
Three New Species of Clonostachys (Hypocreales, Ascomycota) from China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Morphological Studies
2.2. DNA Extraction, PCR Amplification, and Sequencing
2.3. Sequence Alignment and Phylogenetic Analyses
3. Results
3.1. Phylogeny
3.2. Taxonomy
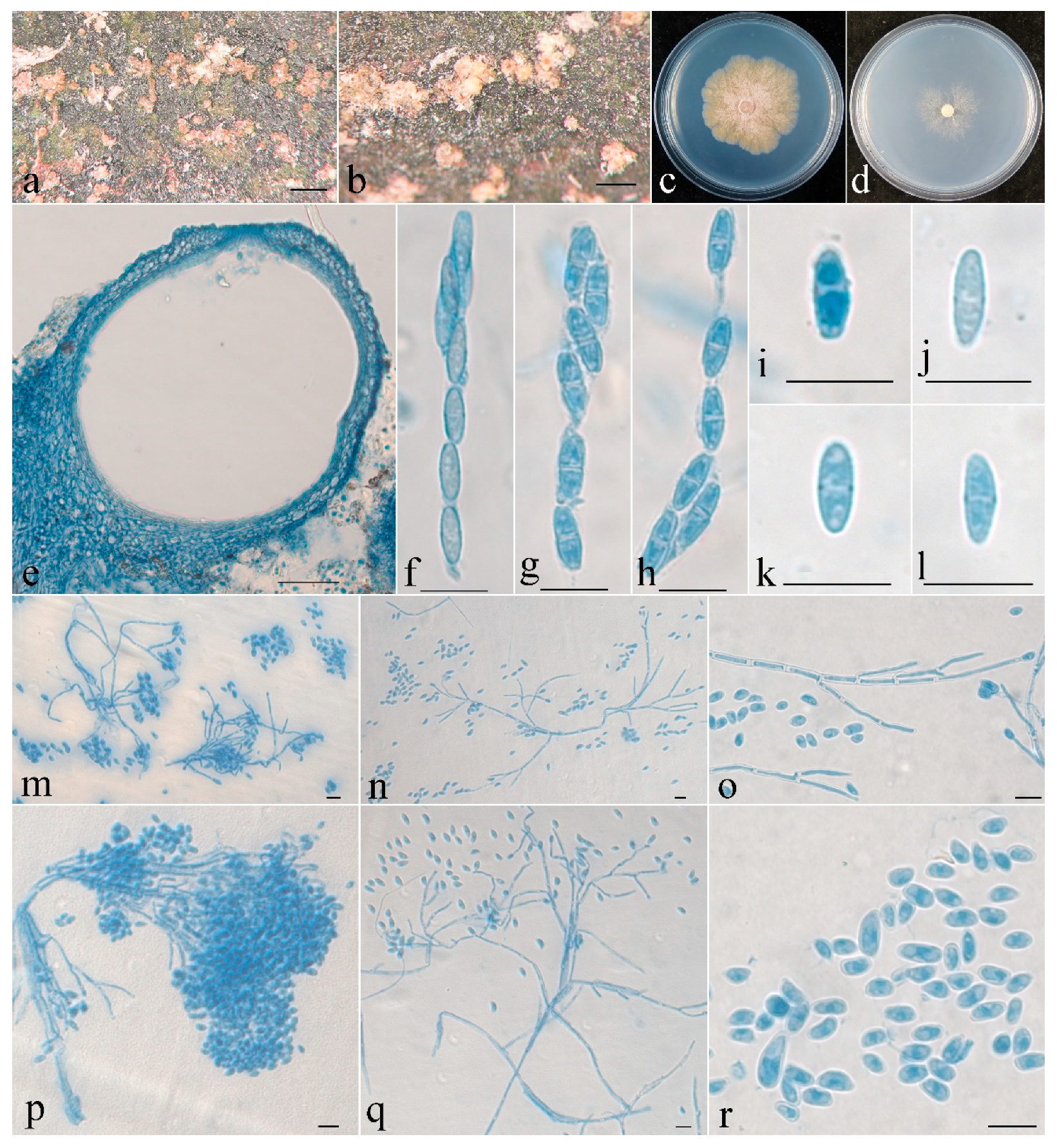
- Clonostachys chongqingensis Z.Q. Zeng and W.Y. Zhuang, sp. nov. (Figure 2).

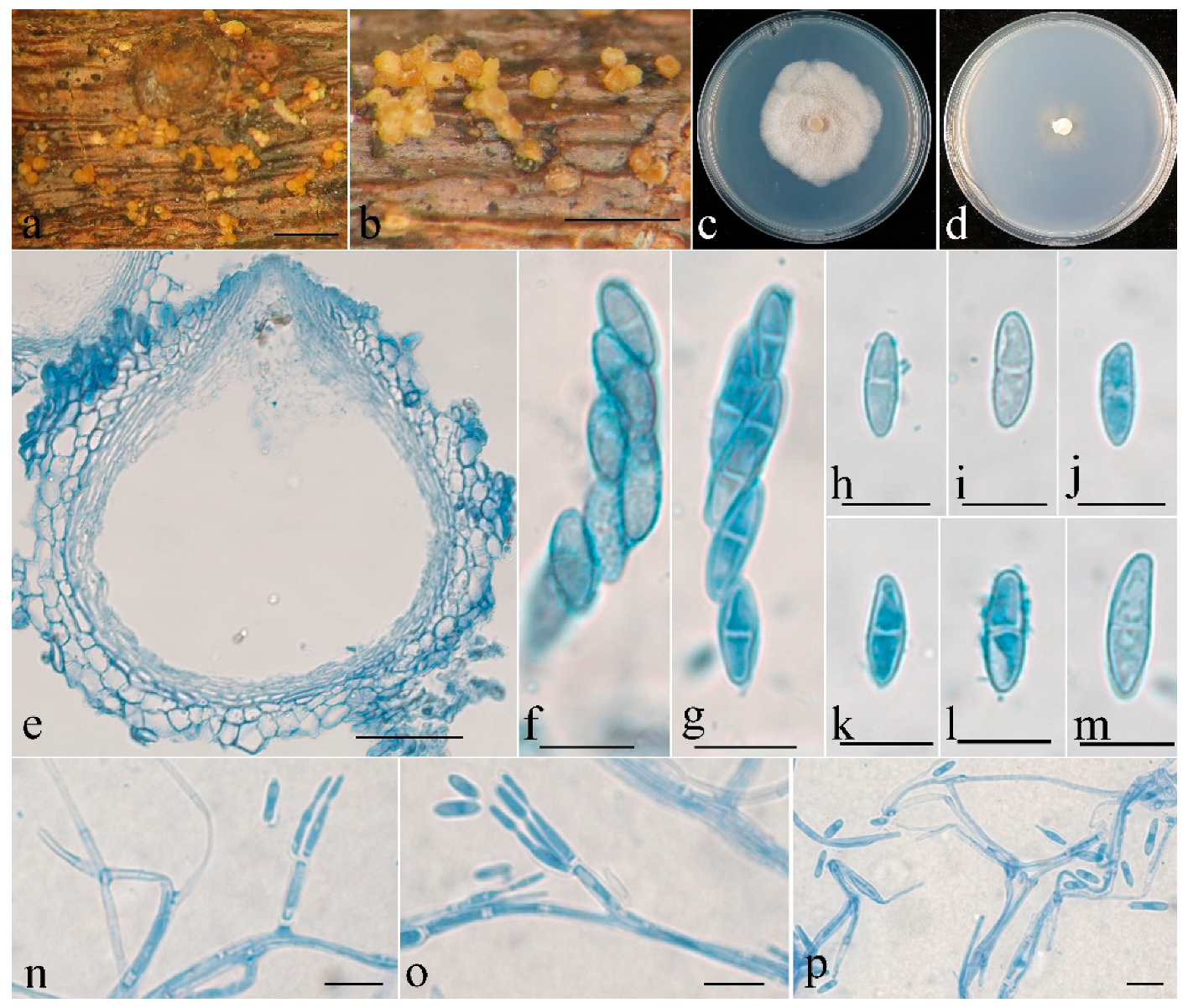
- Clonostachys leptoderma Z.Q. Zeng and W.Y. Zhuang, sp. nov. (Figure 3)

- Clonostachys oligospora Z.Q. Zeng and W.Y. Zhuang, sp. nov. (Figure 4)

4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schroers, H.J. A monograph of Bionectria (Ascomycota, Hypocreales, Bionectriaceae) and its Clonostachys anamorphs. Stud. Mycol. 2001, 46, 1–214. [Google Scholar]
- Chen, W.H.; Han, Y.F.; Liang, J.D.; Zou, X.; Liang, Z.Q.; Jin, D.C. A new araneogenous fungus of the genus Clonostachys. Mycosystema 2016, 35, 1061–1069. (In Chinese) [Google Scholar]
- Torcato, C.; Gonçalves, M.F.M.; Rodríguez-Gálvez, E.; Alves, A. Clonostachys viticola sp. nov., a novel species isolated from Vitis vinifera. Int. J. Syst. Evol. Microbiol. 2020, 70, 4321–4328. [Google Scholar] [CrossRef] [PubMed]
- Han, P.P.; Zhang, X.P.; Xu, D.; Zhang, B.W.; Lai, D.W.; Zhou, L.G. Metabolites from Clonostachys fungi and their biological activities. J. Fungi 2020, 6, 229. [Google Scholar]
- Zheng, C.J.; Kim, C.J.; Bae, K.S.; Kim, Y.H.; Kim, W.G. Bionectins A-C, epidithiodioxopiperazines with anti-MRSA activity, from Bionectria byssicola F120. J. Nat. Prod. 2006, 69, 1816–1819. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M.; Dubey, M.; McEwan, K.; Menzel, U.; Franko, M.A.; Viketoft, M.; Jensen, D.F.; Karlsson, M. Evaluation of Clonostachys rosea for control of plant-parasitic Nematodes in soil and in roots of carrot and wheat. Phytopathology 2018, 108, 52–59. [Google Scholar] [CrossRef]
- Zhuang, W.Y. (Ed.) Flora Fungorum Sinicorum. Vol. Nectriaceae et Bionectriaceae; Science Press: Beijing, China, 2013; p. 162. (In Chinese) [Google Scholar]
- Rossman, A.Y. Lessons learned from moving to one scientific name for fungi. IMA Fungus 2014, 5, 81–89. [Google Scholar] [CrossRef]
- Lombard, L.; Van der Merwe, N.A.; Groenewald, J.Z.; Crous, P.W. Generic concepts in Nectriaceae. Stud. Mycol. 2015, 80, 189–245. [Google Scholar] [CrossRef]
- Dao, H.T.; Beattie, G.A.C.; Rossman, A.Y.; Burgess, L.W.; Holford, P. Four putative entomopathogenic fungi of armoured scale insects on Citrus in Australia. Mycol. Prog. 2016, 15, 47. [Google Scholar] [CrossRef]
- Moreira, G.M.; Abreu, L.M.; Carvalho, V.G.; Schroers, H.J.; Pfenning, L.H. Multilocus phylogeny of Clonostachys subgenus Bionectria from Brazil and description of Clonostachys chloroleuca sp. nov. Mycol. Prog. 2016, 15, 1031–1039. [Google Scholar] [CrossRef]
- Prasher, I.B.; Chauhan, R. Clonostachys indicus sp. nov. from India. Kavaka 2017, 48, 22–26. [Google Scholar]
- Lechat, C.; Fournier, J. Clonostachys spinulosispora (Hypocreales, Bionectriaceae), a new species on palm from French Guiana. Ascomycete.org 2018, 10, 127–130. [Google Scholar]
- Lechat, C.; Fournier, J. Two new species of Clonostachys (Bionectriaceae, Hypocreales) from Saül (French Guiana). Ascomycete.org 2020, 12, 61–66. [Google Scholar]
- Tibpromma, S.; Hyde, K.D.; McKenzie, E.H.C.; Bhat, D.J.; Phillips, A.J.L.; Wanasinghe, D.N.; Samarakoon, M.C.; Jayawardena, R.S.; Dissanayake, A.J.; Tennakoon, D.S.; et al. Fungal diversity notes 840–928: Micro-fungi associated with Pandanaceae. Fungal Divers. 2018, 93, 1–160. [Google Scholar] [CrossRef]
- Lechat, C.; Fournier, J.; Chaduli, D.; Lesage-Meessen, L.; Favel, A. Clonostachys saulensis (Bionectriaceae, Hypocreales), a new species from French Guiana. Ascomycete.org 2019, 11, 65–68. [Google Scholar]
- Lechat, C.; Fournier, J.; Gasch, A. Clonostachys moreaui (Hypocreales, Bionectriaceae), a new species from the island of Madeira (Portugal). Ascomycete.org 2020, 12, 35–38. [Google Scholar]
- Forin, N.; Vizzini, A.; Nigris, S.; Ercole, E.; Voyron, S.; Girlanda, M.; Baldan, B. Illuminating type collections of nectriaceous fungi in Saccardo’s fungarium. Persoonia 2020, 45, 221–249. [Google Scholar] [CrossRef]
- Hyde, K.D.; Dong, Y.; Phookamsak, R.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Liu, N.; Abeywickrama, P.D.; Mapook, A.; Wei, D.; et al. Fungal diversity notes 1151–1276: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2020, 100, 5–277. [Google Scholar] [CrossRef] [Green Version]
- Perera, R.H.; Hyde, K.D.; Maharachchikumbura, S.S.N.; Jones, E.B.G.; McKenzie, E.H.C.; Stadler, M.; Lee, H.B.; Samarakoon, M.C.; Ekanayaka, A.H.; Camporesi, E.; et al. Fungi on wild seeds and fruits. Mycosphere 2020, 11, 2108–2480. [Google Scholar] [CrossRef]
- Zeng, Z.Q.; Zhuang, W.Y. Three new Chinese records of Hypocreales. Mycosystema 2017, 36, 654–662. [Google Scholar]
- Hirooka, Y.; Kobayashi, T.; Ono, T.; Rossman, A.Y.; Chaverri, P. Verrucostoma, a new genus in the Bionectriaceae from the Bonin islands, Japan. Mycologia 2010, 102, 418–429. [Google Scholar] [CrossRef] [PubMed]
- Nirenberg, H.I. Studies on the morphologic and biologic differentiation in Fusarium section Liseola. Mitt. Biol. Bundesanst. Land-Forstw. 1976, 169, 1–117. [Google Scholar]
- Wang, L.; Zhuang, W.Y. Designing primer sets for amplification of partial calmodulin genes from penicillia. Mycosystema 2004, 23, 466–473. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfland, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Gräfenhan, T.; Schroers, H.J.; Nirenberg, H.I.; Seifert, K.A. An overview of the taxonomy, phylogeny and typification of nectriaceous fungi in Cosmospora, Acremonium, Fusarium, Stilbella and Volutella. Stud. Mycol. 2011, 68, 79–113. [Google Scholar] [CrossRef] [PubMed]
- Castlebury, L.A.; Rossman, A.Y.; Sung, G.H.; Hyten, A.S.; Spatafora, J.W. Multigene phylogeny reveals new lineage for Stachybotrys chartarum, the indoor air fungus. Mycol. Res. 2004, 108, 864–872. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgin, D.G. The ClustalX windows interface: Flexible strategies for multiple sequences alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4883. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP 4.0b10: Phylogenetic Analysis Using Parsimony (* and Other Methods); Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Nylander, J.A.A. MrModeltest v2.0. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Chernomor, O.; von Haeseler, A.; Minh, B.Q. Terrace aware data structure for phylogenomic inference from supermatrices. Mol. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef] [PubMed]
- Page, R.D. TreeView: An application to display phylogenetic trees on personal computers. Comp. Appl. Biosci. 1996, 12, 357–358. [Google Scholar] [PubMed]
- Cunningham, C.W. Can three incongruence tests predict when data should be combined? Mol. Biol. Evol. 1997, 14, 733–740. [Google Scholar] [CrossRef]
- Spegazzini, C. Fungi costaricenses nonnulli. Bol. Acad. Nac. Cienc. Córdoba 1919, 23, 541–609. [Google Scholar]
- Rossman, A.Y.; Samuels, G.J.; Rogerson, C.T.; Lowen, R. Genera of Bionectriaceae, Hypocreaceae and Nectriaceae (Hypocreales, Ascomycetes). Stud. Mycol. 1999, 42, 1–248. [Google Scholar]
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.H.; Li, D.Z.; Marhold, K.; et al. International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code); Koeltz Scientific Books: Berlin, Germany, 2018; p. 254. [Google Scholar]
- Rossman, A.Y.; Seifert, K.A.; Samuel, G.J.; Minnis, A.M.; Schroers, H.J.; Lombard, L.; Crous, P.W.; Põldma, K.; Cannon, P.F.; Summerbel, R.C.; et al. Genera in Bionectriaceae, Hypocreaceae, and Nectriaceae (Hypocreales) proposed for acceptance or rejection. IMA Fungus 2013, 4, 41–51. [Google Scholar] [CrossRef]
- Zhu, S.; Ren, F.; Guo, Z.; Liu, J.; Liu, X.; Liu, G.; Che, Y. Rogersonins A and B, imidazolone N-oxide-incorporating indole alkaloids from a verG disruption mutant of Clonostachys rogersoniana. J. Nat. Prod. 2019, 82, 462–468. [Google Scholar] [CrossRef]
- Ouchi, T.; Watanabe, Y.; Nonaka, K.; Muramatsu, R.; Noguchi, C.; Tozawa, M.; Hokari, R.; Ishiyama, A.; Koike, R.; Matsui, H.; et al. Clonocoprogens A, B and C, new antimalarial coprogens from the Okinawan fungus Clonostachys compactiuscula FKR-0021. J. Antibiot. 2020, 73, 365–371. [Google Scholar] [CrossRef]
- Sun, Z.B.; Li, S.D.; Ren, Q.; Xu, J.L.; Lu, X.; Sun, M.H. Biology and applications of Clonostachys rosea. J. Appl. Microbiol. 2020, 129, 486–495. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Xiang, L.G.; Zhuang, W.Y.; Zeng, Z.Q.; Yu, Z.H. Screening of antagonistic Closnostachys strains against Phytophthora nicotianae. Microbiol. China 2022, 49, 3860–3872. [Google Scholar]
- Lee, S.A.; Kang, M.J.; Kim, T.D.; Park, E.J. First report of Clonostachys rosea causing root rot of Gastrodia elata in Korea. Plant Dis. 2020, 104, 3069. [Google Scholar] [CrossRef]
- Haque, M.E.; Parvin, M.S. First report of Clonostachys rosea causing root rot of Beta vulgaris in North Dakota, USA. New Dis. Rep. 2020, 42, 21. [Google Scholar] [CrossRef]
- Zeng, Z.Q.; Zhuang, W.Y. Current understanding of the genus Hypomyces (Hypocreales). Mycosystema 2015, 34, 809–816. [Google Scholar]
- Yu, Y.N.; Mao, X.L. (Eds.) 100 Years of Mycology in China; Science Press: Beijing, China, 2015; p. 763. (In Chinese) [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Herbarium/Strain Numbers | GenBank Accession Numbers | |
|---|---|---|---|
| ITS | BenA | ||
| C. agrawalii | CBS 53381 | AF358241 | AF358187 |
| C. apocyni | CBS 13087 | AF210688 | AF358168 |
| C. aranearum | QLS 0625 | NR164542 | KU212400 |
| C. aureofulvella | CBS 19593 | AF358226 | AF358181 |
| C. buxi | CBS 69693 | KM231840 | KM232111 |
| C. byssicola | CBS 36478 | MH861151 | AF358153 |
| C. candelabrum | CBS 50467/CML 2313 | MH859044 | KF871186 |
| C. capitata | CBS 21893 | AF358240 | AF358188 |
| C. chlorina | CBS 28790 | NR137651 | - |
| C. chloroleuca | CML 1941 | KC806286 | KF871172 |
| C.chongqingensis | HMAS 290894 | OP205475 a | OP205324 |
| C. coccicola | HD 2016 | KU720552 | KU720552 |
| C. compactiuscula | CBS 72987 | AF358242 | AF358193 |
| C. divergens | CBS 96773b | NR137532 | AF358191 |
| C. epichloe | CBS 101037 | AF210675 | AF358209 |
| C. eriocamporesiana | MFLUCC 17-2620 | NR168235 | MN699965 |
| C. eriocamporesii | MFLUCC 190486 | NR168236 | - |
| C. grammicospora | CBS 20993 | NR137650 | AF358206 |
| C. grammicosporopsis | CBS 11587 | AF210679 | AF358204 |
| C. impariphialis | HMAS 275560 | KX096609 | - |
| C. indica | RKV 2015 | KT291441 | KT291441 |
| C. intermedia | CBS 50882 | NR137652 | AF358205 |
| C. kowhai | CBS 46195 | AF358250 | AF358170 |
| C. krabiensis | MFLUCC 160254 | NR168189 | - |
| C. leptoderma | HMAS 255834 | OP205474 | OP205323 |
| C. levigata | CBS 94897 | AF210680 | AF358196 |
| C. lucifer | CBS 100008 | AF210683 | AF358208 |
| C. miodochialis | CBS 99769 | NR137649 | AF358210 |
| C. oblongispora | CBS 100285 | AF358248 | AF358169 |
| C.oligospora | HMAS 290895 | OP205473 | OP205322 |
| C. phyllophila | CBS 92197 | NR137531 | - |
| C. pityrodes | CBS 102033 | AF210672 | AF358212 |
| C. pseudochroleuca | CML 2513 | KJ499909 | KF871188 |
| C. pseudostriata | CBS 12087 | MH862056 | AF358184 |
| C. ralfsii | CBS 102845 | AF358253 | AF358219 |
| C. rhizophaga | CBS 36177 | AF358228 | AF358158 |
| C. rogersoniana | CBS 58289/CML 2557 | AF210691 | KX185047 |
| C. rosea | CBS 71086 | MH862010 | AF358161 |
| C. rossmaniae | CBS 21093 | AF358227 | AF358213 |
| C. samuelsii | CBS 69997 | AF358236 | AF358190 |
| C. saulensis | BRFM 2782 | MK635054 | - |
| C. sesquicillii | CBS 18088 | AF210666 | AF358214 |
| C. setosa | CBS 83491 | AF210670 | AF358211 |
| C. solani | CBS 101926 | AF358230 | AF358179 |
| C. sporondochialis | CBS 101921 | AF210685 | AF358149 |
| C. subquaternata | CBS 100003/CBS 10787 | MT537603 | AF358207 |
| C. vesiculosa | HMAS 183151 | NR119828 | - |
| C. viticola | CAA 944 | MK156282 | MK156290 |
| C. wenpingii | HMAS 172156 | EF612465 | HM054127 |
| C. zelandiaenovae | CBS 23280 | AF210684 | AF358185 |
| F. acutatum | CBS 40297 | NR111142 | KU603870 |
| N. cinnabarina | CBS 18987 | HM484699 | HM484835 |
| Species | Sexual Morph | Asexual Morph | Reference Source | ||||
|---|---|---|---|---|---|---|---|
| Perithecia | Asci | Ascospores | Conidiophores | Phialides | Conidia | ||
| C. chongqingensis | Subglobose to globose, 304–353 × 294–392 μm. | Clavate to subcylindrical, 60–85 × 6–13 μm. | Ellipsoidal to elongate-ellipsoidal, spinulose, 13–16 × 4.5–5.5 μm. | Acremonium- to verticillium-like. | Subulate, 15–74 × 1.6–2.5 μm. | Ellipsoidal to rod-shaped, 4–10 × 2.5–4 μm. | This study |
| C. krabiensis | N.A. | N.A. | N.A. | Aggregated into sporodochia. | Subulate, 10–13 × 1.5–2.5 μm. | Cylindrical to oblong, 5–7 × 1–2 μm. | [15] |
| C. sesquicillii | Globose, 250–300 μm diam. | Narrowly clavate to cylindrical, 35–63 × 5–13 μm. | Ellipsoidal, warted, 8.2–14.4 × 2.2–4.4 μm. | Penicillate conidiophores bi- to quarter-verticillate, verticillate conidiophores sparsely formed. | Cylindrical to flask-shaped, 6.4–18.8 × 1.4–3.6 μm. | Ellipsoidal to oblong ellipsoidal, 4.2–9.6 × 1.6–3 μm. | [1] |
| C. viticola | N.A. | N.A. | N.A. | Primary conidiophores verticillate, secondary conidiophores bi- to ter-verticillate, 45.3–64.7 × 2.1–3.7 μm. | Cylindrical, 10.4–32.8 × 1.7–2.7 μm. | Ellipsoidal to oval, 4.5–6.7 × 2.4–3.4 μm. | [3] |
| C. zelandiaenovae | Subglobose to globose, 290–550 μm diam. | Narrowly clavate, 60–104 × 7–15.5 μm. | Ellipsoidal, spinulose, rarely smooth, 11.6–21.4 × 3.8–7.4 μm. | Primary conidiophores verticillate, secondary conidiophores penicillate, ter- to quinquies-verticillate. | Cylindrical to narrowly flask-shaped, 4.8–20.6 × 1.6–3.4 μm. | Distally broadly rounded, 4–13.2 × 2.4–4.2 μm. | [1] |
| C. leptoderma | Subglobose to globose, 216–284 × 206–265 μm. | Clavate to subcylindrical, 53–63 × 4.8–7 μm. | Ellipsoidal to elongate-ellipsoidal, spinulose, 7.5–11 × 2.5–3.5 μm. | Verticillium-like. | Subulate, 9–18 × 1.5–2.5 μm. | Subglobose, ellipsoidal to subellipsoidal, 2–7 × 2–5 μm. | This study |
| C. agrawalii | N.A. | N.A. | N.A. | Primary conidiophores irregularly branched to ter-verticillate, secondary conidiophores bi- to quarter-verticillate. | Cylindrical to flask-shaped, 7–42 × 1.4–3.4 μm. | Ends broadly rounded, 3.8–5.8 × 2.2–3 μm. | [1] |
| C. capitata | Subglobose to oval, around 300 μm diam. | Narrowly clavate, 50.5–89.5 × 7–12 μm. | Ellipsoidal to oblong-ellipsoidal, spinulose to warted, 11.6–18.8 × 3.6–5.8 μm. | Primary conidiophores verticillium-like, secondary conidiophores ter- to quinquies-verticillium-like. | Cylindrical to narrowly flask-shaped, 8.8–46.6 × 1.4–3.6 μm. | Ends broadly rounded, 4.6–12.4 × 2.2–4.2 μm. | [1] |
| C. epichloe | 140–240 × 140–200 μm. | Clavate to narrowly clavate, 32–65 × 5–10 μm. | Ellipsoidal, smooth to spinulose, 7.2–13 × 2.4–4.4 μm. | Divergently branched to adpressed. | Cylindrical to narrowly flask-shaped, 7–29 × 2.2–3.2 μm. | Ellipsoidal to narrowly clavate, 4.8–9.6 × 1.6–3.6 μm. | [1] |
| C. oligospora | Subglobose to globose, 225–274 × 225–265 μm. | Clavate, 45–65 × 7.5–11 μm. | Ellipsoidal, smooth, 9–17 × 3–5.5 μm. | Verticillium-like. | Subulate, 9–15 × 1.5–2.5 μm. | Sparse, subfusiform, 5–13 × 1.8–2.2 μm. | This study |
| C. indica | N.A. | N.A. | N.A. | Primary conidiophores verticillium-like, secondary conidiophores bi- to quarter-verticillate. | Cylindrical, 10.5–35.9 × 1.9–3.9 μm. | Ovoid to subglobose, 3.9–7.4 × 2–3.7 μm. | [12] |
| C. samuelsii | Subglobose to oval, 250–350 μm diam. | Narrowly clavate, 43–71 × 5–11.5 μm. | Ellipsoidal to broadly ellipsoidal, warted, 7.8–15.4 × 2.8–5.6 μm. | Penicillate or irregularly penicillate, ter- to quarter-verticillate. | Phialides cylindrical to narrowly flask-shaped, 3.8–20.6 × 1.8–5 μm. | Conidia ellipsoidal, 4.4–11.6 × 2.2–3.8 μm. | [1] |
| C. setosa | Globose, around 200 μm diam. | Narrowly clavate, 45–53 × 6.5–9 μm. | Ellipsoidal, smooth to striate, 8.8–13 × 2.4–3.8 μm. | Penicillate, mono- to ter-verticillate. | Cylindrical, 5.4–13.4 × 2.4–4.2 μm. | Cylindrical, 8.6–19.2 × 2–3.2 μm. | [1] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, Z.-Q.; Zhuang, W.-Y. Three New Species of Clonostachys (Hypocreales, Ascomycota) from China. J. Fungi 2022, 8, 1027. https://doi.org/10.3390/jof8101027
Zeng Z-Q, Zhuang W-Y. Three New Species of Clonostachys (Hypocreales, Ascomycota) from China. Journal of Fungi. 2022; 8(10):1027. https://doi.org/10.3390/jof8101027
Chicago/Turabian StyleZeng, Zhao-Qing, and Wen-Ying Zhuang. 2022. "Three New Species of Clonostachys (Hypocreales, Ascomycota) from China" Journal of Fungi 8, no. 10: 1027. https://doi.org/10.3390/jof8101027