

Three New Trichoderma Species in Harzianum Clade Associated with the Contaminated Substrates of Edible Fungi

Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolates and Specimens
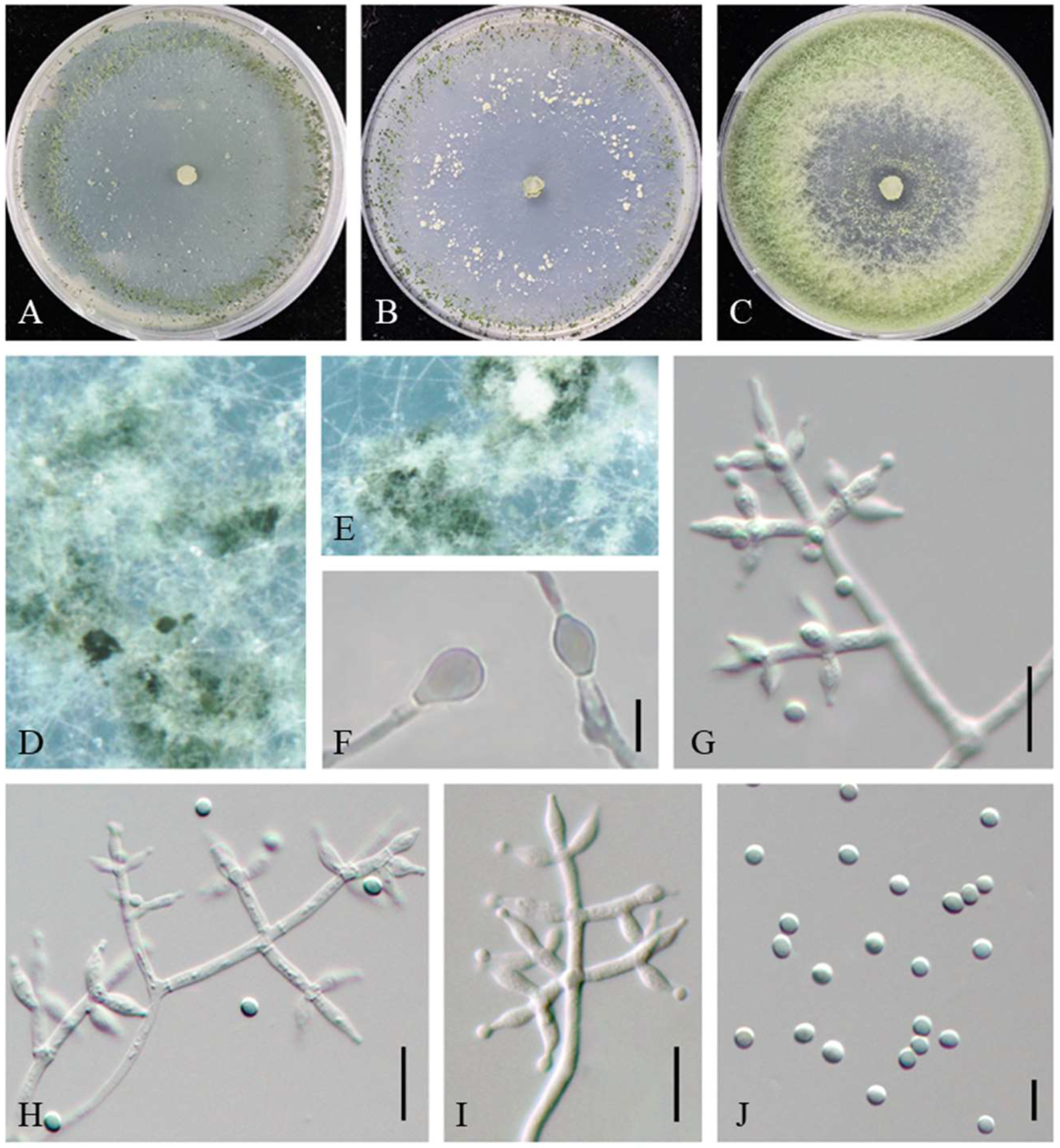
2.2. Morphology and Growth Characterization
2.3. DNA Extraction, PCR Amplification, and Sequencing
2.4. Phylogenetic Analyses
3. Results
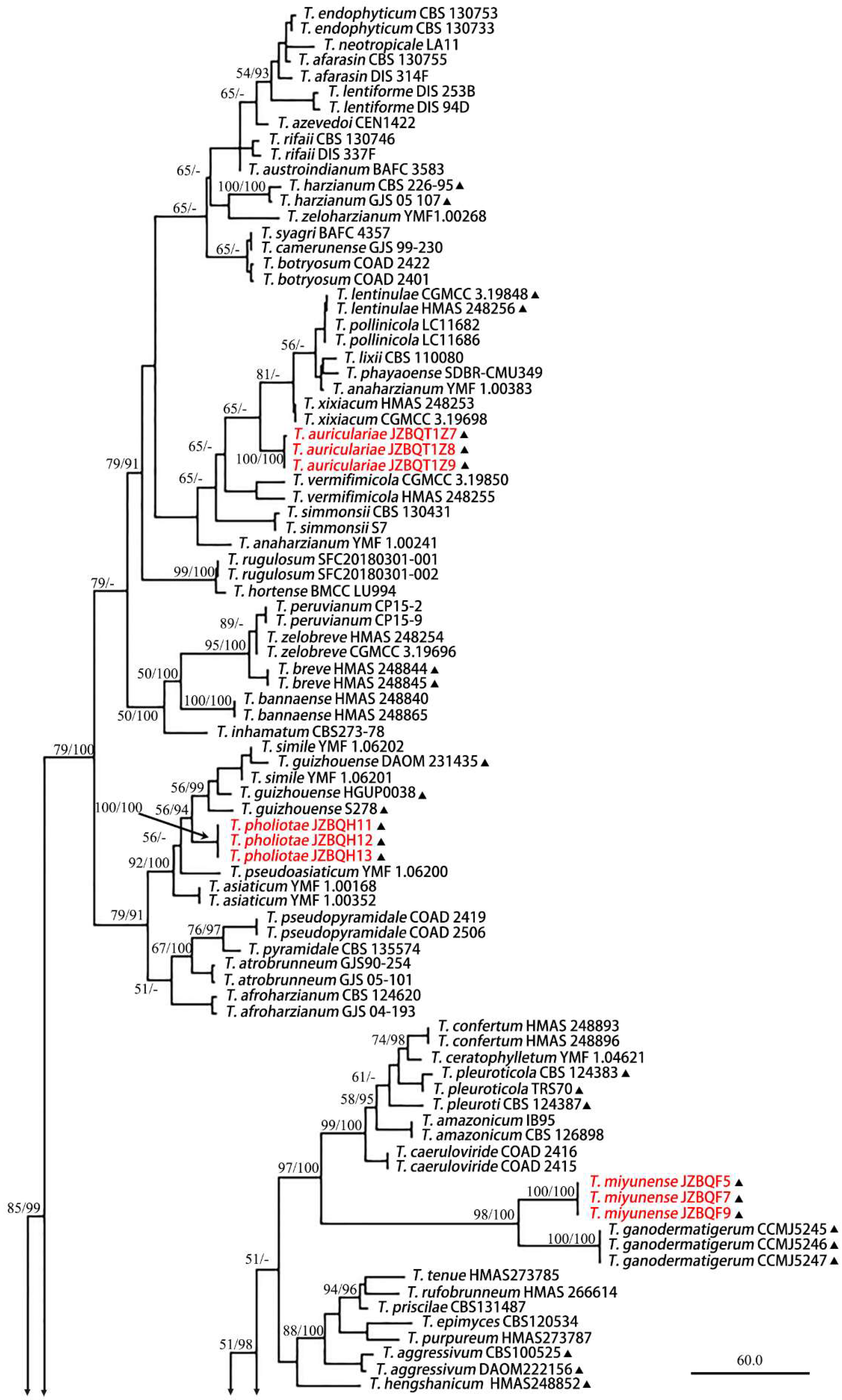
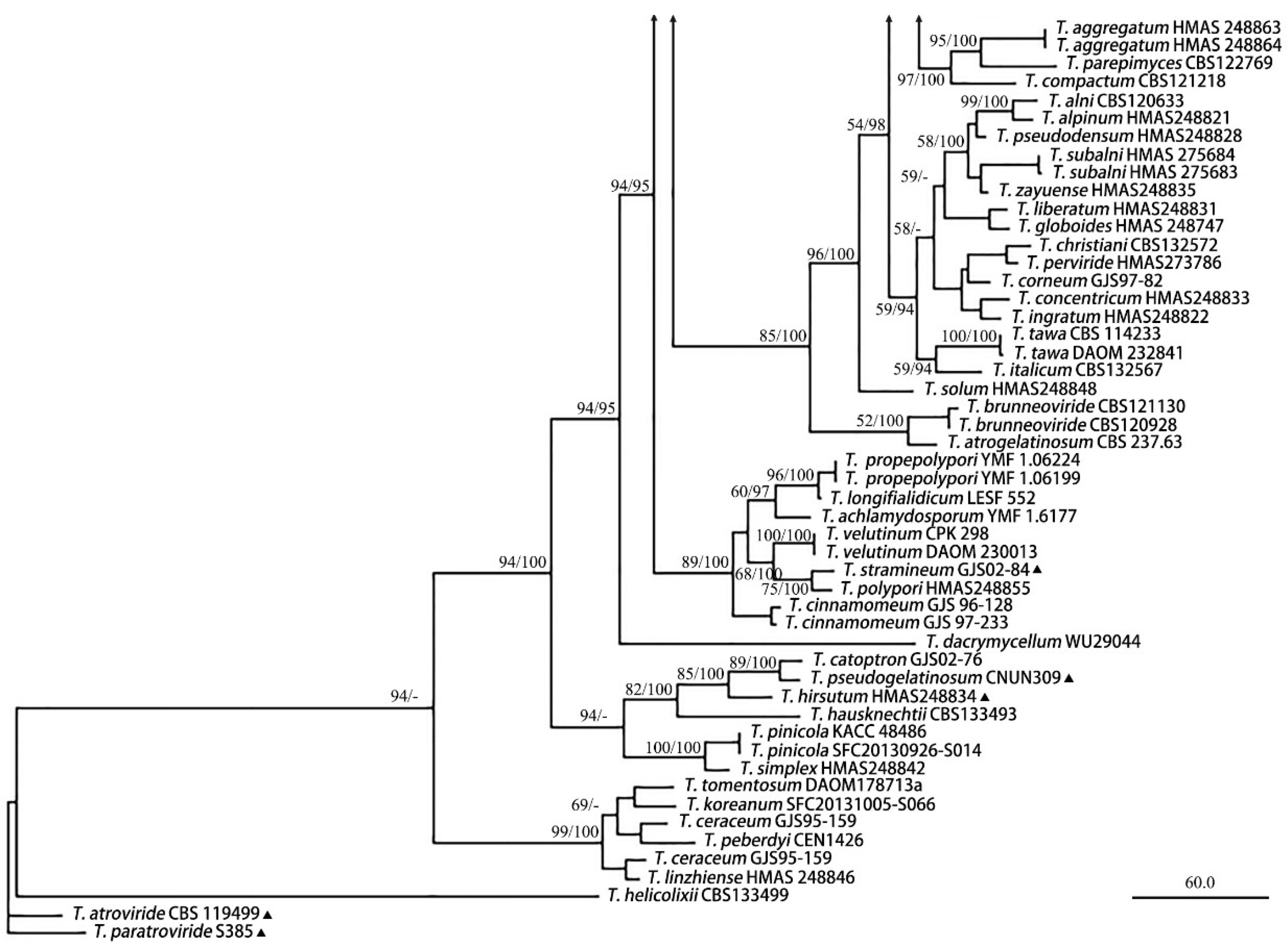
3.1. Phylogenetic Analyses
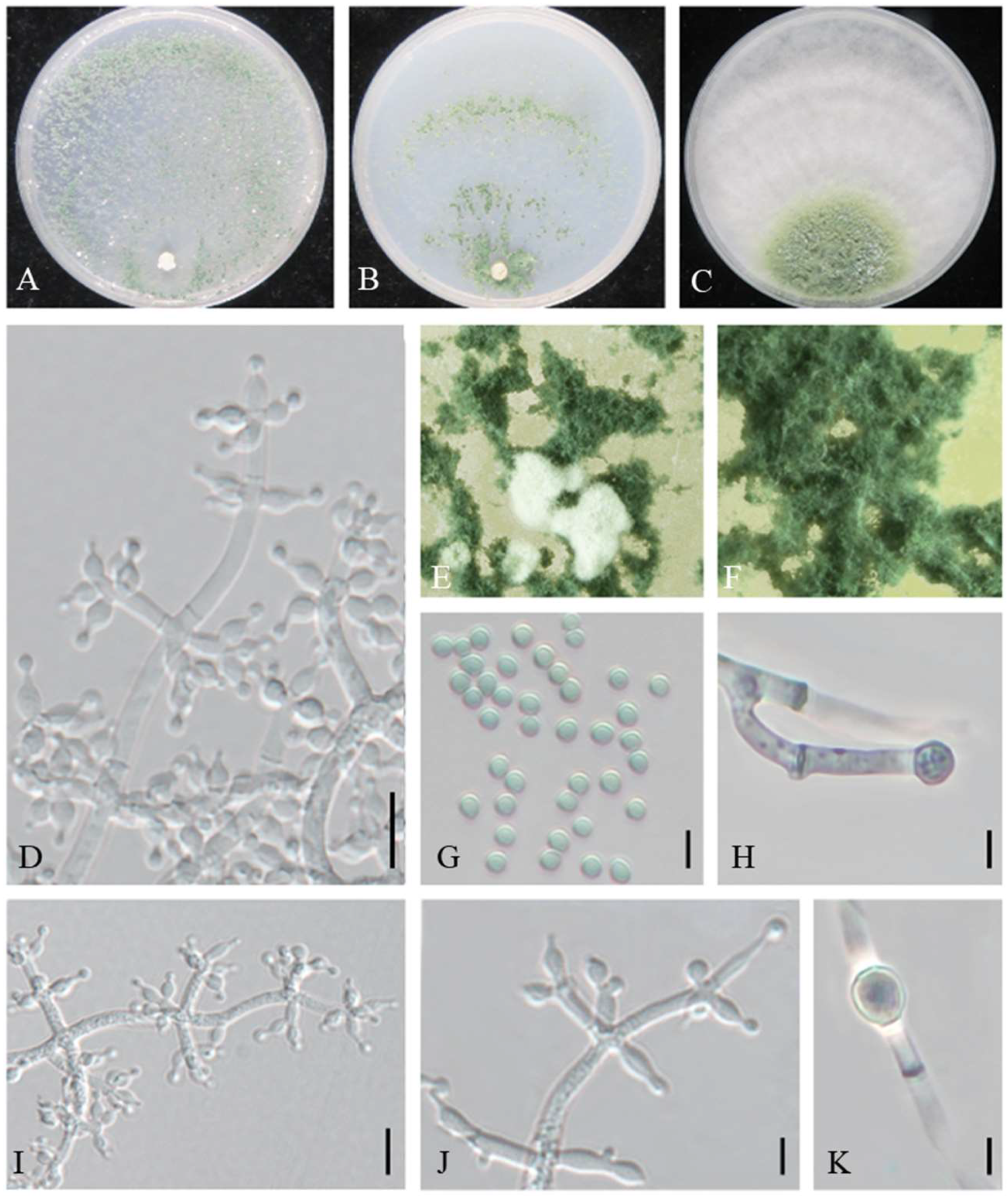
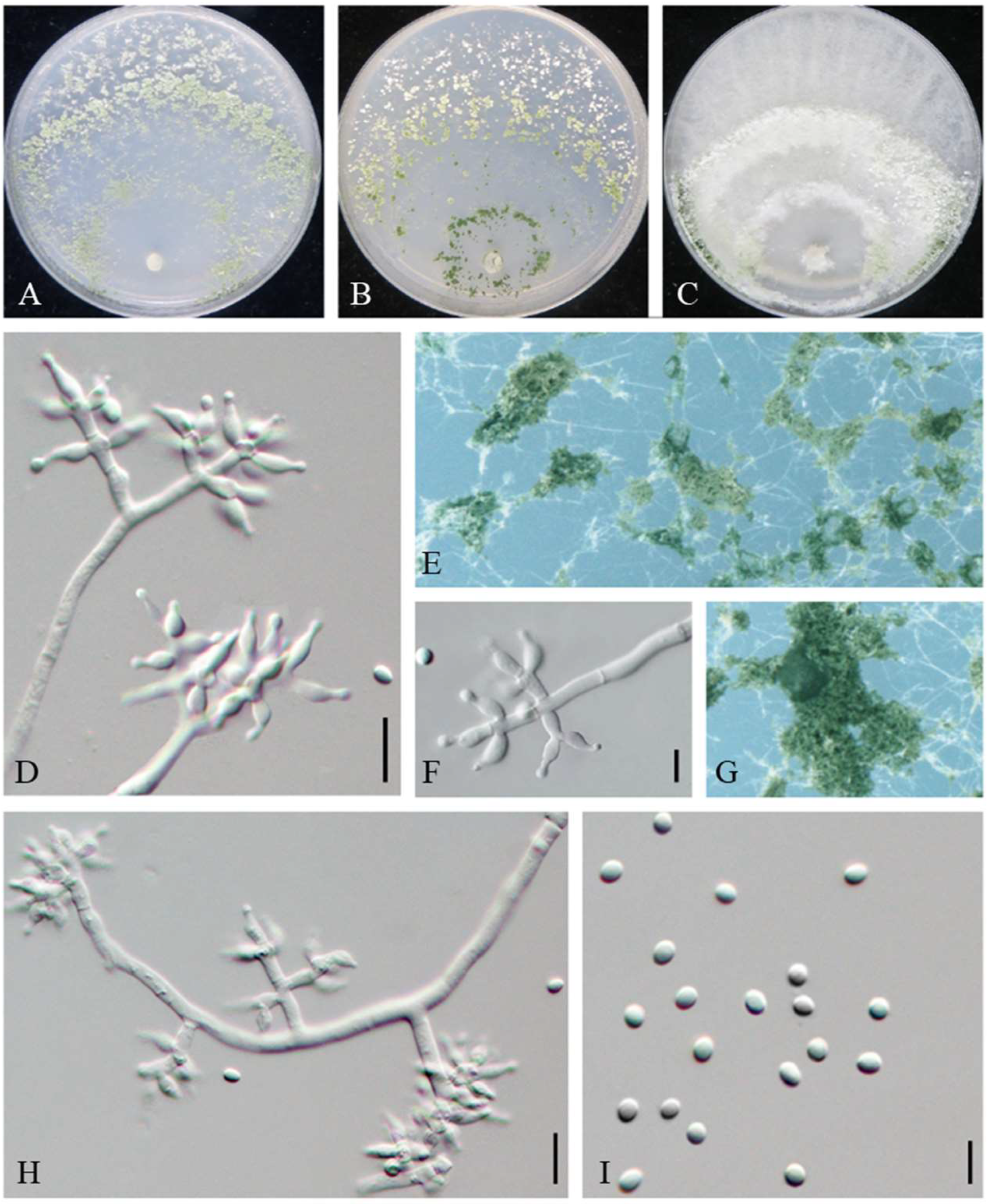
3.2. Taxonomy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cai, F.; Druzhinina, I.S. In honor of John Bissett: Authoritative guidelines on molecular identification of Trichoderma. Fungal Divers. 2021, 107, 1–69. [Google Scholar] [CrossRef]
- Druzhinina, I.S.; Seidl-Seiboth, V.; Herrera-Estrella, A.; Horwitz, B.A.; Kenerley, C.M.; Monte, E.; Mukherjee, P.K.; Zeilinger, S.; Grigoriev, I.V.; Kubicek, C.P. Trichoderma: The genomics of opportunistic success. Nat. Rev. Microbiol. 2011, 9, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Nuangmek, W.; Aiduang, W.; Kumla, J.; Lumyong, S.; Suwannarach, N. Evaluation of a newly identified endophytic fungus, Trichoderma phayaoense for plant growth promotion and biological control of gummy stem blight and wilt of muskmelon. Front. Microbiol. 2021, 12, 634772. [Google Scholar] [CrossRef]
- Carillo, P.; Woo, S.L.; Comite, E.; El-Nakhel, C.; Vinale, F. Application of Trichoderma harzianum, 6-pentyl-α-pyrone and plant biopolymer formulations modulate plant metabolism and fruit quality of plum tomatoes. Plants 2020, 9, 771. [Google Scholar] [CrossRef]
- Wei, H.; Wu, M.; Fan, A.; Su, H. Recombinant protein production in the filamentous fungus Trichoderma. Chin. J. Chem. Eng. 2021, 30, 74–81. [Google Scholar] [CrossRef]
- Gupta, S.; Smith, P.; Boughton, B.; Twt, R.; Sha, N.; Roessner, U. Inoculation of barley with Trichoderma harzianum T-22 modifies lipids and metabolites to improve salt tolerance. J. Exp. Bot. 2021, 72, 7229–7246. [Google Scholar] [CrossRef]
- Samuel, G.J.; Dodd, S.L.; Gams, W.; Castlebury, L.A.; Petrini, O. Trichoderma species associated with the green mold epidemic of commercially grown Agaricus bisporus. Mycologia 2002, 94, 146–170. [Google Scholar] [CrossRef]
- Yan, Y.; Zhang, C.; Moodley, O.; Zhang, L.; Xu, J. Green mold caused by Trichoderma atroviride on the lingzhi medicinal mushroom, Ganoderma lingzhi (Agaricomycetes). Int. J. Med. Mushrooms 2019, 21, 515–521. [Google Scholar] [CrossRef]
- Hatvani, L.; Antal, Z.; Manczinger, L.; Szekeres, A.; Druzhinina, I.S.; Kubicek, C.P.; Nagy, A.; Nagy, E.; Vagvolgyi, C.; Kredics, L. Green mold diseases of Agaricus and Pleurotus spp. are caused by related but phylogenetically different Trichoderma species. Phytopathology 2007, 97, 532–537. [Google Scholar] [CrossRef] [Green Version]
- Komon-Zelazowska, M.; Bissett, J.; Zafari, D.; Hatvani, L.; Manczinger, L.; Woo, S.; Lorito, M.; Kredics, L.; Kubicek, C.P.; Druzhinina, I.S. Genetically closely related but phenotypically divergent Trichoderma species cause green mold disease in oyster mushroom farms worldwide. Appl. Environ. Microb. 2007, 73, 7415–7426. [Google Scholar] [CrossRef]
- Kim, J.Y.; Yun, Y.H.; Hyun, M.W.; Kim, M.H.; Kim, S.H. Identification and characterization of Gliocladium viride isolated from mushroom fly infested oak log beds used for shiitake cultivation. Mycobiology 2010, 38, 7–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, H.; Qiao, M.; Lv, Y.F.; Du, X.; Zhang, K.Q.; Yu, Z.F. New species of Trichoderma isolated as endophytes and saprobes from southwest China. J. Fungi 2021, 7, 467. [Google Scholar] [CrossRef] [PubMed]
- Ignazio, C.; Linda, M.K. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar]
- Jaklitsch, W.M.; Komon, M.; Kubicek, C.P.; Druzhinina, I.S. Hypocrea voglmayrii sp. nov. from the Austrian Alps represents a new phylogenetic clade in Hypocrea/Trichoderma. Mycologia 2005, 97, 1365–1378. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.X.; Zhuang, W.Y. Trichoderma (Hypocrea) species with green ascospores from China. Persoonia 2015, 34, 113–129. [Google Scholar] [CrossRef] [Green Version]
- Dou, K.; Lu, Z.; Wu, Q.; Ni, M.; Yu, C.; Wang, M.; Li, Y.; Wang, X.; Xie, H.; Chen, J.; et al. MIST: A multilocus identification system for Trichoderma. Appl. Environ. Microb. 2020, 86, e01532-20. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acids Symp. 1999, 41, 95–98. [Google Scholar]
- Higgins, D.G.; Jeanmougin, F.; Gibson, T.J.; Plewniak, F.; Thompson, J.D. The Clustal X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Symp. 1997, 25, 4876–4882. [Google Scholar]
- Swofford, D.L. PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods); Version 4.0; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author. 2004. Available online: http://paup.csit.fsu.edu (accessed on 1 January 2018).
- Rambaut, A. FigTree. Tree Figure Drawing Tool, v. 1.4.3. 2016. Available online: http://tree.bio.ed.ac.uk/ (accessed on 1 January 2018).
- Gu, X.; Wang, R.; Sun, Q.; Wu, B.; Sun, J.-Z. Four new species of Trichoderma in the Harzianum clade from northern China. Mycokeys 2020, 73, 109–132. [Google Scholar] [CrossRef] [PubMed]
- Chaverri, P.; Branco-Rocha, F.; Jaklitsch, W.; Gazis, R.; Degenkolb, T.; Samuels, G.J. Systematics of the Trichoderma harzianum species complex and the re-identification of commercial biocontrol strains. Mycologia 2015, 107, 558–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, H.S.; Lu, X.; Dai, Y.C.; Hyde, K.V.D.; Kan, Y.H.; Kusan, I.; He, S.H.; Liu, N.G.; Sarma, V.V.; Zhao, C.L.; et al. Fungal diversity notes 1277–1386: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2020, 104, 1–266. [Google Scholar] [CrossRef]
- An, X.Y.; Cheng, G.H.; Gao, H.X.; Li, D.; Li, Y. Phylogenetic analysis of trichoderma species associated with green mold disease on mushrooms and two new pathogens on Ganoderma sichuanense. J. Fungi 2022, 8, 704. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, M.D.H.; Evans, H.C.; De Abreu, L.M.; De Macedo, D.M.; Ndacnou, M.K.; Bekele, K.B.; Barreto, R.W. New species and records of Trichoderma isolated as mycoparasites and endophytes from cultivated and wild coffee in Africa. Sci. Rep. 2021, 11, 5671. [Google Scholar] [CrossRef]
- Chen, K.; Zhuang, W.Y. Seven new species of Trichoderma from soil in China. Mycosystema 2017, 36, 1441–1462. [Google Scholar]
- Chaverri, P.; Gazis, R.O.; Samuels, G.J. Trichoderma amazonicum, a new endophytic species on Hevea brasiliensis and H. guianensis from the Amazon basin. Mycologia 2011, 103, 139–151. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.R.; Tan, P.; Jiang, Y.L.; Hyde, K.D.; Mckenzie, E.H.C.; Bahkali, A.H.; Kang, J.C.; Wang, Y. A novel Trichoderma species isolated from soil in Guizhou, T. guizhouense. Mycol. Prog. 2013, 12, 167–172. [Google Scholar] [CrossRef]
- Chaverri, P.; Samuels, G.J. Hypocrea/Trichoderma (Ascomycota, Hypocreales, Hypocreaceae): Species with green ascospores. Stud. Mycol. 2003, 48, 1–116. [Google Scholar]
- Jaklitsch, W.M.; Samuels, G.J.; Dodd, S.L.; Lu, B.S.; Druzhinina, I.S. Hypocrea rufa/Trichoderma viride: A reassessment, and description of five closely related species with and without warted conidia. Stud. Mycol. 2006, 56, 135–177. [Google Scholar] [CrossRef] [Green Version]
- Luecking, R.; Aime, M.C.; Robbertse, B.; Miller, A.N.; Ariyawansa, H.A.; Aoki, T.; Cardinali, G.; Crous, P.W.; Druzhinina, I.S.; Geiser, D.M.; et al. Unambiguous identification of fungi: Where do we stand and how accurate and precise is fungal DNA barcoding? IMA Fungus 2020, 11, 14. [Google Scholar] [CrossRef] [PubMed]
- Druzhinina, I.; Kubicek, C.P. Species concepts and biodiversity in Trichoderma and Hypocrea: From aggregate species to species clusters? J. Zhejiang Univ. Sci. B 2005, 6, 100–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Persoon, C.H. Neurospora genetic nomenclature. Romers Neues Mag. Bot. 1794, 1, 81–128. [Google Scholar]
- Jhune, C.S.; Leem, H.T.; Park, H.S.; Lee, C.J.; Weon, H.Y.; Seok, S.J.; Yoo, K.H.; Sung, G.H. Identification of oyster mushroom green mold pathogen that causes and pathological characteristics. J. Mushrooms 2014, 12, 132–137. [Google Scholar] [CrossRef] [Green Version]
- Barrera, V.A.; Iannone, L.; Romero, A.I.; Chaverri, P. Expanding the Trichoderma harzianum species complex: Three new species from Argentine natural and cultivated ecosystems. Mycologia 2021, 113, 1136–1155. [Google Scholar] [CrossRef]
- Allaga, H.; Zhumakayev, A.; Buchner, R.; Kocsube, S.; Szucs, A.; Vagvolgyi, C.; Kredics, L.; Hatvani, L. Members of the Trichoderma harzianum species complex with mushroom pathogenic potential. Agronomy 2021, 11, 2434. [Google Scholar] [CrossRef]
- Kosanovic, D.; Potocnik, I.; Duduk, B.; Vukojevic, J.; Stajic, M.; Rekanovic, E.; Milijasevic-Marcic, S. Trichoderma species on Agaricus bisporus farms in Serbia and their biocontrol. Ann. Appl. Biol. 2013, 163, 218–230. [Google Scholar] [CrossRef]
- Marik, T.; Urban, P.; Tyagi, C.; Szekeres, A.; Leitgeb, B.; Vagvolgyi, M.; Manczinger, L.; Druzhinina, I.S.; Vagvolgyi, C.; Kredics, L. Diversity profile and dynamics of peptaibols produced by green mould Trichoderma species in interactions with their hosts Agaricus bisporus and Pleurotus ostreatus. Chem. Biodivers. 2017, 14, e1700033. [Google Scholar] [CrossRef]
- Wu, X.J.; Hu, F.P.; He, H.Z.; Xie, B.G. Identification of Trichoderma species associated with cultivated edible fungi. J. Agric. Biotechnol. 2008, 16, 1048–1055. [Google Scholar]
- Kim, C.S.; Park, M.S.; Kim, S.C.; Maekawa, N.; Yu, S.H. Identification of Trichoderma, a competitor of shiitake mushroom (Lentinula edodes), and competition between Lentinula edodes and Trichoderma species in Korea. Plant Pathol. J. 2012, 28, 137–148. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.H. Isolation, Identification and Diversity Analysis of the Contaminating Fungi from the Edible Mushroom-Growing Synthetic Wood Logs. Master’s Thesis, Liaoning Normal University, Dalian, China, 2017. [Google Scholar]
- Wang, Y. Identification of the Pathogen of Lentinus edodes Sticks Rot and Preliminary Study on Its Occurrence. Master’s Thesis, Guizhou University, Guiyang, China, 2021. [Google Scholar]
- Samuels, G.J.; Ismaiel, A.; Mulaw, T.B.; Szakacs, G.; Druzhinina, I.S.; Kubicek, C.P.; Jaklitsch, W.M. The Longibrachiatum clade of Trichoderma: A revision with new species. Fungal Divers. 2012, 55, 77–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, M.S.; Seo, G.S.; Bae, K.S.; Yu, S.H. Characterization of Trichoderma spp. associated with green mold of oyster mushroom by PCR-RFLP and sequence analysis of ITS regions of rDNA. Plant Pathol. J. 2005, 21, 229–236. [Google Scholar] [CrossRef]
- Innocenti, G.; Montanari, M.; Righini, H.; Roberti, R. Trichoderma species associated with green mould disease of Pleurotus ostreatus and their sensitivity to prochloraz. Plant Pathol. 2019, 68, 392–398. [Google Scholar] [CrossRef]
- Song, X.X.; Wang, Q.; Jun, J.X.; Zhang, J.J.; Chen, H.; Chen, M.J.; Huang, J.C.; Xie, B.Q. Study on accurate identification of four Trichoderma diseases in Agaricus bisporus in factory cultivation. Edible Fungi 2019, 41, 67–72. [Google Scholar]
- Choi, I.Y.; Choi, J.N.; Hyu, L.W.; Sharma, P.K. Isolation and identification of mushroom pathogens from Agrocybe aegerita. Mycobiology 2010, 38, 310–315. [Google Scholar] [CrossRef] [Green Version]
- Cai, M.; Idrees, M.; Zhou, Y.; Zhang, C.; Xu, J. First report of green mold disease caused by Trichoderma hengshanicum on Ganoderma lingzhi. Mycobiology 2020, 48, 427–430. [Google Scholar] [CrossRef]
- Chen, X.; Zhou, X.; Zhao, J.; Tang, X.; Pasquali, M.; Migheli, Q.; Berg, G.; Cernava, T. Occurrence of green mold disease on Dictyophora rubrovolvata caused by Trichoderma koningiopsis. J. Plant Pathol. 2021, 103, 981–984. [Google Scholar] [CrossRef]
- Kim, S.W.; Kim, S.; Lee, H.J.; Park, J.W.; Ro, H.S. Isolation of fungal pathogens to an edible mushroom, Pleurotus eryngii, and development of specific ITS primers. Mycobiology 2013, 41, 252–255. [Google Scholar] [CrossRef]
- Wang, G.; Cao, X.; Ma, X.; Guo, M.; Liu, C.; Yan, L.; Bian, Y. Diversity and effect of Trichoderma spp. associated with green mold disease on Lentinula edodes in China. Microbiologyopen 2016, 5, 709–718. [Google Scholar] [CrossRef] [Green Version]
- Hua, R.; Li, J.Y.; Liu, S.X.; Luo, X.K.; Wang, X.Y.; Liu, C.L.; Zhang, L.Y.; Sun, D.F. Investigation on common dieases of Pleurotus ostreatus and identification of pathogens. Edible Fungi China 2021, 40, 80–86. [Google Scholar]
- Park, M.S.; Bae, K.S.; Yu, S.H. Two new species of Trichoderma associated with green mold of oyster mushroom cultivation in Korea. Mycobiology 2006, 34, 111–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazaki, K.; Tsuchiya, Y.; Okuda, T. Specific PCR assays for the detection of Trichoderma harzianum causing green mold disease during mushroom cultivation. Mycoscience 2009, 50, 94–99. [Google Scholar] [CrossRef]
- Kim, C.S.; Yu, S.H.; Nakagiri, A.; Shirouzu, T.; Sotome, K.; Kim, S.C.; Maekawa, N. Re-evaluation of Hypocrea pseudogelatinosa and H. pseudostraminea isolated from shiitake mushroom (Lentinula edodes) cultivation in Korea and Japan. Plant Pathol. J. 2012, 28, 341–356. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.S.; Shirouzu, T.; Nakagiri, A.; Sotome, K.; Maekawa, N. Trichoderma eijii and T. pseudolacteum, two new species from Japan. Mycol. Prog. 2013, 12, 739–753. [Google Scholar] [CrossRef]
- Lan, B.M. Isolation and identification of Trichoderma on oyster mushroom in Quanzhou. Chin. J. Trop. Agric. 2022, 42, 81–85. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Voucher | GenBank Accession Number | |
|---|---|---|---|
| rpb2 | tef1-α | ||
| T. achlamydosporum | YMF 1.6177 | MT052180 | MT070156 |
| T. afarasin | CBS 130755 | – | AF348093 |
| T. afarasin | DIS 314F | FJ442778 | FJ463400 |
| T. afroharzianum | CBS 124620 ET | FJ442691 | FJ463301 |
| T. afroharzianum | GJS 04-193 | FJ442709 | FJ463298 |
| T. aggregatum | HMAS 248863 | KY688001 | KY688062 |
| T. aggregatum | HMAS 248864 | KY688002 | KY688063 |
| T. aggressivum | CBS 100525 | AF545541 | AF348095 |
| T. aggressivum | DAOM 222156 ET | FJ442752 | AF348098 |
| T. alni | CBS 120633 ET | EU498349 | EU498312 |
| T. alpinum | HMAS 248821 T | KY687958 | KY688012 |
| T. amazonicum | IB95 | HM142368 | HM142377 |
| T. amazonicum | CBS126898 ET | HM142367 | HM142376 |
| T. anaharzianum | YMF 1.00241 | MH262577 | MH236493 |
| T. anaharzianum | YMF 1.00383 T | MH158995 | MH183182 |
| T. asiaticum | YMF 1.00168 | MH262575 | MH236492 |
| T. asiaticum | YMF 1.00352 T | MH158994 | MH183183 |
| T. atrobrunneum | GJS90-254 | FJ442735 | AF443943 |
| T. atrobrunneum | GJS 05-101 | FJ442745 | FJ463392 |
| T. atrogelatinosum | CBS 237.63 ET | KJ842201 | – |
| T. atroviride | CBS 119499 | FJ860518 | FJ860611 |
| T. auriculariae | JZBQT1Z7 T | ON649949 | ON649896 |
| T. auriculariae | JZBQT1Z8 | ON649950 | ON649897 |
| T. auriculariae | JZBQT1Z9 | ON649951 | ON649898 |
| T. austroindianum | BAFC 3583 | – | MH352421 |
| T. azevedoi | CEN1422 | MK696821 | MK696660 |
| T. bannaense | HMAS 248840 T | KY687979 | KY688037 |
| T. bannaense | HMAS 248865 | KY688003 | KY688038 |
| T. botryosum | COAD 2422 | MK044212 | MK044119 |
| T. botryosum | COAD 2401 | MK044181 | MK044088 |
| T. breve | HMAS 248844 T | KY687983 | KY688045 |
| T. breve | HMAS 248845 | KY687984 | KY688046 |
| T. brunneoviride | CBS 121130 | EU498357 | EU498316 |
| T. brunneoviride | CBS 120928 | EU498358 | EU498318 |
| T. caeruloviride | COAD 2416 | MK044201 | MK044108 |
| T. caeruloviride | COAD 2415 | MK044202 | MK044109 |
| T. camerunense | GJS 99-230 | – | AF348107 |
| T. catoptron | GJS 02-76 T | AY391900 | AY737726 |
| T. ceraceum | GJS 95-159 | AF545508 | AY937437 |
| T. ceratophylletum | YMF 1.04621 T | MK327580 | MK327579 |
| T. cerinum | DAOM 230012 | KJ842184 | KJ871242 |
| T. christiani | CBS 132572 ET | KJ665244 | KJ665439 |
| T. cinnamomeum | GJS 96-128 | AY391916 | AY391977 |
| T. cinnamomeum | GJS 97-233 | AY391919 | AY391978 |
| T. compactum | CBS 121218 | KF134789 | KF134798 |
| T. concentricum | HMAS 248833 | KY687971 | KY688027 |
| T. confertum | HMAS 248893 | MF371203 | MF371218 |
| T. confertum | HMAS 248896 | MF371205 | MF371220 |
| T. corneum | GJS 97-82 ET | KJ665252 | KJ665455 |
| T. dacrymycellum | WU29044 | FJ860533 | FJ860633 |
| T. endophyticum | CBS 130753 | FJ442722 | FJ463326 |
| T. endophyticum | CBS 130733 | FJ442690 | FJ463330 |
| T. epimyces | CBS120534 ET | EU498360 | EU498320 |
| T. ganodermatigerum | CCMJ5245 T | ON567189 | ON567195 |
| T. ganodermatigerum | CCMJ5246 | ON567190 | ON567196 |
| T. ganodermatigerum | CCMJ5247 | ON567191 | ON567197 |
| T. globoides | HMAS 248747 | KX026963 | KX026955 |
| T. guizhouense | HGUP0038 T | JQ901400 | JN215484 |
| T. guizhouense | S278 | KF134791 | KF134799 |
| T. guizhouense | DAOM 231435 | – | EF191321 |
| T. harzianum | CBS 226-95 | AF545549 | AF348101 |
| T. harzianum | GJS 05 107 | FJ442708 | FJ463329 |
| T. hausknechtii | CBS 133493 | KJ665276 | KJ665515 |
| T. helicolixii | CBS 133499 | KJ665278 | KJ665517 |
| T. hengshanicum | HMAS 248852 T | KY687991 | KY688054 |
| T. hirsutum | HMAS 248834 T | KY687972 | KY688029 |
| T. hortense | BMCC LU994 | – | KJ871185 |
| T. ingratum | HMAS 248822 | KY687973 | KY688018 |
| T. inhamatum | CBS 273-78 | FJ442725 | AF348099 |
| T. italicum | CBS 132567 | KJ665282 | KJ665525 |
| T. koreanum | SFC20131005-S066 | MH025988 | MH025979 |
| T. lentiforme | DIS 253B | FJ442756 | FJ851875 |
| T. lentiforme | DIS 94D | FJ442749 | FJ463379 |
| T. lentinulae | HMAS 248256 | MN605867 | MN605878 |
| T. lentinulae | CGMCC 3.19848 | MN605868 | MN605879 |
| T. liberatum | HMAS 248831 T | KY687969 | KY688025 |
| T. linzhiense | HMAS 248846 T | KY687985 | KY688047 |
| T. lixii | CBS 110080 | KJ665290 | FJ716622 |
| T. longifialidicum | LESF 552 | KT278955 | KT279020 |
| T. miyunense | JZBQF5 | ON649968 | ON649915 |
| T. miyunense | JZBQF7 T | ON649969 | ON649916 |
| T. miyunense | JZBQF9 | ON649970 | ON649917 |
| T. neotropicale | LA11 ET | – | HQ022771 |
| T. paratroviride | S385 | KJ665321 | KJ665627 |
| T. parepimyces | CBS 122769 ET | FJ860562 | FJ860664 |
| T. peberdyi | CEN1426 | MK696825 | MK696664 |
| T. peruvianum | CP15-2 | MW480153 | MW480145 |
| T. peruvianum | CP15-9 | MW480154 | MW480146 |
| T. perviride | HMAS 273786 | KX026962 | KX026954 |
| T. phayaoense | SDBR-CMU349 | MW002074 | MW002073 |
| T. pholiotae | JZBQH11 | ON649971 | ON649918 |
| T. pholiotae | JZBQH12 T | ON649972 | ON649919 |
| T. pholiotae | JZBQH13 | ON649973 | ON649920 |
| T. pinicola | KACC 48486 ET | MH025993 | MH025981 |
| T. pinicola | SFC20130926-S014 | MH025991 | MH025978 |
| T. pleuroti | CBS 124387 ET | HM142372 | HM142382 |
| T. pleuroticola | CBS 124383 ET | HM142371 | HM142381 |
| T. pleuroticola | TRS70 ET | KP009172 | KP008951 |
| T. pollinicola | LC11682 = LF1542 ET | MF939604 | MF939619 |
| T. pollinicola | LC11686 = LF2050 | MF939605 | MF939620 |
| T. polypori | HMAS 248855 T | KY687994 | KY688058 |
| T. priscilae | CBS 131487 ET | KJ665333 | KJ665691 |
| T. propepolypori | YMF 1.06224 T | MT052181 | MT070158 |
| T. propepolypori | YMF 1.06199 | MT052182 | MT070157 |
| T. pseudoasiaticum | YMF 1.06200 T | MT052183 | MT070155 |
| T. pseudodensum | HMAS 248828 T | KY687967 | KY688023 |
| T. pseudogelatinosum | CNUN309 ET | HM920173 | HM920202 |
| T. pseudopyramidale | COAD 2419 | MK044206 | MK044113 |
| T. pseudopyramidale | COAD 2506 | MK044207 | MK044114 |
| T. purpureum | HMAS 273787 T | KX026961 | KX026953 |
| T. pyramidale | CBS 135574 ET | KJ665334 | KJ665699 |
| T. rifaii | CBS 130746 | – | FJ463324 |
| T. rifaii | DIS 337F | FJ442720 | FJ463321 |
| T. rufobrunneum | HMAS 266614 T | KF730010 | KF729989 |
| T. rugulosum | SFC20180301-001 T | MH025986 | MH025984 |
| T. rugulosum | SFC20180301-002 | MH025987 | MH025985 |
| T. simile | YMF 1.06201 T | MT052184 | MT070154 |
| T. simile | YMF 1.06202 | MT052185 | MT070153 |
| T. simmonsii | CBS 130431 | FJ442757 | AF443935 |
| T. simmonsii | S7 | KJ665337 | KJ665719 |
| T. simplex | HMAS 248842 T | KY687981 | KY688041 |
| T. solum | HMAS 248848 T | KY687987 | KY688050 |
| T. stramineum | GJS 02-84 | AY391945 | AY391999 |
| T. subalni | HMAS 275683 | MH612371 | MH612377 |
| T. subalni | HMAS 275684 | MH612370 | MH612376 |
| T. syagri | BAFC 4357 | – | MG822711 |
| T. tawa | CBS 114233 ET | AY391956 | FJ463313 |
| T. tawa | DAOM 232841 | KJ842187 | EU279972 |
| T. tenue | HMAS 273785 ET | KX026960 | KX026952 |
| T. tomentosum | DAOM 178713a | AF545557 | AY750882 |
| T. velutinum | CPK 298 | KF134794 | KJ665769 |
| T. velutinum | DAOM 230013 ET | JN133569 | AY937415 |
| T. vermifimicola | CGMCC 3.19850 | MN605870 | MN605881 |
| T. vermifimicola | HMAS 248255 | MN605871 | MN605882 |
| T. xixiacum | HMAS 248253 T | MN605874 | MN605885 |
| T. xixiacum | CGMCC 3.19698 | MN605875 | MN605886 |
| T. zayuense | HMAS 248835 T | KY687974 | KY688031 |
| T. zelobreve | HMAS 248254 T | MN605872 | MN605883 |
| T. zelobreve | CGMCC 3.19696 | MN605873 | MN605884 |
| T. zeloharzianum | YMF 1.00268 | MH158996 | MH183181 |
| Species | Cultivated Mushroom | Reference |
|---|---|---|
| T. aggressivum | Agaricus bisporus | [40,41] |
| T. asperellum | A. bisporus | [9,42] |
| T. atroviride | L. edodes, Pleurotus ostreatus, A. bisporus, Ganoderma lingzhi | [8,9,43] |
| T. aureoviride | Auricularia heimuer, Flammulina filiformis, L. edodes | [44] |
| T. breve | L. edodes | [45] |
| T. capillare | Agaricus sp. | [46] |
| T. citrinviride | L. edodes, P. ostreatus | [43,47] |
| T. deliquescens | L. edodes | [11] |
| T. ganodermatigerum | G. sichuanense | [27] |
| T. ghanense | A. bisporus | [9] |
| T. guizhouense | P. ostreatus | [48] |
| T. hamatum | A. bisporus | [49] |
| T. harzianum | L. edodes, A. bisporus, P. ostreatus, Agrocybe aegerita | [43,50] |
| T. hengshanicum | G. lingzhi | [51] |
| T. hirsutum | L. edodes | [45] |
| T. koningii | P. ostreatus, A. bisporus | [37,40] |
| T. koningiopsis | Dictyophora rubrovolvata, P. eryngii | [52,53] |
| T. lentinulae | L. edodes | [24] |
| T. longibrachiatum | L. edodes, P. ostreatus, A. aegerita | [9,43,50] |
| T. oblongisporum | L. edodes | [54] |
| T. patella | P. ostreatus | [55] |
| T. pleuroti | P. ostreatus | [56] |
| T. pleuroticola | P. ostreatus, L. edodes, G. lingzhi | [50,54,56] |
| T. polysporum | L. edodes | [57] |
| T. pseudogelatinosum | L. edodes | [58] |
| T. pseudokoningii | P. ostreatus | [37] |
| T. pseudolacteum | L. edodes | [59] |
| T. pseudostramineum | L. edodes | [58] |
| T. reesei | P. ostreatus | [60] |
| T. stramineum | L. edodes | [57] |
| T. stromaticum | A. bisporus | [49] |
| T. virens | P. ostreatus, A. bisporus | [37,40] |
| T. viride | L. edodes | [54] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, Z.-J.; Qin, W.-T.; Zhao, J.; Liu, Y.; Wang, S.-X.; Zheng, S.-Y. Three New Trichoderma Species in Harzianum Clade Associated with the Contaminated Substrates of Edible Fungi. J. Fungi 2022, 8, 1154. https://doi.org/10.3390/jof8111154
Cao Z-J, Qin W-T, Zhao J, Liu Y, Wang S-X, Zheng S-Y. Three New Trichoderma Species in Harzianum Clade Associated with the Contaminated Substrates of Edible Fungi. Journal of Fungi. 2022; 8(11):1154. https://doi.org/10.3390/jof8111154
Chicago/Turabian StyleCao, Zi-Jian, Wen-Tao Qin, Juan Zhao, Yu Liu, Shou-Xian Wang, and Su-Yue Zheng. 2022. "Three New Trichoderma Species in Harzianum Clade Associated with the Contaminated Substrates of Edible Fungi" Journal of Fungi 8, no. 11: 1154. https://doi.org/10.3390/jof8111154