
New Insights into Lichenization in Agaricomycetes Based on an Unusual New Basidiolichen Species of Omphalina s. str.
 , and
, and 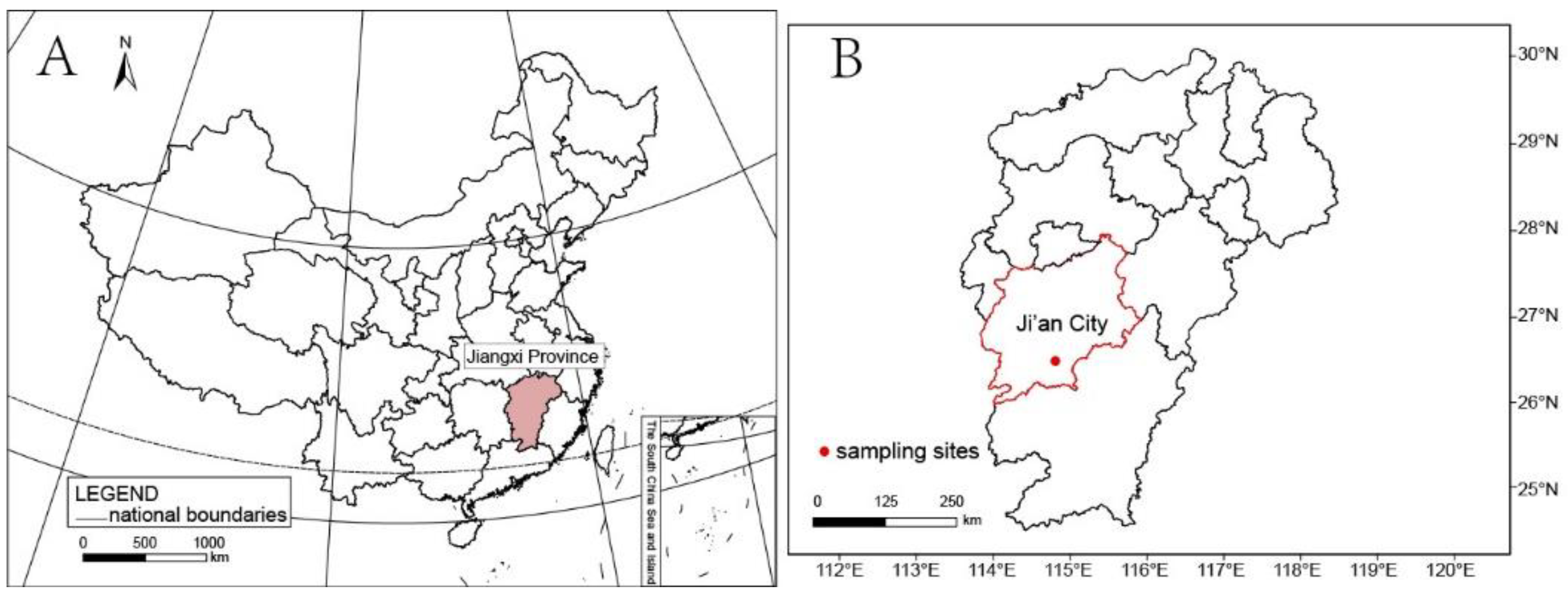{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Taxon Sampling and Morphological Examination
2.2. DNA Extraction, PCR, and Sequencing
2.3. Sequence Alignment and Phylogenetic Analysis
3. Results
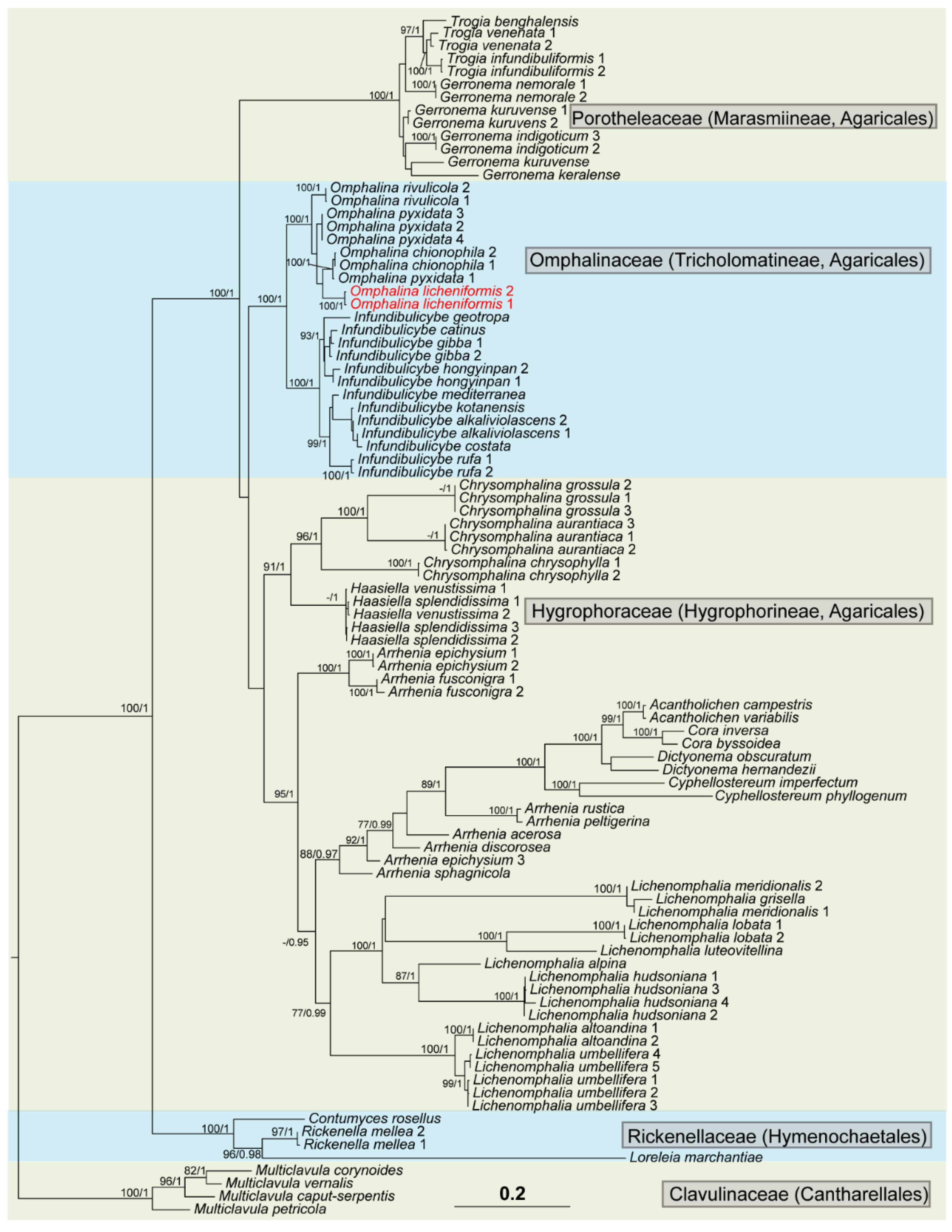
3.1. Phylogenetic Analysis
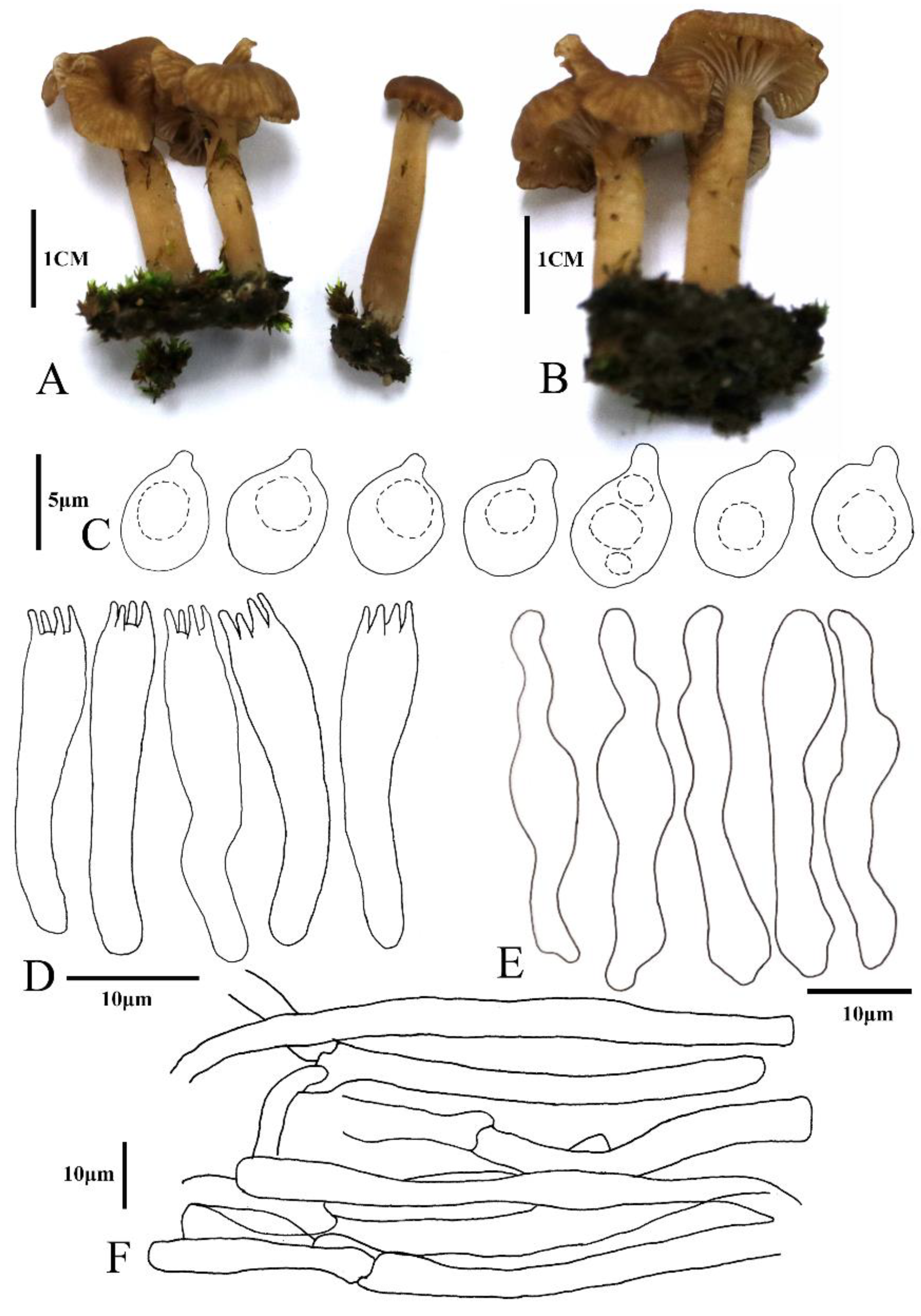
3.2. Taxonomy
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lücking, R.; Hodkinson, B.P.; Leavitt, S.D. The 2016 classification of lichenized fungi in the Ascomycota and Basidiomycota—Approaching one thousand genera. Bryologist 2017, 119, 361–416. [Google Scholar] [CrossRef]
- Redhead, S.; Lutzoni, F.; Moncalvo, J.M.; Vilgalys, R. Phylogeny of agarics: Partial systematics solutions for core Omphalinoid genera in the Agaricales (Euagarics). Mycotaxon 2002, 83, 19–57. [Google Scholar]
- Lücking, R.; Moncada, B. Dismantling Marchandiomphalina into Agonimia (Verrucariaceae) and Lawreymyces gen. nov. (Corticiaceae): Setting a precedent to the formal recognition of thousands of voucherless fungi based on type sequences. Fungal Divers. 2017, 84, 119–138. [Google Scholar] [CrossRef]
- Lutzoni, F.; Vilgalys, R. Omphalina (Basidiomycota, Agaricales) as a model system for the study of coevolution in lichens. Cryptogam. Bot. 1995, 5, 71–81. [Google Scholar]
- Lutzoni, F.M. Phylogeny of lichen- and non-lichen-forming omphalinoid mushrooms and the utility of testing for combinability among multiple data sets. Syst. Biol. 1997, 46, 373–406. [Google Scholar] [CrossRef] [PubMed]
- Bigelow, H.E. Omphalina in North America. Mycologia 1970, 62, 1–32. [Google Scholar] [CrossRef]
- Clémençon, H. Kompendium der Blätterpilze Europäische omphalinoide Tricholomataceae. Zeitschr. F. Mykol. 1982, 48, 195–237. [Google Scholar]
- Norvell, L.L.; Redhead, S.A.; Ammirati, J.F. Omphalina sensu lato in North America 1-2. 1: Omphalina wynniae and the genus Chrysomphalina. 2: Omphalina sensu Bigelow. Mycotaxon 1994, 50, 379–407. [Google Scholar]
- Bon, M. Flore mycologique d’Europe 4. Les Clitocybes, Omphales et ressemblants. Doc. Mycologia. Mém. Hors Sér 1997, 4, 1–181. [Google Scholar]
- Moncalvo, J.-M.; Vilgalys, R.; Redhead, S.; Johnson, J.; James, T.; Aime, M.; Valerie, H.; Verduin, S.; Larsson, E.; Baroni, T.; et al. One Hundred and Seventeen Clades of Eu-agarics. Mol. Phylogenet. Evol. 2002, 23, 357–400. [Google Scholar] [CrossRef]
- Matheny, P.; Curtis, J.; Valerie, H.; Aime, M.; Moncalvo, J.-M.; Ge, Z.-W.; Slot, J.; Ammirati, J.; Baroni, T.; Bougher, N.; et al. Major clades of Agaricales: A multilocus phylogenetic overview. Mycologia 2006, 98, 982–995. [Google Scholar] [CrossRef] [PubMed]
- Lawrey, J.; Lücking, R.; Sipman, H.; Chaves, J.; Redhead, S.; Bungartz, F.; Sikaroodi, M.; Gillevet, P. High concentration of basidiolichens in a single family of agaricoid mushrooms (Basidiomycota: Agaricales: Hygrophoraceae). Mycol. Res. 2009, 113, 1154–1171. [Google Scholar] [CrossRef] [PubMed]
- Lodge, D.; Padamsee, M.; Matheny, P.; Aime, M.; Cantrell, S.; Boertmann, D.; Kovalenko, A.; Vizzini, A.; Dentinger, B.; Kirk, P.; et al. Molecular phylogeny, morphology, pigment chemistry and ecology in Hygrophoraceae (Agaricales). Fungal Divers. 2013, 64, 1–99. [Google Scholar] [CrossRef]
- Varga, T.; Krizsan, K.; Földi, C.; Dima, B.; Sanchez-Garcia, M.; Sanchez-Ramirez, S.; Szollosi, G.; Szarkándi, J.; Papp, V.; Albert, L.; et al. Megaphylogeny resolves global patterns of mushroom evolution. Nat. Ecol. Evol. 2019, 3, 668–678. [Google Scholar] [CrossRef]
- Redhead, S.; Moncalvo, J.-M.; Vilgalys, R.; Lutzoni, F. Phylogeny of agarics: Partial systematics solutions for bryophilous omphalinoid agarics outside of the Agaricales (Euagarics). Mycotaxon 2002, 82, 151–168. [Google Scholar]
- Larsson, K.-H.; Parmasto, E.; Fischer, M.; Langer, E.; Nakasone, K.; Redhead, S. Hymenochaetales: A molecular phylogeny for the hymenochaetoid clade. Mycologia 2006, 98, 926–936. [Google Scholar] [CrossRef]
- Korotkin, H.; Swenie, R.; Miettinen, O.; Budke, J.; Chen, K.-H.; Lutzoni, F.; Smith, M.; Matheny, P. Stable isotope analyses reveal previously unknown trophic mode diversity in the Hymenochaetales. Am. J. Bot. 2018, 105, 1869–1887. [Google Scholar] [CrossRef]
- Olariaga, I.; Huhtinen, S.; Læssøe, T.; Petersen, J.; Hansen, K. Phylogenetic origins and family classification of typhuloid fungi, with emphasis on Ceratellopsis, Macrotyphula and Typhula (Basidiomycota). Stud. Mycol. 2020, 96, 155–184. [Google Scholar] [CrossRef]
- Vizzini, A.; Consiglio, G.; Marchetti, M.; Borovička, J.; Campo, E.; Cooper, J.; Lebeuf, R.; Ševčíková, H. New data in Porotheleaceae and Cyphellaceae: Epitypification of Prunulus scabripes Murrill, the status of Mycopan Redhead, Moncalvo & Vilgalys and a new combination in Pleurella Horak emend. Mycol. Prog. 2022, 21, 44. [Google Scholar] [CrossRef]
- Vizzini, A.; Picillo, B.; Luigi, P.; Dovana, F. Chrysomycena perplexa gen. et sp. nov. (Agaricales, Porotheleaceae), a new entity from the Lazio region (Italy). Riv. Micol. Romana 2019, 107, 96–107. [Google Scholar]
- Sanchez-Garcia, M.; Matheny, P. Is the switch to an ectomycorrhizal state an evolutionary key innovation in mushroom-forming fungi? A case study in the Tricholomatineae (Agaricales). Evolution 2016, 71, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Vizzini, A. Segnalazioni di muscinputa laevis (Basidiomycota, Agaricomycetes) per il Nord Italia. Micol. Veg. Mediterr. 2010, 25, 141–148. [Google Scholar]
- Redhead, S.; Norvell, L.L. Notes on Bondarzewia, Heterobasidion and Pleurogala, new genus. Mycotaxon 1993, 48, 371–380. [Google Scholar]
- Moncalvo, J.-M.; Lutzoni, F.; Rehner, S.; Johnson, J.; Vilgalys, R. Phylogenetic Relationships of Agaric Fungi Based on Nuclear Large Subunit Ribosomal DNA Sequences. Syst. Biol. 2000, 49, 278–305. [Google Scholar] [CrossRef]
- Lamoure, D. Agaricales de la zone alpine. Genre Omphalina (1ère partie). Trav. Sci. Parc. Natl. Vanoise 1974, 5, 149–164. [Google Scholar]
- Lamoure, D. Alpine and circumpolar Omphalina species. In Arctic and Alpine Mycology 1; Laursen, G.A., Ammirati, J.F., Eds.; University of Washington Press: Seattle, WA, USA, 1982; pp. 201–215. [Google Scholar]
- Bigelow, H.E. The Clitocybe pyxidata group. In Travaux Mycologiques Dédiés à R. Kühner, Numéro spécial du Bulletin de la Société Linnéenne de Lyon; Société Linnéenne de Lyon: Lyon, France, 1974; pp. 39–46. [Google Scholar]
- Bigelow, H.E. North American species of Clitocybe. Part I. Beih. Nova Hedwigia 1982, 72, 1–280. [Google Scholar]
- Bigelow, H.E. North American species of Clitocybe. Part II. Beih. Nova Hedwigia 1985, 81, 281–471. [Google Scholar]
- Sanchez-Garcia, M.; Ryberg, M.; Khan, F.; Varga, T.G.; Nagy, L.; Hibbett, D. Fruiting body form, not nutritional mode, is the major driver of diversification in mushroom-forming fungi. Proc. Natl. Acad. Sci. USA 2020, 117, 32528–32534. [Google Scholar] [CrossRef]
- Vizzini, A.; Para, R.; Fontenla, R.; Ghignone, S.; Ercole, E. A preliminary ITS phylogeny of Melanoleuca (Agaricales), with special reference to European taxa. Mycotaxon 2012, 118, 361–381. [Google Scholar] [CrossRef]
- Vizzini, A.; Marco, C.; Musumeci, E.; Ercole, E. A new taxon in the Infundibulicybe gibba complex (Basidiomycota, Agaricales, Tricholomataceae) from Sardinia (Italy). Mycologia 2010, 103, 203–208. [Google Scholar] [CrossRef]
- Vizzini, A.; Consiglio, G.; Marchetti, M.; Alvarado, P. Insights into the Tricholomatineae (Agaricales, Agaricomycetes): A new arrangement of Biannulariaceae and Callistosporium, Callistosporiaceae fam. nov., Xerophorus stat. nov., and Pleurocollybia incorporated into Callistosporium. Fungal Divers. 2020, 101, 211–259. [Google Scholar] [CrossRef]
- Liu, D.; Goffinet, B.; Wang, X.Y.; Hur, J.S.; Shi, H.X.; Zhang, Y.Y.; Yang, M.X.; LI, l.J.; Yin, A.C.; Wang, L.S. Another lineage of basidiolichen in China, the genera Dictyonema and Lichenomphalia (Agaricales: Hygrophoraceae). Mycosystema 2018, 37, 849–864. [Google Scholar] [CrossRef]
- Liu, D.; Goffinet, B.; Ertz, D.; De Kesel, A.; Wang, X.; Hur, J.S.; Shi, H.; Zhang, Y.; Yang, M.; Wang, L. Circumscription and phylogeny of the Lepidostromatales (lichenized Basidiomycota) following discovery of new species from China and Africa. Mycologia 2017, 109, 730–748. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.C. The Enumeration of Lichenized Fungi in China; China Forestry Publishing House: Beijing, China, 2020; pp. 1–606. [Google Scholar]
- Largent, D.L. How to Identify Mushrooms to Genus; Mad River Press: Eureka, CA, USA, 1986; Volume 1–5. [Google Scholar]
- Rogers, S.O.; Bendich, A.J. Extraction of DNA from plant tissues. In Plant Molecular Biology Manual A6; Gelvin, S.B., Schilperoort, R.A., Eds.; Kluwer Academic Publishers: Boston, MA, USA, 1988; pp. 1–10. [Google Scholar]
- White, T.; Bruns, T.; Lee, S.; Taylor, J.; Innis, M.; Gelfand, D.; Sninsky, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 31, 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- Yang, Q.; Cheng, X.; Zhang, T.; Liu, X.; Wei, X. Five new species of the lichen-forming fungal genus Peltula from China. J. Fungi 2022, 8, 134. [Google Scholar] [CrossRef]
- Swindell, S.R.; Plasterer, T.N. SEQMAN. In Sequence Data Analysis Guidebook; Swindell, S.R., Ed.; Humana Press: Totowa, NJ, USA, 1997; pp. 75–89. [Google Scholar]
- Katoh, K.; Asimenos, G.; Toh, H. Multiple alignment of DNA sequences with MAFFT. Methods Mol. Biol. 2009, 537, 39–64. [Google Scholar] [CrossRef] [PubMed]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef]
- Swofford, D. PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods); Version 4.0b10; Sinauer Associates, Inc.: Sunderland, UK.
- Zhang, T.; Zhang, X.; Yang, Q.; Wei, X. Hidden species diversity was explored in two genera of catapyrenioid lichens (Verrucariaceae, Ascomycota) from the deserts of China. J. Fungi 2022, 8, 729. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML Version 8: A Tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Vizzini, A.; Curti, M.; Contu, M.; Ercole, E. A new cystidiate variety of Omphalina pyxidata (Basidiomycota, tricholomatoid clade) from Italy. Mycotaxon 2012, 120, 361–371. [Google Scholar] [CrossRef]
- Vesterholt, J. Funga Nordica. Agaricoid, boletoid, cyphelloid and gasteroid genera. In Funga Nordica; Knudsen, H., Vesterholt, J., Eds.; Nordsvamp: Copenhagen, Denmark, 2012; pp. 485–487. [Google Scholar]
- Kutschera, U.; Niklas, K.J. Endosymbiosis, cell evolution, and speciation. Theory Biosci. 2005, 124, 1–24. [Google Scholar] [CrossRef]
- Masumoto, H.; Ohmura, Y.; Degawa, Y. Lichenomphalia meridionalis (Hygrophoraceae, lichenized Basidiomycota) new to Asia. Opusc. Philolichenum 2019, 18, 379–389. [Google Scholar]
- Voitk, A.; Saar, I.; Moncada, B.; Lickey, E. Circumscription and typification of sphagnicolous omphalinoid species of Arrhenia (Hygrophoraceae) in Newfoundland and Labrador: Three obligate and one facultative species. Mycol. Prog. 2022, 21, 57. [Google Scholar] [CrossRef]
- Oberwinkler, F. Basidiolichens. In Fungal Associations; Hock, B., Ed.; Springer: Berlin/Heidelberg, Germany, 2001; pp. 211–225. [Google Scholar]
- Gulden, G.; Senn-Irlet, B.; Jenssen, K.M.; Stordal, J. Arctic and Alpine Fungi Volume 3: Agarics of the Swiss Alps; Soppkonsulenten: Drammen, Norway, 1990; pp. 1–58. [Google Scholar]
- Courtecuisse, R.; Duhem, B. Guide des Champignons de France et d’Europe; Delachaux et NiestIe: Paris, France, 1994; pp. 1–544. [Google Scholar]
- Courtecuisse, R.; Duhem, B. Les Champignons de France; Eclectis, La Maison d’édition de la CAMIF: Paris, France, 1994. [Google Scholar]
- Lutzoni, F.; Pagel, M.; Reeb, V. Major fungal lineages are derived from lichen symbiotic ancestors. Nature 2001, 411, 937–940. [Google Scholar] [CrossRef]
- Gargas, A.; DePriest, P.; Grube, M.; Tehler, A. Multiple origins of lichen symbioses in fungi suggested by SSU rDNA phylogeny. Science 1995, 268, 1492–1495. [Google Scholar] [CrossRef] [PubMed]
- Honegger, R. Functional aspects of the lichen symbiosis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1991, 42, 553–578. [Google Scholar] [CrossRef]
- Tschermak-Woess, E. The algal partner. In Handbook of Lichenology; Galum, M., Ed.; CRC Press: Boca Raton, FL, USA, 1988; pp. 39–92. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, T.; Zhu, X.; Vizzini, A.; Li, B.; Cao, Z.; Guo, W.; Qi, S.; Wei, X.; Zhao, R. New Insights into Lichenization in Agaricomycetes Based on an Unusual New Basidiolichen Species of Omphalina s. str. J. Fungi 2022, 8, 1033. https://doi.org/10.3390/jof8101033
Zhang T, Zhu X, Vizzini A, Li B, Cao Z, Guo W, Qi S, Wei X, Zhao R. New Insights into Lichenization in Agaricomycetes Based on an Unusual New Basidiolichen Species of Omphalina s. str. Journal of Fungi. 2022; 8(10):1033. https://doi.org/10.3390/jof8101033
Chicago/Turabian StyleZhang, Tingting, Xinyu Zhu, Alfredo Vizzini, Biting Li, Zhenghua Cao, Wenqing Guo, Sha Qi, Xinli Wei, and Ruilin Zhao. 2022. "New Insights into Lichenization in Agaricomycetes Based on an Unusual New Basidiolichen Species of Omphalina s. str." Journal of Fungi 8, no. 10: 1033. https://doi.org/10.3390/jof8101033