
Enhancement of Fungal Enzyme Production by Radio-Frequency Electromagnetic Fields

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strain and Growth Conditions
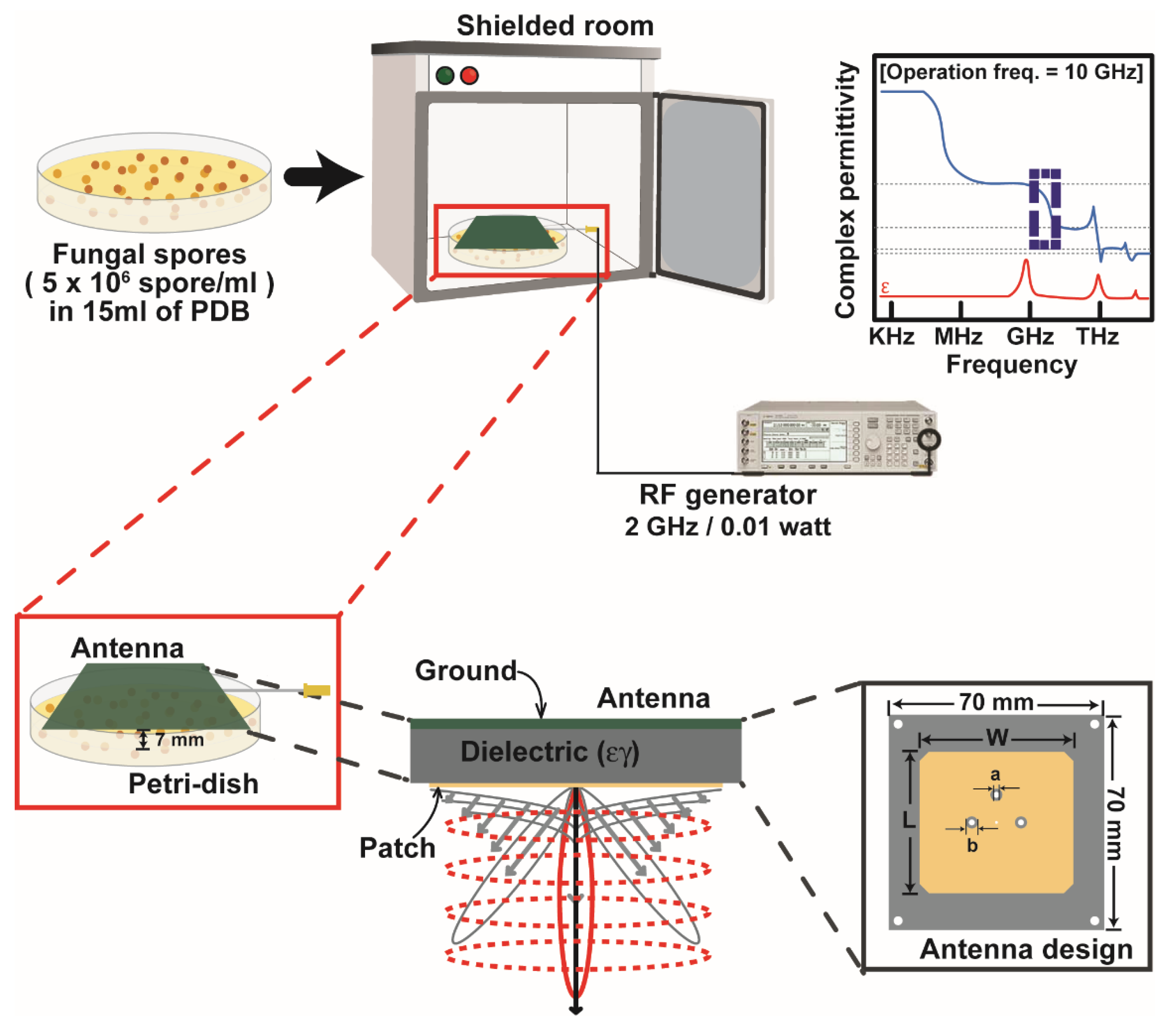
2.2. Exposure of Fungal Spores to Radio-Frequency Electromagnetic Fields (RF-EMF)
2.3. Measurement of the Total Protein Level and α-Amylase Activity in the Media
2.4. Analysis of Extracellular α-Amylase Levels Using Aptamers
2.5. Membrane Potential Assay and Vesicle Staining
2.6. Detection of Intracellular Calcium Ions
2.7. Quantitative PCR Analysis
2.8. Measurement of pH and Levels of Hydrogen Peroxide (H2O2), Nitrogen Oxides (NOx) and Inositol 1,4,5-Triphosphates (IP3)
2.9. Statistical Analysis
3. Results
3.1. Production of α-Amylase was Enhanced in the Fungus after Exposure to RF-EMF
3.2. RF-EMF Enhanced Vesicle Accumulation and the Transcription of Genes Involved in Cellular Trafficking
3.3. RF-EMF Exposure Enhanced the Level of Intracellular Ca2+ in Fungal Hyphae
3.4. Analysis for the Level of H2O2, NOx, and pH
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, R.; Kumar, M.; Mittal, A.; Mehta, P.K. Microbial enzymes: Industrial progress in 21st century. 3 Biotech 2016, 6, 174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurung, N.; Ray, S.; Bose, S.; Rai, V. A broader view: Microbial enzymes and their relevance in industries, medicine, and beyond. BioMed Res. Int. 2013, 2013, 329121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raveendran, S.; Parameswaran, B.; Ummalyma, S.B.; Abraham, A.; Mathew, A.K.; Madhavan, A.; Rebello, S.; Pandey, A. Applications of microbial enzymes in food industry. Food Technol. Biotechnol. 2018, 56, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Overmann, J.; Scholz, A.H. Microbiological research under the Nagoya protocol: Facts and fiction. Trends Microbiol. 2017, 25, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, K.; Nagashima, T.; Yamamoto, Y.; Gomi, K.; Kitamoto, K.; Kumagai, C.; Tamura, G. High level secretion of calf chymosin using a glucoamylase-prochymosin fusion gene in Aspergillus oryzae. Biosci. Biotechnol. Biochem. 1994, 58, 895–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakajima, K.-I.; Asakura, T.; Maruyama, J.-I.; Morita, Y.; Oike, H.; Shimizu-Ibuka, A.; Misaka, T.; Sorimachi, H.; Arai, S.; Kitamoto, K.; et al. Extracellular production of neoculin, a sweet-tasting heterodimeric protein with taste-modifying activity, by Aspergillus oryzae. Appl. Environ. Microbiol. 2006, 72, 3716–3723. [Google Scholar] [CrossRef] [Green Version]
- Ito, K.; Asakura, T.; Morita, Y.; Nakajima, K.-I.; Koizumi, A.; Shimizu-Ibuka, A.; Masuda, K.; Ishiguro, M.; Terada, T.; Maruyama, J.-I.; et al. Microbial production of sensory-active miraculin. Biochem. Biophys. Res. Commun. 2007, 360, 407–411. [Google Scholar] [CrossRef]
- Jin, F.J.; Watanabe, T.; Juvvadi, P.R.; Maruyama, J.-I.; Arioka, M.; Kitamoto, K. Double disruption of the proteinase genes, tppA and pepE, increases the production level of human lysozyme by Aspergillus oryzae. Appl. Microbiol. Biotechnol. 2007, 76, 1059. [Google Scholar] [CrossRef]
- Tsuchiya, K.; Tada, S.; Gomi, K.; Kitamoto, K.; Kumagai, C.; Jigami, Y.; Tamura, G. High level expression of the synthetic human lysozyme gene in Aspergillus oryzae. Appl. Microbiol. Biotechnol. 1992, 38, 109–114. [Google Scholar] [CrossRef]
- Volesky, B.; Luong, J.H.T.; Aunstrup, K. Microbial Enzymes: Production, Purification, and Isolation. Crit. Rev. Biotechnol. 1984, 2, 119–146. [Google Scholar] [CrossRef]
- Niyonzima, F.N.; Veena, S.M.; More, S. Industrial Production and Optimization of Microbial Enzymes. In Microorganisms for Sustainability; Kumar Arora, N., Mishra, J., Mishra, V., Eds.; Springer: Singapore, 2020; Volume 11, pp. 115–135. [Google Scholar]
- Idiris, A.; Tohda, H.; Kumagai, H.; Takegawa, K. Engineering of protein secretion in yeast: Strategies and impact on protein production. Appl. Microbiol. Biotechnol. 2010, 86, 403–417. [Google Scholar] [CrossRef] [PubMed]
- Mahalik, S.; Sharma, A.K.; Mukherjee, K.J. Genome engineering for improved recombinant protein expression in Escherichia coli. ISSN Int. Centre 2014, 13, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veerana, M.; Mitra, S.; Ki, S.-H.; Kim, S.-M.; Choi, E.-H.; Lee, T.; Park, G. Plasma-mediated enhancement of enzyme secretion in Aspergillus oryzae. Microb. Biotechnol. 2021, 14, 262–276. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.; Lin, B.; Xu, Y.; Yang, G.; Xu, J.; Zhang, S.; Li, R.; Wang, S. Recent developments in pasteurising seeds and their products using radio frequency heating: A review. Int. J. Food Sci. Technol. 2022, 57, 3223–3243. [Google Scholar] [CrossRef]
- Soto-Reyes, N.; Sosa-Morales, M.E.; Rojas-Laguna, R.; López-Malo, A. Advances in radio frequency pasteurisation equipment for liquid foods: A review. Int. J. Food Sci. Technol. 2022, 57, 3207–3222. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhao, W.; Yang, R.; Singh Sidhu, J.; Kong, F. Radio frequency heating to inactivate microorganisms in broccoli powder. Food Qual. Saf. 2017, 1, 93–100. [Google Scholar] [CrossRef]
- Hou, L.; Kou, X.; Li, R.; Wang, S. Thermal inactivation of fungi in chestnuts by hot air assisted radio frequency treatments. Food Control 2018, 93, 297–304. [Google Scholar] [CrossRef]
- Cui, B.; Sun, Y.; Wang, K.; Liu, Y.; Fu, H.; Wang, Y.; Wang, Y. Pasteurization mechanism on the cellular level of radio frequency heating and its possible non-thermal effect. IFSET 2022, 78, 103026. [Google Scholar] [CrossRef]
- Xu, J.; Yang, G.; Li, R.; Xu, Y.; Lin, B.; Wang, S. Effects of radio frequency heating on microbial populations and physicochemical properties of buckwheat. Int. J. Food Microbiol. 2022, 363, 109500. [Google Scholar] [CrossRef]
- Patruno, A.; Ferrone, A.; Costantini, E.; Franceschelli, S.; Pesce, M.; Speranza, L.; Amerio, P.; D’Angelo, C.; Felaco, M.; Grilli, A.; et al. Extremely low-frequency electromagnetic fields accelerates wound healing modulating MMP-9 and inflammatory cytokines. Cell Prolif 2018, 51, e12432. [Google Scholar] [CrossRef]
- Erica, C.; Aielli, L.; Serra, F.; De Dominicis, L.; Falasca, K.; Di Giovanni, P.; Reale, M. Evaluation of Cell Migration and Cytokines Expression Changes under the Radiofrequency Electromagnetic Field on Wound Healing In Vitro Model. Int. J. Mol. Sci. 2022, 23, 2205. [Google Scholar] [CrossRef]
- Rahardjo, Y.S.P.; Weber, F.J.; Haemers, S.; Tramper, J.; Rinzema, A. Aerial mycelia of Aspergillus oryzae accelerate α-amylase production in a model solid-state fermentation system. Enz. Microbial Technol. 2005, 36, 900–902. [Google Scholar] [CrossRef]
- Chutmanop, J.; Chuichulcherm, S.; Chisti, Y.; Srinophakun, P. Protease production by Aspergillus oryzae in solid-state fermentation using agroindustrial substrates. J. Chem. Technol. Biotechnol. 2008, 83, 1012–1018. [Google Scholar] [CrossRef]
- Hong, S.-B.; Hong, S.-Y.; Jo, K.-H.; Kim, Y.-S.; Do, J.-H.; Do, J.-Y.; Noh, S.-B.; Yoon, H.-H.; Chung, S.-H. Taxonomic character-ization and safety of Nuruk molds used industrially in Korea. Kor. J. Mycol. 2015, 43, 149–157. [Google Scholar]
- Awan, W.; Naqvi, S.; Naqvi, A.; Abbas, S.M.; Zaidi, A.; Hussain, N. Design and Characterization of Wideband Printed Antenna Based on DGS for 28 GHz 5G Applications. J. Electromagn. Eng. Sci. 2021, 21, 177–183. [Google Scholar] [CrossRef]
- Cha, J.; Leem, C.-S.; Kim, I.; Lee, H.; Lee, H. Broadband Dual-Polarized 2 × 2 MIMO Antenna for a 5G Wireless Communication System. Electronics 2021, 10, 2141. [Google Scholar] [CrossRef]
- Kim, I.; Lee, B. Wideband Antenna for High-Frequency 5G Wireless Communication. J. Electromagn. Eng. Sci. 2022, 22, 296–301. [Google Scholar] [CrossRef]
- Kim, I.; Lee, D.M.; Lee, Y.J.; Shin, J.W.; Kim, E.S.; Lee, H.; Kim, N.Y. Dual-Band On-Body Near Field Antenna for Measuring Deep Core Temperature With a Microwave Radiometer. IEEE Access 2022, 10, 63715–63722. [Google Scholar] [CrossRef]
- Yahya Alkhalaf, H.; Yazed Ahmad, M.; Ramiah, H. Self-Sustainable Biomedical Devices Powered by RF Energy: A Review. Sensors 2022, 22, 6371. [Google Scholar] [CrossRef]
- Balanis, C.A. Micorstrip and Mobile Communications Antennas. In Antenna Theory Analysis and Design, 4th ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016; pp. 789–858. [Google Scholar]
- Stutzman, W.L.; Thiele, G.A. Low-Profile Antennas. In Antenna Theory and Design, 3rd ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; pp. 465–530. [Google Scholar]
- Minagawa, H.; Onodera, K.; Fujita, H.; Sakamoto, T.; Akitomi, J.; Kaneko, N.; Shiratori, I.; Kuwahara, M.; Horii, K.; Waga, I. Selection, characterization and application of artificial DNA aptamer containing appended bases with sub-nanomolar affinity for a salivary biomarker. Sci. Rep. 2017, 7, 42716. [Google Scholar] [CrossRef] [Green Version]
- Veerana, M.; Lim, J.-S.; Choi, E.-H.; Park, G. Aspergillus oryzae spore germination is enhanced by non-thermal atmospheric pressure plasma. Sci. Rep. 2019, 9, 11184. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Kumar, A.; Baruah, D.; Tamuli, R. Calcium signaling is involved in diverse cellular processes in fungi. Mycology 2021, 12, 10–24. [Google Scholar] [CrossRef] [PubMed]
- Kou, X.; Li, R.; Hou, L.; Zhang, L.; Wang, S. Identifying possible non-thermal effects of radio frequency energy on inactivating food microorganisms. Int. J. Food Microbiol. 2018, 269, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, S.M.J.; Taheri, M.; Paknahad, M.; Khandadash, S. Effects of radiofrequency electromagnetic fields emitted from mobile phones and Wi-Fi router on the growth rate and susceptibility of Enterococcus faecalis to antibiotics. J. Biomed. Phys. Eng. 2022, 12, 387–394. [Google Scholar] [CrossRef]
- Lin, K.-W.; Yang, C.-J.; Lian, H.-Y.; Cai, P. Exposure of ELF-EMF and RF-EMF increase the rate of glucose transport and TCA cycle in budding yeast. Front. Microbiol. 2016, 7, 1378. [Google Scholar] [CrossRef] [Green Version]
- Bassett, C.A. Fundamental and practical aspects of therapeutic uses of pulsed electromagnetic fields (PEMFs). Crit. Rev. Biomed. Eng. 1989, 17, 451–529. [Google Scholar]
- Guo, L.; Kubat, N.J.; Nelson, T.R.; Isenberg, R.A. Meta-Analysis of Clinical Efficacy of Pulsed Radio Frequency Energy Treatment. Ann. Surg. 2012, 255, 457–467. [Google Scholar] [CrossRef]
- Aleem, B.; Rashid, M.H.; Zeb, N.; Saqib, A.; Ihsan, A.; Iqbal, M.; Ali, H. Random mutagenesis of super Koji (Aspergillus oryzae): Improvement in production and thermal stability of α-amylases for maltose syrup production. BMC Microbiol. 2018, 18, 200. [Google Scholar] [CrossRef] [Green Version]
- Krusen, F.H.; Ellwood, P.M.; Kottke, F.J. Handbook of Physical Medicine and Rehabilitation; Philadelphia & London: London, UK, 1965. [Google Scholar]
- Valberg, P.A. Radio frequency radiation (RFR): The nature of exposure and carcinogenic potential. Cancer Causes Control 1997, 8, 323–332. [Google Scholar] [CrossRef]
- Wood, A.; Karipidis, K. Radiofrequency Fields and Calcium Movements Into and Out of Cells. Radiat. Res. 2021, 195, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Murry, R.; Traxler, L.; Pötschner, J.; Krüger, T.; Kniemeyer, O.; Krause, K.; Kothe, E. Inositol Signaling in the Basidiomycete Fungus Schizophyllum commune. J. Fungi 2021, 7, 470. [Google Scholar] [CrossRef] [PubMed]
- Micaroni, M. The role of calcium in intracellular trafficking. Curr. Mol. Med. 2010, 10, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Martín, J.F. Calcium-containing phosphopeptides pave the secretory pathway for efficient protein traffic and secretion in fungi. Microb Cell Fact. 2014, 13, 117. [Google Scholar] [CrossRef] [PubMed]
- Saadi, S.; Alimohammadi, M.; Nabizadeh, R.; Mesdaghinia, A.; Aslani, H.; Nazmara, S.; Ghani, M.; Mahmoodi, B.; Asgari, M.; Mousavipour, N. Evaluating efficiency of radio waves for microbial removal in water samples. J. Adv. Environ. Health Res. 2014, 2, 157–164. [Google Scholar] [CrossRef]
- Kiontke, S.; Göbel, T.; Brych, A.; Batschauer, A. DASH-type cryptochromes—Solved and open questions. Biol. Chem. 2020, 401, 1487–1493. [Google Scholar] [CrossRef]
- Tagua, V.G.; Pausch, M.; Eckel, M.; Gutiérrez, G.; Miralles-Durán, A.; Sanz, C.; Eslava, A.P.; Pokorny, R.; Corrochano, L.M.; Batschauer, A. Fungal cryptochrome with DNA repair activity reveals an early stage in cryptochrome evolution. Proc. Natl. Acad. Sci. USA 2015, 112, 15130–15135. [Google Scholar] [CrossRef] [Green Version]
- Yoshii, T.; Ahmad, M.; Helfrich-Förster, C. Cryptochrome mediates light-dependent magnetosensitivity of Drosophila’s circadian clock. PLoS Biol. 2009, 7, e1000086. [Google Scholar] [CrossRef]
- Albaqami, M.; Hammad, M.; Pooam, M.; Procopio, M.; Sameti, M.; Ritz, T.; Ahmad, M.; Martino, C.F. Arabidopsis cryptochrome is responsive to Radiofrequency (RF) electromagnetic fields. Sci. Rep. 2020, 10, 11260. [Google Scholar] [CrossRef]
- Özgür, B.; Christoph, B.; Sven, K.; Paul, G.; Gerhard, H.B. More Than a Repair Enzyme: Aspergillus nidulans Photolyase-like CryA Is a Regulator of Sexual Development. Mol. Biol. Cell. 2008, 19, 3254–3262. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequences |
|---|---|
| α-amylase aptamer * | 5′-GGATACCTTAACGCCGCCTATTGTGAACGACGTGAATCGTGTTTGTGGGTCCGGAGTTGCACCCGTCTCGAAATC-3′ |
| Genes | Primer Sequences |
|---|---|
| α-Amylase | Forward-5′ACTGGGTGGGATCATTGGTA3′ Reverse-5′ACAAGTGTAGGCCGGATCAC3′ |
| GTPase (ER to Golgi), SAR1 homolog | Forward-5′CGAAGTGAGCGGTATCGTTT3′ Reverse-5′CCCTTTCCTGTGGTCTGGTA3′ |
| GTPase (cis to medial Golgi), YPT1 homolog | Forward-5′TGATGGCAAGACAGTGAAGC3′ Reverse-5′TTGACACCCTCAGTGGCATA3′ |
| AO090012000557 | Forward-5′CCAGTCTGCCTCCTCTGTCTAGC3′ Reverse-5′TGGTTGTGAACGAGCGTCTATCTTG3′ |
| AO090005001593 | Forward-5′TGGTGGCATTGAACTGGCAGAAC3′ Rverse-5′TCTAGCTGACGACGCACGATAGTAG3′ |
| AO090005001097 | Foward-5′ACAAAGGTGGAGACTGGAATGATGC3′ Rverse-5′TCGTGATTGAGGGATGCTTGGTTG3′ |
| 18S ribosomal RNA | Forward-5′GGAAACTCACCAGGTCCAGA3′ Reverse-5′AGCCGATAGTCCCCCTAAGA3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veerana, M.; Yu, N.-N.; Bae, S.-J.; Kim, I.; Kim, E.-S.; Ketya, W.; Lee, H.-Y.; Kim, N.-Y.; Park, G. Enhancement of Fungal Enzyme Production by Radio-Frequency Electromagnetic Fields. J. Fungi 2022, 8, 1187. https://doi.org/10.3390/jof8111187
Veerana M, Yu N-N, Bae S-J, Kim I, Kim E-S, Ketya W, Lee H-Y, Kim N-Y, Park G. Enhancement of Fungal Enzyme Production by Radio-Frequency Electromagnetic Fields. Journal of Fungi. 2022; 8(11):1187. https://doi.org/10.3390/jof8111187
Chicago/Turabian StyleVeerana, Mayura, Nan-Nan Yu, Si-Jin Bae, Ikhwan Kim, Eun-Seong Kim, Wirinthip Ketya, Hak-Yong Lee, Nam-Young Kim, and Gyungsoon Park. 2022. "Enhancement of Fungal Enzyme Production by Radio-Frequency Electromagnetic Fields" Journal of Fungi 8, no. 11: 1187. https://doi.org/10.3390/jof8111187