aderem, A. The evolution of vertebrate Toll-like receptors. Proc. Natl. Acad. Sci. USA 2005, 102, 9577–9582. [Google Scholar] [CrossRef] [Green Version]
- Bigot, J.; Guillot, L.; Guitard, J.; Ruffin, M.; Corvol, H.; Balloy, V.; Hennequin, C. Bronchial Epithelial Cells on the Front Line to Fight Lung Infection-Causing Aspergillus fumigatus. Front. Immunol. 2020, 11, 1041. [Google Scholar] [CrossRef]
- Kang, H.-K.; Kim, C.; Seo, C.H.; Park, Y. The therapeutic applications of antimicrobial peptides (AMPs): A patent review. J. Microbiol. 2017, 55, 1–12. [Google Scholar] [CrossRef]
- Richard, N.; Marti, L.; Varrot, A.; Guillot, L.; Guitard, J.; Hennequin, C.; Imberty, A.; Corvol, H.; Chignard, M.; Balloy, V. Human Bronchial Epithelial Cells Inhibit Aspergillus fumigatus Germination of Extracellular Conidia via FleA Recognition. Sci. Rep. 2018, 8, 15699. [Google Scholar] [CrossRef]
- Netea, M.G.; Quintin, J.; van der Meer, J.W.M. Trained Immunity: A Memory for Innate Host Defense. Cell Host Microbe 2011, 9, 355–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ifrim, D.C.; Quintin, J.; Joosten, L.A.B.; Jacobs, C.; Jansen, T.; Jacobs, L.; Gow, N.A.R.; Williams, D.L.; van der Meer, J.W.M.; Netea, M.G. Trained Immunity or Tolerance: Opposing Functional Programs Induced in Human Monocytes after Engagement of Various Pattern Recognition Receptors. Clin. Vaccine Immunol. 2014, 21, 534–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Meer, J.W.M.; Joosten, L.A.B.; Riksen, N.; Riksen, N.; Netea, M.G. Trained immunity: A smart way to enhance innate immune defence. Mol. Immunol. 2015, 68, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.A.; Elliott, J.M.; Keyel, P.A.; Yang, L.; Carrero, J.A.; Yokoyama, W.M. Cytokine-induced memory-like natural killer cells. Proc. Natl. Acad. Sci. USA 2009, 106, 1915–1919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaufmann, E.; Sanz, J.; Dunn, J.L.; Khan, N.; Mendonça, L.E.; Pacis, A.; Tzelepis, F.; Pernet, E.; Dumaine, A.; Grenier, J.-C.; et al. BCG Educates Hematopoietic Stem Cells to Generate Protective Innate Immunity against Tuberculosis. Cell 2018, 172, 176–190.e19. [Google Scholar] [CrossRef] [Green Version]
- Naik, S.; Larsen, S.B.; Gomez, N.C.; Alaverdyan, K.; Sendoel, A.; Yuan, S.; Polak, L.; Kulukian, A.; Chai, S.; Fuchs, E. Inflammatory Memory Sensitizes Skin Epithelial Stem Cells to Tissue Damage. Nature 2017, 550, 475–480. [Google Scholar] [CrossRef] [Green Version]
- Bigot, J.; Guillot, L.; Guitard, J.; Ruffin, M.; Corvol, H.; Chignard, M.; Hennequin, C.; Balloy, V. Respiratory Epithelial Cells Can Remember Infection: A Proof of Concept Study. J. Infect. Dis. 2019, 221, 1000–1005. [Google Scholar] [CrossRef]
- Reihill, J.A.; Moore, J.E.; Elborn, J.S.; Ennis, M. Effect of Aspergillus fumigatus and Candida albicans on pro-inflammatory response in cystic fibrosis epithelium. J. Cyst. Fibros. 2011, 10, 401–406. [Google Scholar] [CrossRef] [Green Version]
- Odabasi, Z.; Paetznick, V.L.; Rodriguez, J.R.; Chen, E.; McGinnis, M.R.; Ostrosky-Zeichner, L. Differences in beta-glucan levels in culture supernatants of a variety of fungi. Med. Mycol. 2006, 44, 267–272. [Google Scholar] [CrossRef] [Green Version]
- Turcios, N.L. Cystic Fibrosis Lung Disease: An Overview. Respir. Care 2020, 65, 233–251. [Google Scholar] [CrossRef]
- Lin, V.Y.; Kaza, N.; Birket, S.E.; Kim, H.; Edwards, L.J.; LaFontaine, J.; Liu, L.; Mazur, M.; Byzek, S.A.; Hanes, J.; et al. Excess mucus viscosity and airway dehydration impact COPD airway clearance. Eur. Respir. J. 2020, 55, 1900419. [Google Scholar] [CrossRef] [PubMed]
- Balloy, V.; Chignard, M. The innate immune response to Aspergillus fumigatus. Microbes Infect. 2009, 11, 919–927. [Google Scholar] [CrossRef] [PubMed]
- Xisto, M.I.D.S.; Liporagi-Lopes, L.C.; Muñoz, J.E.; Bittencourt, V.C.B.; Santos, G.M.P.; Dias, L.S.; Figueiredo, R.T.; Pinto, M.R.; Taborda, C.P.; Barreto-Bergter, E. Peptidorhamnomannan negatively modulates the immune response in a scedosporiosis murine model. Med. Mycol. 2016, 54, 846–855. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Alarcón, D.; Balloy, V.; Bouchara, J.-P.; Pieters, R.J.; Varrot, A. Biochemical and structural studies of target lectin SapL1 from the emerging opportunistic microfungus Scedosporium apiospermum. Sci. Rep. 2021, 11, 16109. [Google Scholar] [CrossRef]
- Tiew, P.Y.; Jaggi, T.K.; Chan, L.L.Y.; Chotirmall, S.H. The airway microbiome in COPD, bronchiectasis and bronchiectasis-COPD overlap. Clin. Respir. J. 2021, 15, 123–133. [Google Scholar] [CrossRef]
- Zhao, J.; Cheng, W.; He, X.; Liu, Y. The co-colonization prevalence of Pseudomonas aeruginosa and Aspergillus fumigatus in cystic fibrosis: A systematic review and meta-analysis. Microb. Pathog. 2018, 125, 122–128. [Google Scholar] [CrossRef]
- Gao, N.; Kumar, A.; Guo, H.; Wu, X.; Wheater, M.; Yu, F.-S.X. Topical Flagellin-Mediated Innate Defense against Candida albicans Keratitis. Investig. Ophthalmol. Vis. Sci. 2011, 52, 3074–3082. [Google Scholar] [CrossRef] [Green Version]
- Sohrabi, Y.; Reinecke, H.; Joosten, L.A.B.; Netea, M.G. Deadly COVID-19 among the elderly: Innate immune memory helping those most in need. Med 2021, 2, 378–383. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A. fumigatus | A. flavus | A. terreus | S. apiospermum | C. albicans | |
|---|---|---|---|---|---|
| Cytotoxicity | → | → | → | → | → |
| Antifungal Activity | → | → | ↓ | → | → |
| IL-8 Synthesis | ↑ | ↑ | → | → | ↑ |
| IL-6 Synthesis | ↑ | → | → | ↑ | → |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bigot, J.; Ruffin, M.; Guitard, J.; Vellaissamy, S.; Thorez, S.; Corvol, H.; Guillot, L.; Balloy, V.; Hennequin, C. Effect of Flagellin Pre-Exposure on the Inflammatory and Antifungal Response of Bronchial Epithelial Cells to Fungal Pathogens. J. Fungi 2022, 8, 1268. https://doi.org/10.3390/jof8121268
Bigot J, Ruffin M, Guitard J, Vellaissamy S, Thorez S, Corvol H, Guillot L, Balloy V, Hennequin C. Effect of Flagellin Pre-Exposure on the Inflammatory and Antifungal Response of Bronchial Epithelial Cells to Fungal Pathogens. Journal of Fungi. 2022; 8(12):1268. https://doi.org/10.3390/jof8121268
Chicago/Turabian StyleBigot, Jeanne, Manon Ruffin, Juliette Guitard, Sandra Vellaissamy, Sophie Thorez, Harriet Corvol, Loïc Guillot, Viviane Balloy, and Christophe Hennequin. 2022. "Effect of Flagellin Pre-Exposure on the Inflammatory and Antifungal Response of Bronchial Epithelial Cells to Fungal Pathogens" Journal of Fungi 8, no. 12: 1268. https://doi.org/10.3390/jof8121268