

Four New Species and New Records of Orbilia from China Based on Molecular and Morphological Data

Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphological Studies
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Phylogenetic Analysis
3. Results
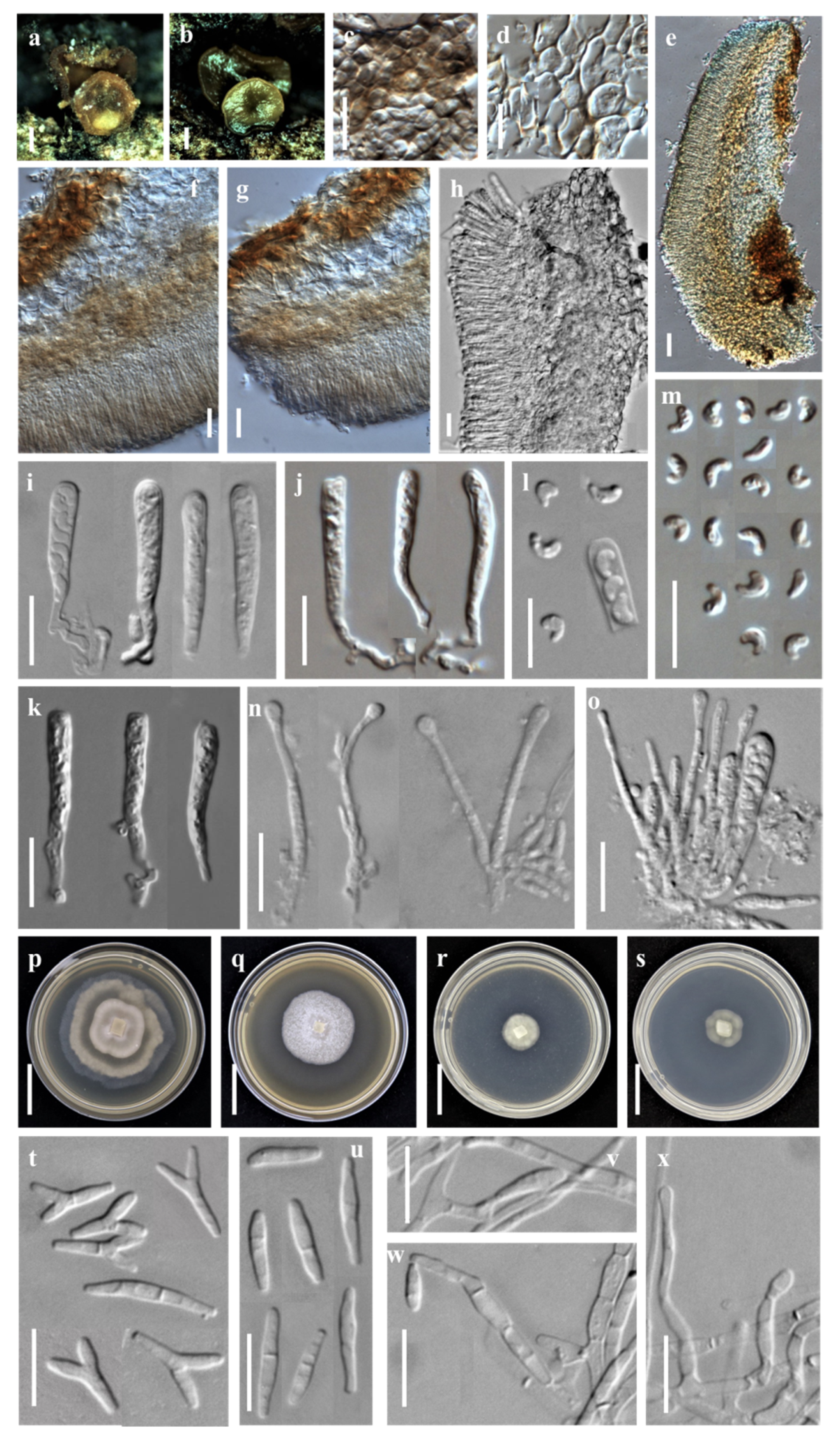
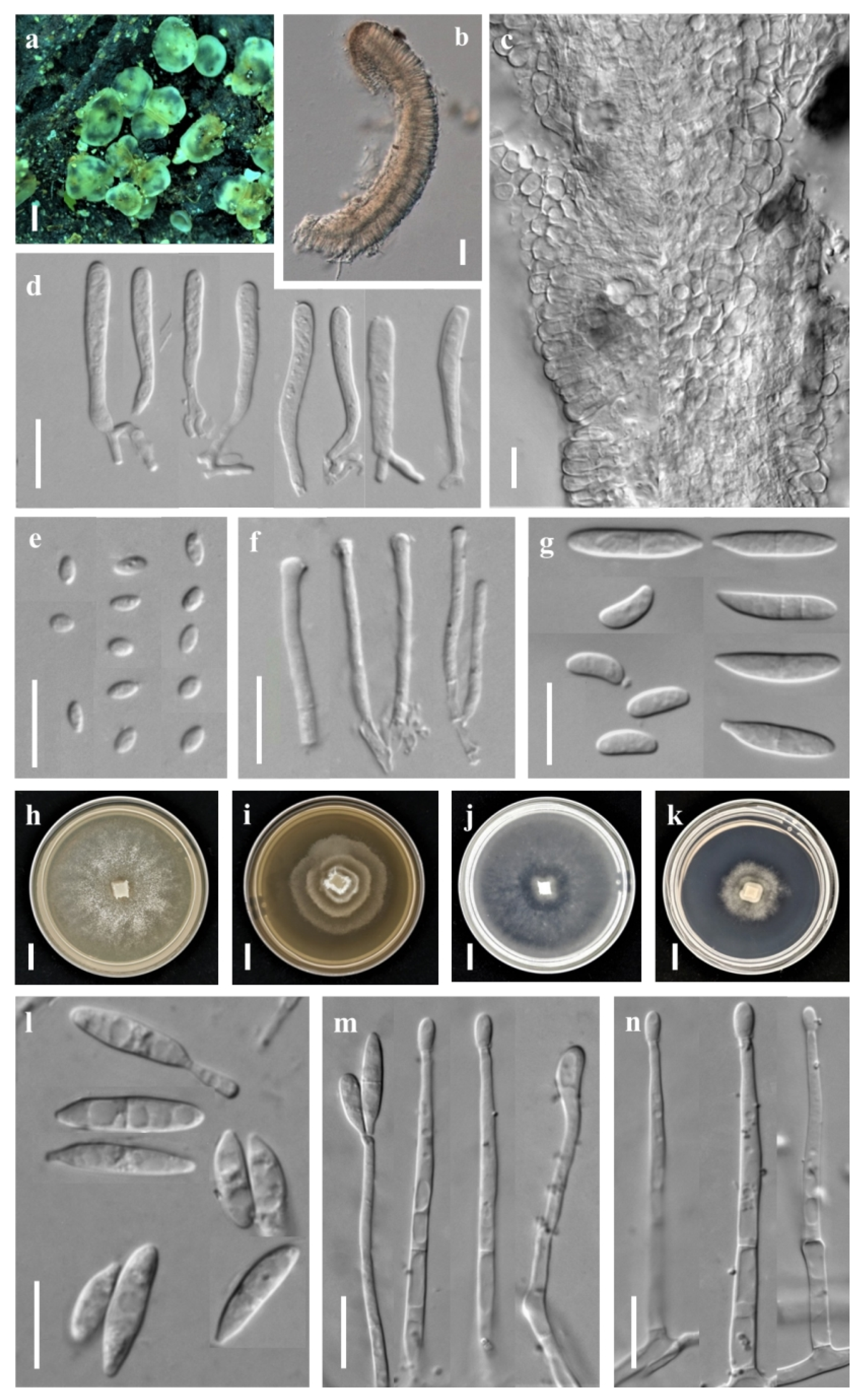
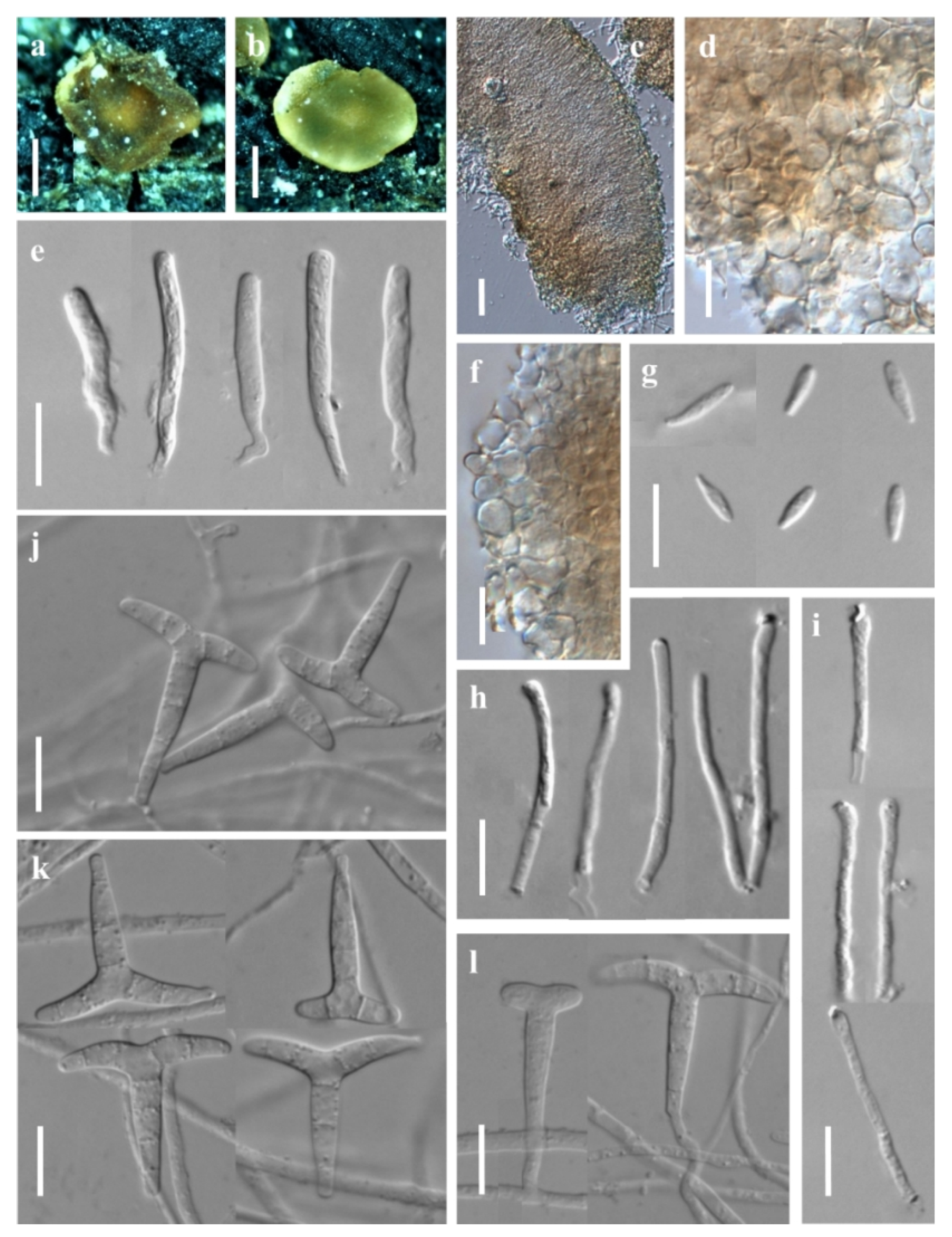
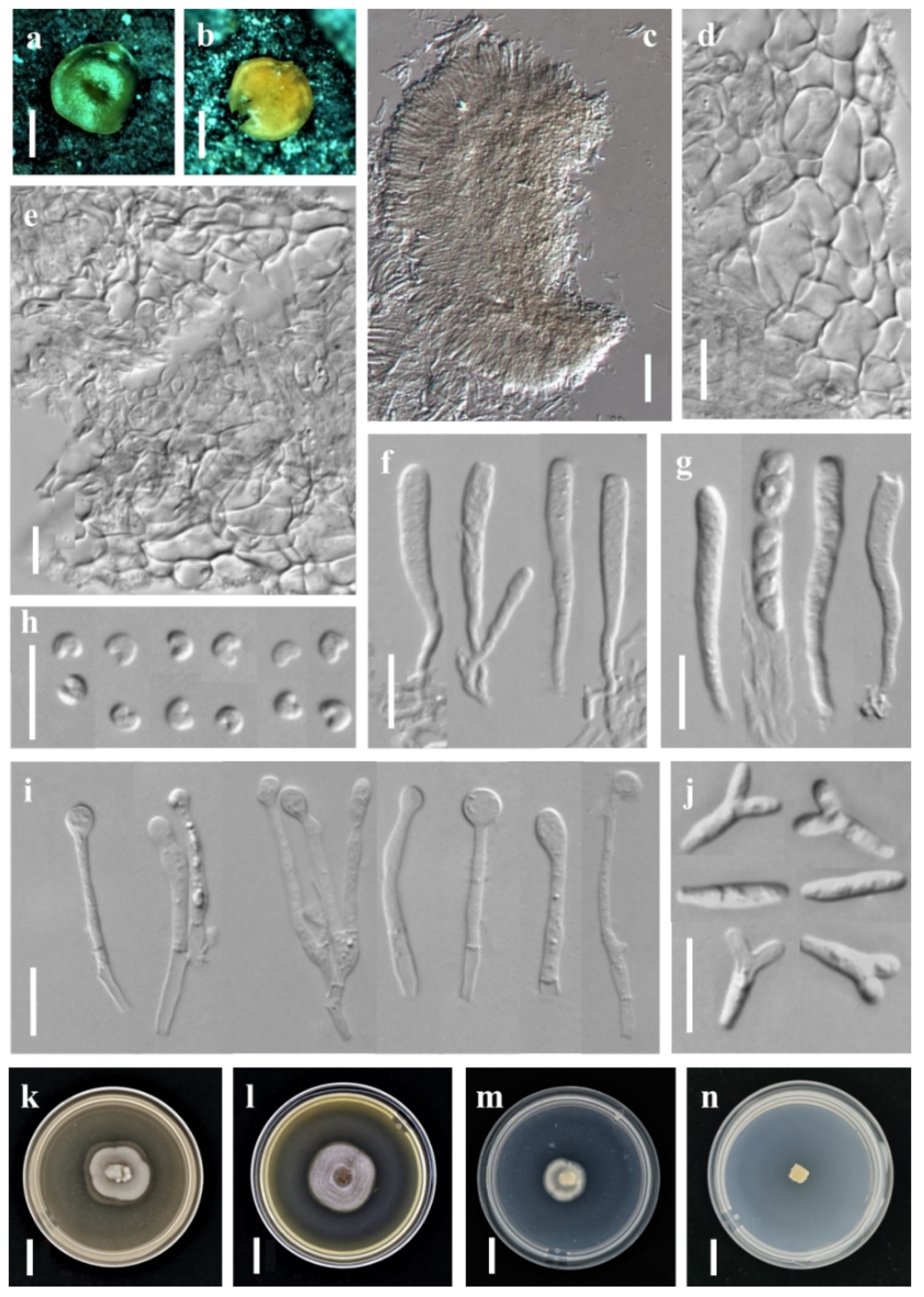
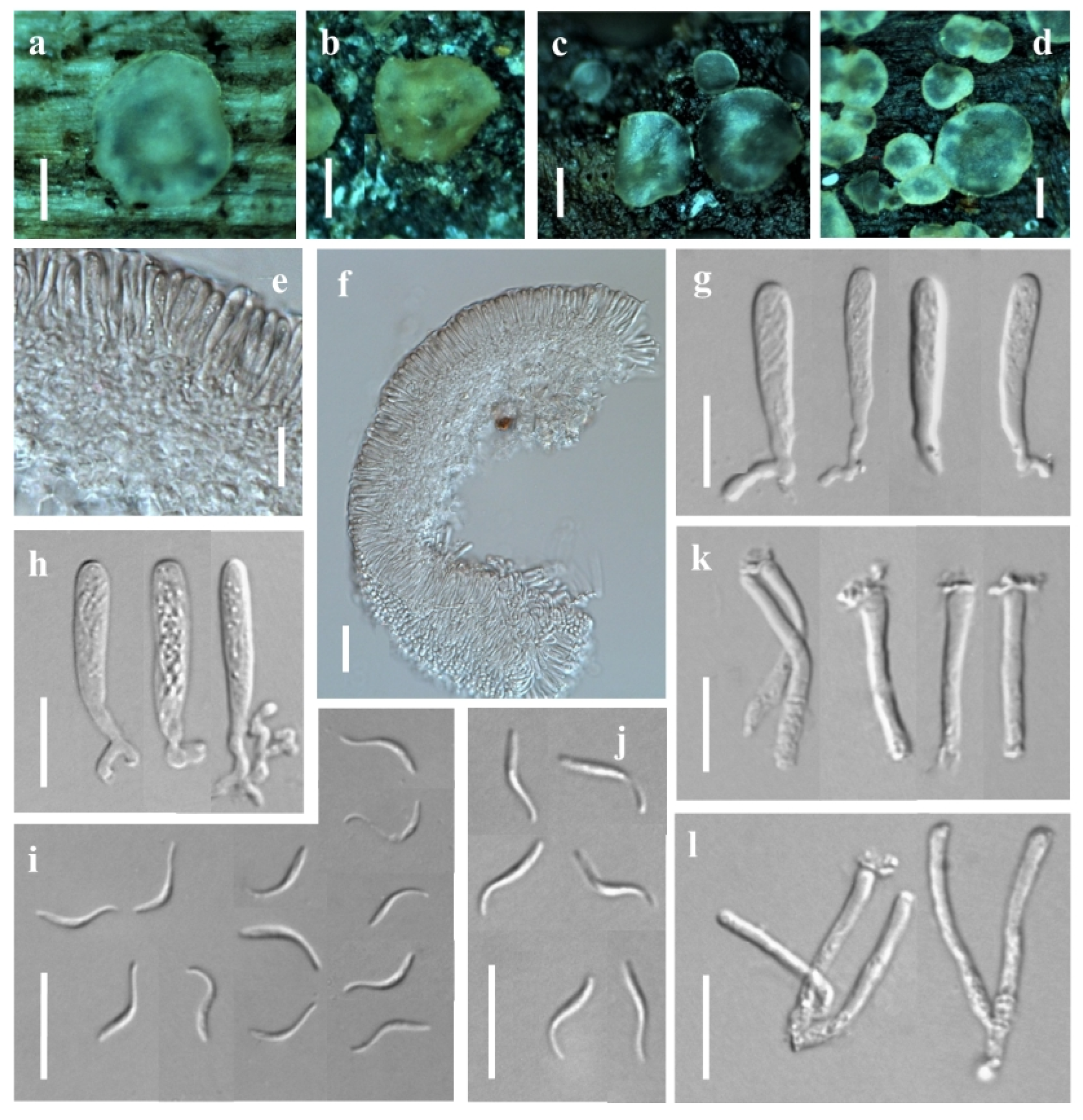
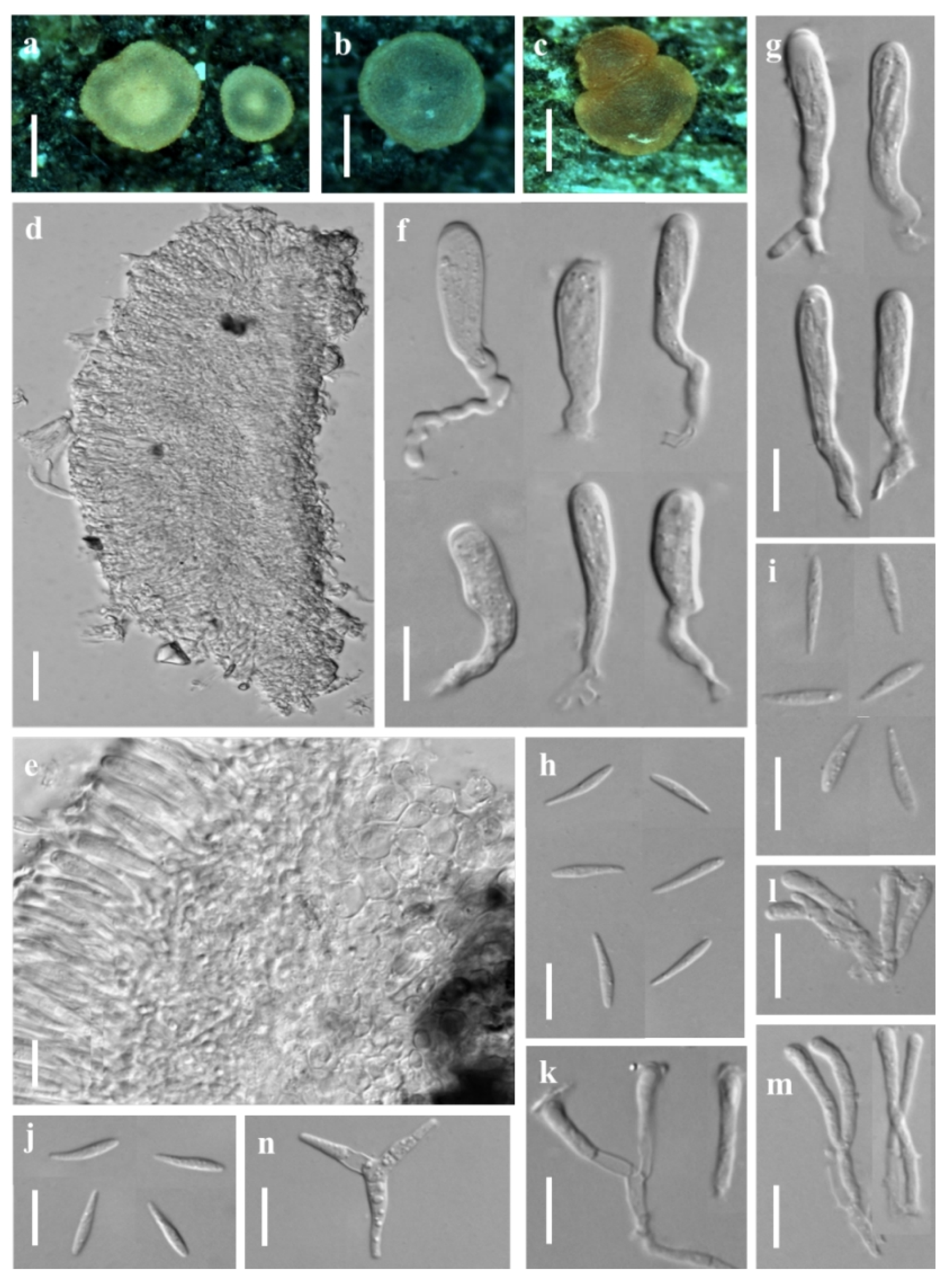
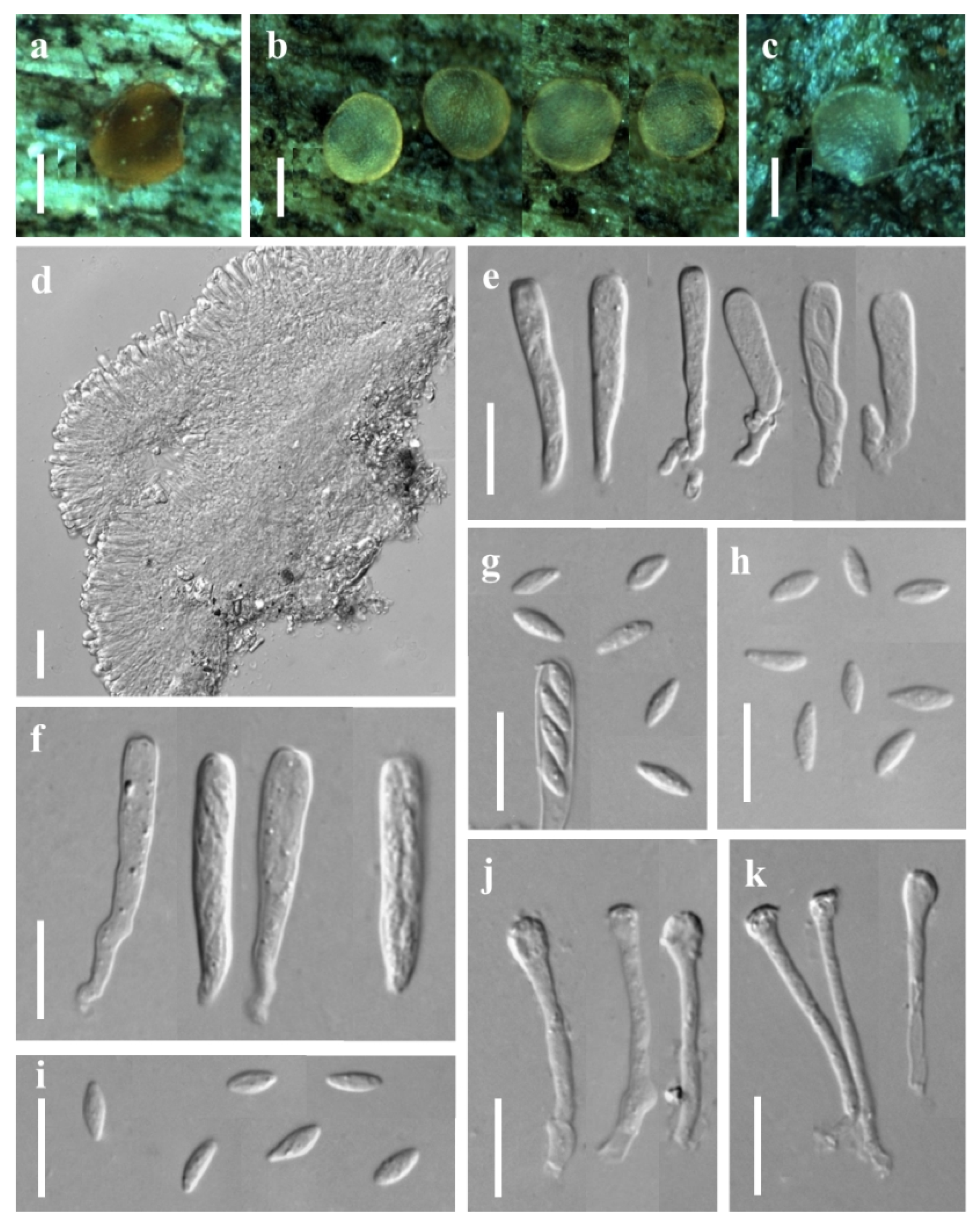
3.1. Taxonomic Description
3.1.1. New Species
3.1.2. New Record Species
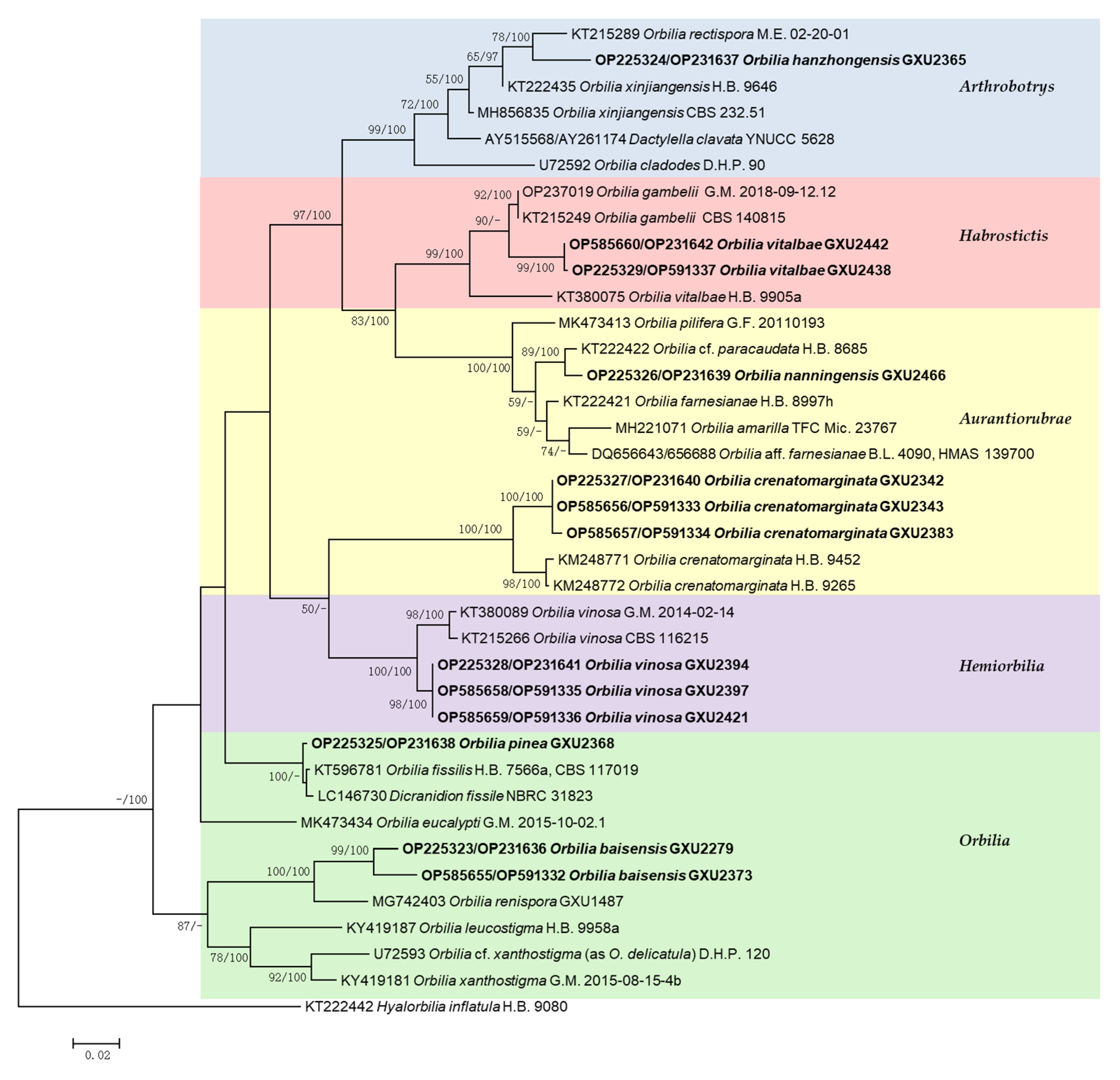
3.2. Phylogenetic Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ekanayaka, A.H.; Hyde, K.D.; Jones, E.B.G.; Zhao, Q. Orbiliaceae from Thailand. Mycosphere 2018, 9, 155–168. [Google Scholar] [CrossRef]
- Marchal, E. Champignons coprophiles de Belgique. Bull. Soc. R. Bot. Belg. 1885, 24, 56–77. [Google Scholar]
- Baral, H.O.; Weber, E.; Marson, G. Monograph of Orbiliomycetes (Ascomycota) Based on Vital Taxonomy; National Museum of Natural History: Luxembourg, 2020. [Google Scholar]
- Baral, H.O.; Weber, E.; Gams, W.; Hagedorn, G.; Liu, B.; Liu, X.Z.; Marson, G.; Marvanová, L.; Stadler, M.; Weiβ, M. Generic names in the Orbiliaceae (Orbiliomycetes) and recommendations on which names should be protected or suppressed. Mycol. Prog. 2018, 17, 5–31. [Google Scholar] [CrossRef] [Green Version]
- Fires, E.M. Corpus florarum Provincialium Sueciae. I. Floram scanicum. XXIV; Sebell & C: Upasla, Sweden, 1835; 394p. [Google Scholar]
- Nannfeldt, J.Z. Studien über die Morphologie und Systematik der nich-lichenisierten inoperculaten Discomyceten. Nova Acta Regiae Soc. Sci. Ups. Ser. 1932, 48, 368. [Google Scholar]
- Spooner, M.P. Helotiales of Australasia: Geoglossaceae, Orbiliaceae, Sclerotiniaceae, Hyaloscyphaceae. Biblioth Mycol. 1987, 116, 1–711. [Google Scholar]
- Eriksson, O.E.; Baral, H.O.; Currah, R.S.; Hansen, K.; Kurtzman, C.P.; Læssøe, T.; Rambold, G. Notes on ascomycete systematics Nos 3580–3623. Myconet 2003, 9, 91–103. [Google Scholar]
- Zhang, Y.; Yu, Z.F.; Baral, H.O.; Qiao, M.; Zhang, K.Q. Pseudorbilia gen. nov. (Orbiliaceae) from Yunnan, China. Fungal Divers. 2007, 26, 305–312. [Google Scholar]
- Kumar, T.A.; Healy, R.; Spatafora, J.W.; Blackwell, M.; Mclanughlin, D.J. Orbilia ultrastructure, character evolution and phylogeny of Pezizomycotina. Mycologia 2012, 104, 462–476. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.W.; Li, S.F.; Yang, L.F.; Yang, J.; Ye, T.Z.; Yang, L. New records and new distribution of known species in the family Orbiliaceae from China. Afr. J. Microbiol. Res. 2014, 8, 3178–3190. [Google Scholar]
- Qiao, M.; Li, J.Y.; Baral, H.O.; Zhang, Y.; Qiao, W.Y.; Su, H.Y.; Yu, Z.F. Orbilia yuanensis sp. nov. and its anamorph. Mycol. Prog. 2015, 14, 1. [Google Scholar] [CrossRef]
- Quijada, L.; Baral, H.O.; Beltrán-Tejera, E. New species of Orbilia (Orbiliales) from arid ecosystems of the Canary Islands (Spain). Nova Hedwig. 2012, 96, 237–248. [Google Scholar] [CrossRef]
- Quijada, L.; Baral, H.O.; Jaen-Molina, R.; Weiss, M.; Caujapé-Castelis, J.; Beltrán-Tejera, E. Phylogenetic and morphological circumscription of the Orbilia aurantiorubra group. Phytotaxa 2014, 175, 30. [Google Scholar] [CrossRef]
- Quijada, L.; Baral, H.O.; Beltrán-Tejera, E. Diversity of Hyalorbilia (Orbiliales) in the Macaronesian Region. Nova Hedwig. 2015, 100, 1–14. [Google Scholar] [CrossRef]
- Quijada, L.; Baral, H.O.; Beltrán-Tejera, E. A revision of the genus Orbilia in the Canary Islands. Phytotaxa 2016, 284, 231–262. [Google Scholar] [CrossRef]
- Quijada, L.; Baral, H.O. Orbilia beltraniae, a new succulenticolous species from the Canary Islands. MycoKeys 2017, 25, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Shao, Y.Y.; Baral, H.O.; Ou, X.Y.; Wu, H.; Huang, F.C.; Zheng, H.F.; Liu, B. New species and records of orbiliaceous fungi from Georgia, USB. Mycol. Prog. 2018, 17, 1225–1235. [Google Scholar] [CrossRef]
- Deng, C.; Yu, Z.F. The complete mitochondrial genomes of Dactylellina leptospora (Orbiliales, Orbiliaceae). Mitochondrial DNA Part B Resour. 2019, 4, 1615–1616. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.J.; Fang, M.L.; Xu, J.P.; Jiang, L.L.; Zhou, D.Y.; Zhang, K.Q.; Zhang, Y. Complete mitochondrial genome and phylogenetic analysis of Orbilia dorsalia, a species producing mature sexual structures on culture. Mitochondrial DNA B 2019, 4, 573–574. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4239–4246. [Google Scholar] [CrossRef] [Green Version]
- Baral, H.O.; Johnston, P.; Quijada, L.; Healy, R.; Pfister, D.H.; Lobuglio, K.F.; Rodriguez, V.; Weber, E. Cryptic speciation in Orbilia xanthostigma and O. leucostigma (Orbiliomycetes): An aggregate with worldwide distribution. Mycol. Prog. 2021, 20, 1503–1537. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Woudenberg, J.H.C.; Aveskamp, M.M.; De-Gruyter, J.; Spiers, A.G.; Crous, P.W. Multiple Didymella teleomorphs are linked to the Phoma clematidina morphotype. Persoonia 2009, 22, 56–62. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, G.H.; Sung, J.M.; Hywel-Jones, N.L.; Spatafora, J.W. A multigene phylogeny of Clavicipitaceae (Ascomycota, fungi): Identification of localized incongruence using a combinational bootstrap approach. Mol. Phylogenet. Evol. 2007, 44, 1204–1223. [Google Scholar] [CrossRef] [PubMed]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Brefeld, O. AmericaUntersuchungen aus dem Gesammtgebiete der Mykologie, Ascomyceten II. Mycology 1891, 7, 50–51. [Google Scholar]
- Pfister, D.H. Castor, Pollux, and life histories of fungi. Micologia 1997, 89, 1–23. [Google Scholar] [CrossRef]
- Harkness, H.W. Fungi of the Pacific Coast. Bull. Calif. Acad. Sci. 1885, 1, 159–176. [Google Scholar]
- Hughes, S.J. Conidiophores, conidia, and classification. Can. J. Bot. 1953, 31, 577–659. [Google Scholar] [CrossRef]
- Peek, C.A.; Solheim, W.G. The hyphomycetous genera of H. W. Harkness and the ascomycetous genus Cleistosama Harkn. Mycologia 1958, 50, 844–861. [Google Scholar] [CrossRef]
- Berthet, P. Forrnes conidiennes de divers Discomycetes. Bull. Soc. Mycol. Fr. 1964, 80, 125–149. [Google Scholar]
- Liu, B.; Liu, X.Z.; Zhuang, W.Y. Orbilia querci sp. nov. and its knob-forming nematophagous anamorph. FEMS Microbiol. Lett. 2005, 245, 99–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, J.J. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Vitória, N.S.; Bezerra, J.L.; Gramacho, K.P. A simplified DNA Extraction Method for PCR Analysis of Camarotella spp. Braz. Arch. Biol. Technol. 2010, 55, 249–252. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The Clustal_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Gao, F.L.; Jakovlic, I.; Zhou, H.; Zhang, J.; Li, W.X.; Wang, G.T. Phylosuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP 4.0b10: Phylogenetic Analysis Using Parsimony (* and Other Methods); Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree v1.4.3 2016. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 1 August 2022).
- Liu, B.; Liu, X.Z.; Zhuang, W.Y.; Baral, H.O. Orbiliaceous fungi from Tibet, China. Fungal Divers. 2006, 22, 107–120. [Google Scholar]
- Priou, J.P.; Poncelet, A. Orbilia crystallina et Tapesina griseovitellina, deux rare ascomycètes nouveaux pour la Bretagne. Cah. Mycol. Nantais 2006, 18, 3–8. [Google Scholar]
- Albertini, I.B.; Schweinitz, L.D. Conspectus Fungorum in Lusatiae Superioris Agro Niskiensi Crescentium e Methodo Persooniana; Sumtibus Kummerianis: Lipsiae, Germany, 1805; 376p. [Google Scholar]
- Ade, A. Mykologische Beiträge. Hedwigia 1923, 64, 286–320. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain Number | GenBank Accession Number | |
|---|---|---|---|
| ITS | LSU | ||
| Dactylella clavata | YNUCC 5628 | AY515568 | AY261174 |
| Dicranidion fissile | NBRC 31823 | LC146730 | LC146730 |
| Hyalorbilia inflatula | H.B. 9080 | KT222442 | KT222442 |
| Orbilia amarilla | TFC Mic. 23767 | MH221071 | MH221071 |
| Orbilia baisensis * | DL17 (GXU2279) | OP225323 | OP231636 |
| Orbilia baisensis * | BY44 (GXU2373) | OP585655 | OP591332 |
| Orbilia cladodes | D.H.P. 90 | U72592 | U72592 |
| Orbilia crenatomarginata ** | (GXU2342) | OP225327 | OP231640 |
| Orbilia crenatomarginata ** | (GXU2343) | OP585656 | OP591333 |
| Orbilia crenatomarginata ** | (GXU2383) | OP585657 | OP591334 |
| Orbilia crenatomarginata | H.B. 9452 | KM248771 | KM248771 |
| Orbilia crenatomarginata | H.B. 9265 | KM248772 | KM248772 |
| Orbilia eucalypti | G.M. 2015-10-02.1 | MK473434 | MK473434 |
| Orbilia farnesianae | H.B. 8997h | KT222421 | KT222421 |
| Orbilia aff. farnesianae | B.L. 4090 (HMAS 139700) | DQ656643 | DQ656688 |
| Orbilia fissilis | CBS 117019 | KT596781 | KT596781 |
| Orbilia gambelii | CBS 140815 | KT215249 | KT215249 |
| Orbilia gambelii | G.M. 2018-09-12.12 | OP237019 | OP237019 |
| Orbilia hanzhongensis * | BY35 (GXU2365) | OP225324 | OP231637 |
| Orbilia leucostigma | H.B. 9958a | KY419187 | KY419187 |
| Orbilia nanningensis * | NN01 (GXU2466) | OP225326 | OP231639 |
| Orbilia cf. paracaudata | H.B. 8685 | KT222422 | KT222422 |
| Orbilia pilifera | G.F. 20110193 | MK473413 | MK473413 |
| Orbilia pinea * | BY38 (GXU2368) | OP225325 | OP231638 |
| Orbilia rectispora | M.E. 02-20-01 | KT215289 | KT215289 |
| Orbilia renispora | GXU1487 | MG742403 | MG742404 |
| Orbilia vinosa ** | (GXU2394) | OP225328 | OP231641 |
| Orbilia vinosa ** | (GXU2397) | OP585658 | OP591335 |
| Orbilia vinosa ** | (GXU2421) | OP585659 | OP591336 |
| Orbilia vinosa | G.M. 2014-02-14 | KT380089 | KT380089 |
| Orbilia vinosa | CBS 116215 | KT215266 | KT215266 |
| Orbilia vitalbae ** | (GXU2438) | OP225329 | OP231642 |
| Orbilia vitalbae ** | (GXU2442) | OP585660 | OP591337 |
| Orbilia vitalbae | H.B. 9905a | KT380075 | KT380075 |
| Orbilia xanthostigma | G.M. 2015-08-15-4b | KY419181 | KY419181 |
| Orbilia cf. xanthostigma | D.H.P. 120 | U72593 | U72593 |
| Orbilia xinjiangensis | CBS 232.51 | MH856835 | MH856835 |
| Orbilia xinjiangensis | H.B. 9646 | KT222435 | KT222435 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ou, X.-Y.; Shao, Y.-Y.; Zheng, H.-F.; Liu, B. Four New Species and New Records of Orbilia from China Based on Molecular and Morphological Data. J. Fungi 2022, 8, 1188. https://doi.org/10.3390/jof8111188
Ou X-Y, Shao Y-Y, Zheng H-F, Liu B. Four New Species and New Records of Orbilia from China Based on Molecular and Morphological Data. Journal of Fungi. 2022; 8(11):1188. https://doi.org/10.3390/jof8111188
Chicago/Turabian StyleOu, Xiao-Yun, Yuan-Yuan Shao, Hai-Fu Zheng, and Bin Liu. 2022. "Four New Species and New Records of Orbilia from China Based on Molecular and Morphological Data" Journal of Fungi 8, no. 11: 1188. https://doi.org/10.3390/jof8111188