

Forest Type and Site Conditions Influence the Diversity and Biomass of Edible Macrofungal Species in Ethiopia
 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
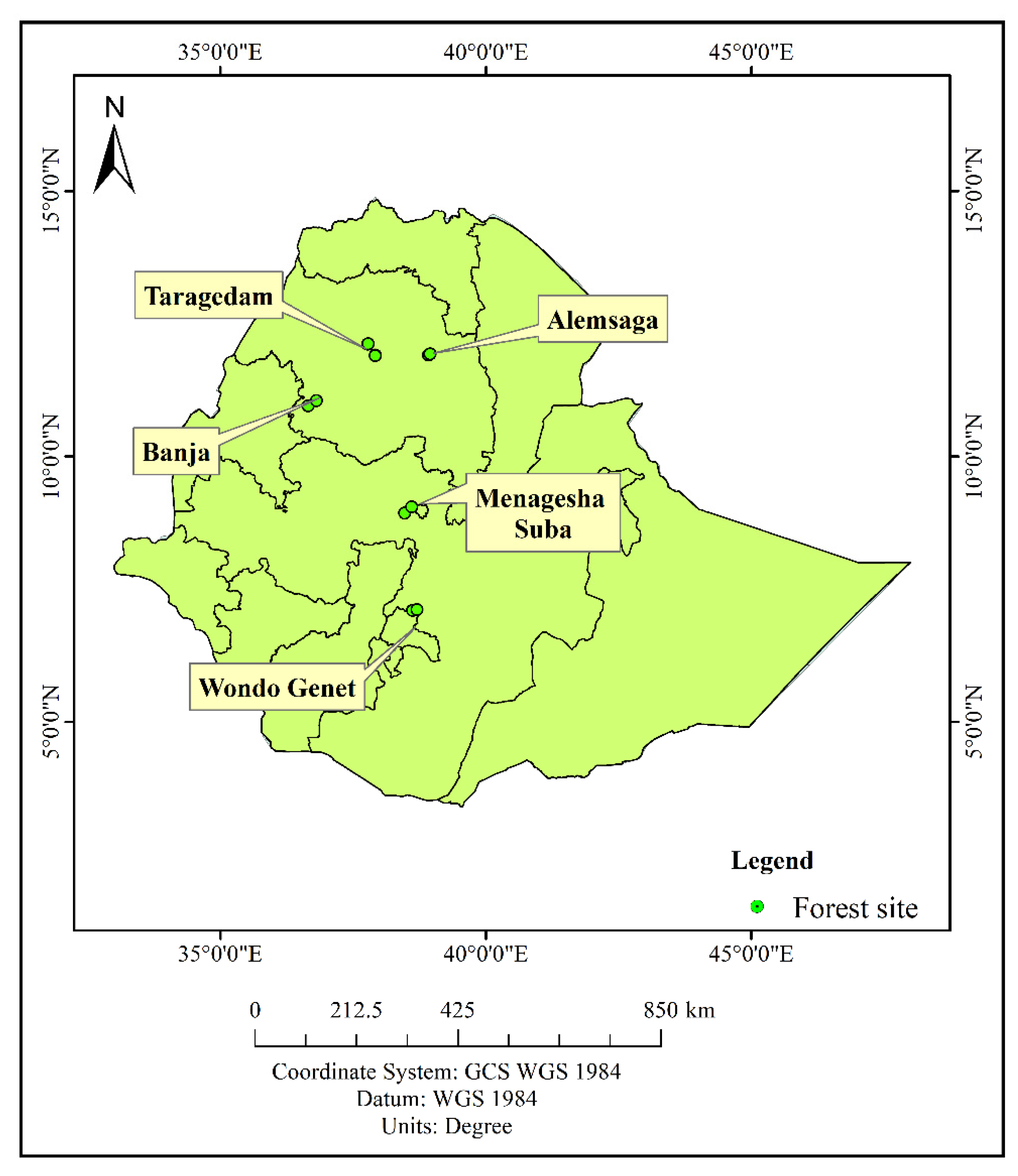
2.1. Study Sites
2.2. Experimental Design and Sporocarp Sampling
2.3. Species Identification and Characterization
2.4. Environmental Data Collection and Analysis
2.5. Data Analysis
3. Results
3.1. Edible Fungal Richness
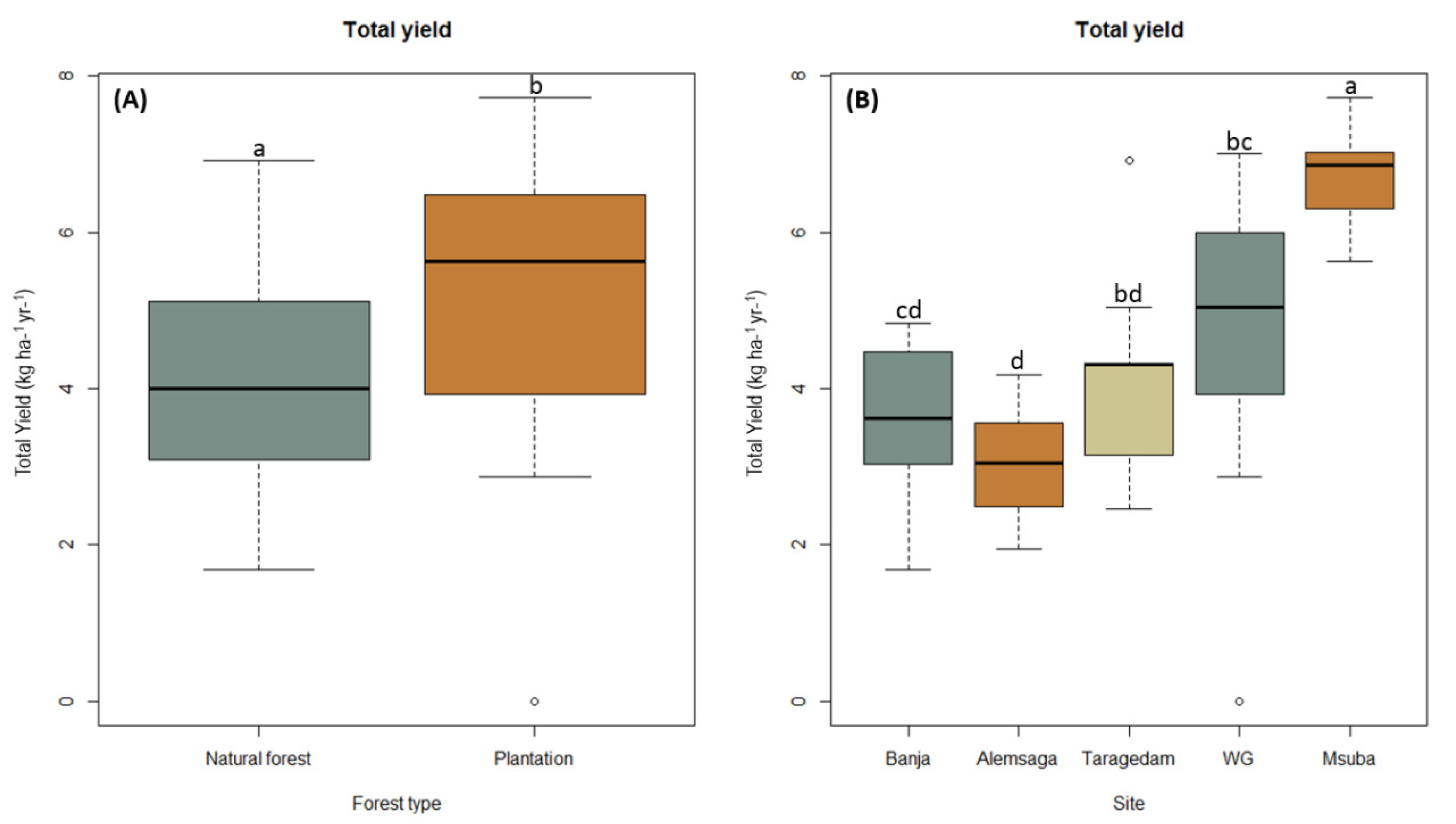
3.2. Sporocarp Production
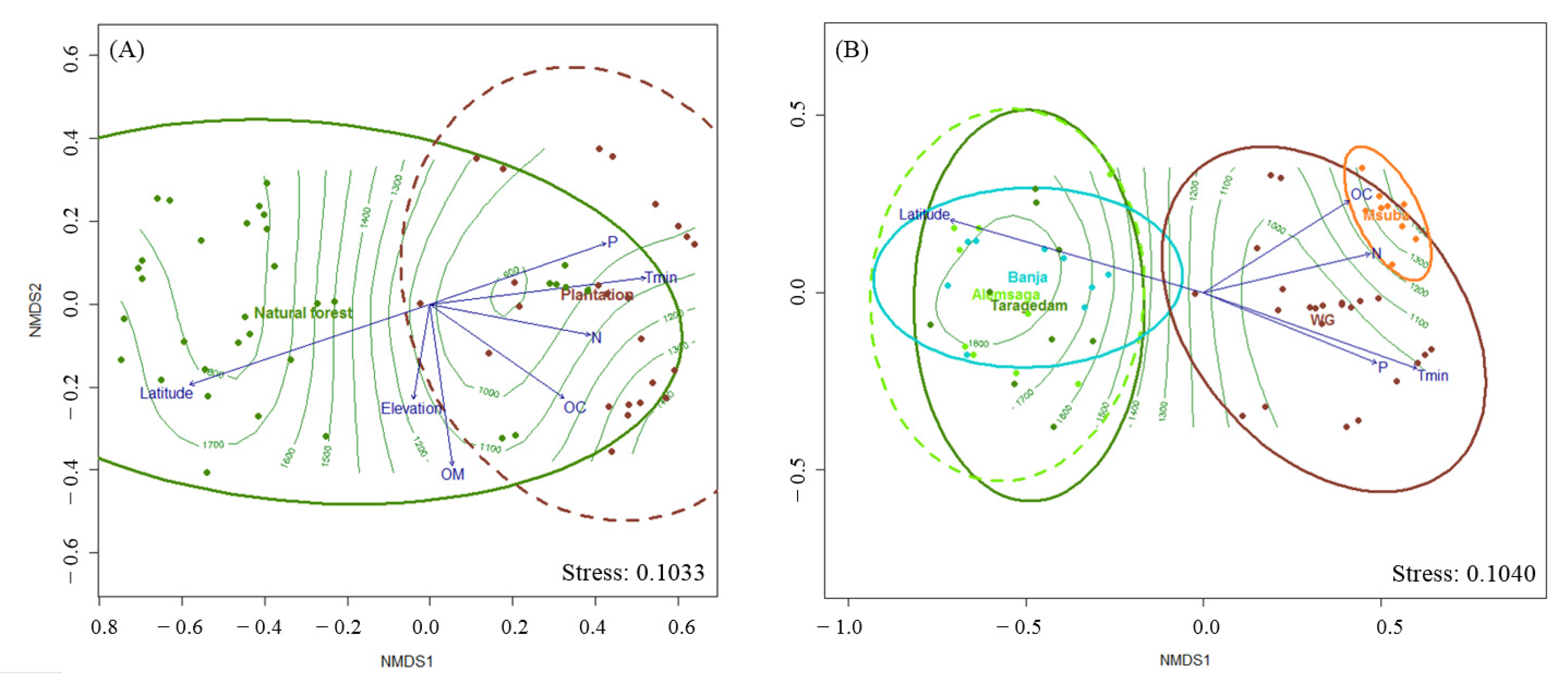
3.3. Sporocarp Composition and Environmental Variables
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Friis, B.; Demissew, S.; Breugel, P. Atlas of the Potential Vegetation of Ethiopia; The Royal Danish Academy of Sciences and Letters: Copenhagen, Denmark, 2010. [Google Scholar]
- Badege, B. Deforestation and Land Degradation in the Ethiopian Highlands: A Strategy for Physical Recovery. Northeast Afr. Stud. 2001, 8, 7–26. [Google Scholar]
- Bekele, M. Forest Plantations and Woodlots in Ethiopia. Afr. For. forum 2011, 1, 52. [Google Scholar]
- Adilo, M. The Contribution of Non-Timber Forest Products to Rural Livelihood in Southwest Ethiopia. Master’s Thesis, Wageningen University, Wageningen, The Netherlands, 2007. [Google Scholar]
- Seyoum, A. Economic value of Afromontane Forests of Sheka: Forest of Sheka. In Forest of Sheka; Fetene, M., Ed.; MEHBER: Addis Ababa, Ethiopia, 2007; pp. 183–218. [Google Scholar]
- Sultan, M. The Role of Non-Timber Forest Products to Rural Livelihoods and Forest Conservation: A Case Study at Harana Bulluk District Oromia National Regional State, Ethiopia. Master’s Thesis, Wondo Genet College of Forestry and Natural Resource, Hawassa University, Wondo Genet, Ethiopia, 2009. [Google Scholar]
- Lulekal, E.; Asfaw, Z.; Kelbessa, E.; Van Damme, P. Wild edible plants in Ethiopia: A review on their potential to combat food insecurity. Afrika Focus 2011, 24, 71–122. [Google Scholar] [CrossRef]
- Melaku, E.; Ewnetu, Z.; Teketay, D. Non-timber forest products and household incomes in Bonga forest area, southwestern Ethiopia. J. For. Res. 2014, 25, 215–223. [Google Scholar] [CrossRef]
- Asfaw, Z.; Tadesse, M. Prospects for sustainable use and development of wild food plants in Ethiopia. Econ. Bot. 2001, 55, 47–62. [Google Scholar] [CrossRef]
- Sitotaw, R.; Lulekal, E.; Abate, D. Ethnomycological study of edible and medicinal mushrooms in Menge District, Asossa Zone, Benshangul Gumuz Region, Ethiopia. J. Ethnobiol. Ethnomed. 2020, 16, 11. [Google Scholar] [CrossRef]
- Dejene, T.; Oria-de-Rueda, J.A.; Martín-Pinto, P. Edible wild mushrooms of Ethiopia: Neglected non-timber forest products. Rev. Fitotec. Mex. 2017, 40, 391–397. [Google Scholar] [CrossRef]
- Boa, E. Wild Edible Fungi: A Global Overview of Their Use and Importance to People; FAO: Rome, Italy, 2004; ISBN 9251051577. [Google Scholar]
- Barroetaveña, C.; La Manna, L.; Alonso, M.V. Variables affecting Suillus luteus fructification in ponderosa pine plantations of Patagonia (Argentina). For. Ecol. Manag. 2008, 256, 1868–1874. [Google Scholar] [CrossRef]
- De Román, M.; Boa, E. International Seminar on Ethnomycology Collection, Marketing and Cultivation of Edible Fungi in Spain. Micol. Apl. Int. 2004, 16, 25–33. [Google Scholar]
- Chang, Y.S.; Lee, S.S. Utilisation of macrofungi species in Malaysia. Fungal Divers 2004, 15, 15–22. [Google Scholar]
- Mau, J.; Chang, C.; Huang, S.; Chen, C. Antioxidant properties of methanolic extracts from Grifola frondosa, Morchella esculenta and Termitomyces albuminosus mycelia. Food Chem. 2004, 87, 111–118. [Google Scholar] [CrossRef]
- Chen, Y.; Yuan, Z.; Bi, S.; Wang, X.; Ye, Y.; Svenning, J.-C. Macrofungal species distributions depend on habitat partitioning of topography, light, and vegetation in a temperate mountain forest. Sci. Rep. 2018, 8, 13589. [Google Scholar] [CrossRef]
- Brundrett, M.C. Mycorrhizal associations and other means of nutrition of vascular plants: Understanding the global diversity of host plants by resolving conflicting information and developing reliable means of diagnosis. Plant Soil 2009, 320, 37–77. [Google Scholar] [CrossRef]
- Courty, P.-E.; Buée, M.; Diedhiou, A.G.; Frey-Klett, P.; Le Tacon, F.; Rineau, F.; Turpault, M.-P.; Uroz, S.; Garbaye, J. The role of ectomycorrhizal communities in forest ecosystem processes: New perspectives and emerging concepts. Soil Biol. Biochem. 2010, 42, 679–698. [Google Scholar] [CrossRef]
- Fernández-Toirán, L.; Agreda, T.; Olano, J. Stand age and sampling year effect on the fungal fruit body community in Pinus pinaster forests in central Spain. Can. J. Bot. 2006, 84, 1249–1258. [Google Scholar] [CrossRef]
- Martín-Pinto, P.; Vaquerizo, H.; Peñalver, F.; Olaizola, J.; Oria-De-Rueda, J.A. Early effects of a wildfire on the diversity and production of fungal communities in Mediterranean vegetation types dominated by Cistus ladanifer and Pinus pinaster in Spain. For. Ecol. Manag. 2006, 225, 296–305. [Google Scholar] [CrossRef]
- Yanagihara, H.; Kamo, K.; Imori, S.; Satoh, K. Bias-corrected AIC for selecting variables in multinomial logistic regression models. Linear Algebra Appl. 2012, 436, 4329–4341. [Google Scholar] [CrossRef]
- Ferris, R.; Peace, A.J.; Newton, A.C. Macrofungal communities of lowland Scots pine (Pinus sylvestris L.) and Norway spruce (Picea abies (L.) Karsten.) plantations in England: Relationships with site factors and stand structure. For. Ecol. Manag. 2000, 131, 255–267. [Google Scholar] [CrossRef]
- Tuno, N. Mushroom utilization by the Majangir, an Ethiopian tribe. Mycologist 2001, 15, 78–79. [Google Scholar] [CrossRef]
- Alemu, F. Assessment of wild mushrooms and wood decaying fungi in Dilla University, main campus, Ethiopia. Int. J. Adv. Res. 2013, 1, 458–467. [Google Scholar]
- Muleta, D.; Woyessa, D.; Teferi, Y. Mushroom consumption habits of Wacha Kebele residents, southwestern Ethiopia. Glob. Res. J. Agric. Biol. Sci. 2013, 4, 6–16. [Google Scholar]
- Kassa, H.; Campbell, B.; Sandewall, M.; Kebede, M.; Tesfaye, Y.; Dessie, G.; Seifu, A.; Tadesse, M.; Garedew, E.; Sandewall, K. Building future scenarios and uncovering persisting challenges of participatory forest management in Chilimo Forest, Central Ethiopia. J. Environ. Manag. 2009, 90, 1004–1013. [Google Scholar] [CrossRef] [PubMed]
- Gassibe, P.V.; Oria-de-Rueda, J.A.; Martín-Pinto, P.P. pinaster under extreme ecological conditions provides high fungal production and diversity. For. Ecol. Manag. 2015, 337, 161–173. [Google Scholar] [CrossRef]
- Bonet, J.A.; Fischer, C.R.; Colinas, C. The relationship between forest age and aspect on the production of sporocarps of ectomycorrhizal fungi in Pinus sylvestris forests of the central Pyrenees. For. Ecol. Manag. 2004, 203, 157–175. [Google Scholar] [CrossRef]
- Smith, J.E.; Molina, R.; Huso, M.M.; Luoma, D.L.; McKay, D.; Castellano, M.A.; Lebel, T.; Valachovic, Y. Species richness, abundance, and composition of hypogeous and epigeous ectomycorrhizal fungal sporocarps in young, rotation-age, and old-growth stands of Douglas-fir (Pseudotsuga menziesii) in the Cascade Range of Oregon, U.S.A. Can. J. Bot. 2002, 80, 186–204. [Google Scholar] [CrossRef]
- Bonet, J.A.; Palahí, M.; Colinas, C.; Pukkala, T.; Fischer, C.R.; Miina, J.; Martínez de Aragón, J. Modelling the production and species richness of wild mushrooms in pine forests of the Central Pyrenees in northeastern Spain. Can. J. For. Res. 2010, 40, 347–356. [Google Scholar] [CrossRef]
- Pinna, S.; Gévry, M.F.; Côté, M.; Sirois, L. Factors influencing fructification phenology of edible mushrooms in a boreal mixed forest of Eastern Canada. For. Ecol. Manag. 2010, 260, 294–301. [Google Scholar] [CrossRef]
- Santos-Silva, C.; Gonçalves, A.; Louro, R. Canopy cover influence on macrofungal richness and sporocarp production in montado ecosystems. Agrofor. Syst. 2011, 82, 149–159. [Google Scholar] [CrossRef]
- Straatsma, G.; Ayer, F.; Egli, S. Species richness, abundance, and phenology of fungal fruit bodies over 21 years in a Swiss forest plot. Mycol. Res. 2001, 105, 515–523. [Google Scholar] [CrossRef]
- Reverchon, F.; Del Ortega-Larrocea, P.M.; Pérez-Moreno, J. Saprophytic fungal communities change in diversity and species composition across a volcanic soil chronosequence at Sierra del Chichinautzin, Mexico. Ann. Microbiol. 2010, 60, 217–226. [Google Scholar] [CrossRef]
- Jaleta, D.; Mbilinyi, B.; Mahoo, H.; Lemenih, M. Eucalyptus expansion as relieving and provocative tree in Ethiopia. J. Agric. Ecol. Res. Int. 2016, 6, 1–12. [Google Scholar] [CrossRef]
- Teketay, D. The ecological effects of Eucalyptus: Ground for making wise and informed decision. In Proceedings of the Eucalyptus Dilemma, Addis Ababa, Ethiopia, 15 November 2000. [Google Scholar]
- Castellano, M.; Molina, R. Mycorrhizae. In The Biological Component: Nursery Pests and Mycorrhizal; Landis, T.D., Tinus, R.W., Mc Donald, S.E., Barnett, J.P., Eds.; The Container Tree Nursery Manual. Agriculture Handbook 674, Forest Service; USDA: Washington, DC, USA, 1989; Volume 5, 171p. [Google Scholar]
- Trappe, J. Selection of fungi for ectomycorrhizal inoculation in nurseries. Annu. Rev. Phytopatho. 1977, 15, 203–222. [Google Scholar] [CrossRef]
- Glassman, S.I.; Wang, I.J.; Bruns, T.D. Environmental filtering by pH and soil nutrients drives community assembly in fungi at fine spatial scales. Mol. Ecol. 2017, 26, 6960–6973. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Li, W.; Dumbrell, A.J.; Liu, M.; Li, G.; Wu, M.; Jiang, C.; Li, Z. Spatial Variation in Soil Fungal Communities across Paddy Fields in Subtropical China. mSystems 2020, 5, e00704-19. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Bahram, M.; Polme, S.; Koljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Global diversity and geography of soil fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef]
- Kranabetter, J.M.; Friesen, J.; Gamiet, S.; Kroeger, P. Ectomycorrhizal mushroom distribution by stand age in western hemlock—Lodgepole pine forests of northwestern British Columbia. Can. J. For. Res. 2005, 35, 1527–1539. [Google Scholar] [CrossRef]
- Pérez-Moreno, J.; Martínez-Reyes, M.; Yescas-Pérez, A.; Delgado-Alvarado, A.; Xoconostle-Cázares, B. Wild mushroom markets in central Mexico and a case study at Ozumba. Econ. Bot. 2008, 62, 425–436. [Google Scholar] [CrossRef]
- Collado, E.; Bonet, J.A.; Camarero, J.J.; Egli, S.; Peter, M.; Salo, K.; Martínez-Peña, F.; Ohenoja, E.; Martín-Pinto, P.; Primicia, I.; et al. Mushroom productivity trends in relation to tree growth and climate across different European forest biomes. Sci. Total Environ. 2019, 689, 602–615. [Google Scholar] [CrossRef] [PubMed]
- Martínez de Aragón, J.; Bonet, J.A.; Fischer, C.R.; Colinas, C. Productivity of ectomycorrhizal and selected edible saprotrophic fungi in pine forests of the pre-Pyrenees mountains, Spain: Predictive equations for forest management of mycological resources. For. Ecol. Manag. 2007, 252, 239–256. [Google Scholar] [CrossRef]
- Newsham, K.K.; Hopkins, D.W.; Carvalhais, L.C.; Fretwell, P.T.; Rushton, S.P.; O’Donnell, A.G.; Dennis, P.G. Relationship between soil fungal diversity and temperature in the maritime Antarctic. Nat. Clim. Chang. 2016, 6, 182–186. [Google Scholar] [CrossRef]
- Worku, M. The Role of Forest Biodiversity Conservation Practices for Tourism Development in a Case of Tara Gedam Monastery, South Gonder Zone, Ethiopia. J. Ecosyst. Ecography 2017, 7, 2–7. [Google Scholar] [CrossRef]
- Esubalew, E.; Giday, K.; Hishe, H.; Goshu, G. Carbon stock of woody species along Altitude gradient in Alemsaga Forest, South Gondar, North Western Ethiopia. Agric. J. IJOEAR 2019, 5, 13–21. [Google Scholar]
- Abere, F.; Belete, Y.; Kefalew, A.; Soromessa, T. Carbon stock of Banja forest in Banja district, Amhara region, Ethiopia: An implication for climate change mitigation. J. Sustain. For. 2017, 36, 604–622. [Google Scholar] [CrossRef]
- Alem, D.; Dejene, T.; Oria-de-Rueda, J.A.; Geml, J.; Castaño, C.; Smith, J.E.; Martín-Pinto, P. Soil fungal communities and succession following wildfire in Ethiopian dry Afromontane forests, a highly diverse underexplored ecosystem. For. Ecol. Manag. 2020, 474, 118328. [Google Scholar] [CrossRef]
- Jemal, A.; Getu, E. Diversity of butterfly communities at different altitudes of Menagesha-suba state forest, Ethiopia. J. Entomol. Zool. Stud. 2018, 6, 2197–2202. [Google Scholar]
- Dejene, T.; Oria-de-Rueda, J.A.; Martín-Pinto, P. Fungal diversity and succession under Eucalyptus grandis plantations in Ethiopia. For. Ecol. Manag. 2017, 405, 179–187. [Google Scholar] [CrossRef]
- Gassibe, P.V.; Fabero, R.F.; Hernández-Rodríguez, M.; Oria-de-Rueda, J.A.; Martín-Pinto, P. Fungal community succession following wildfire in a Mediterranean vegetation type dominated by Pinus pinaster in Northwest Spain. For. Ecol. Manag. 2011, 262, 655–662. [Google Scholar] [CrossRef]
- Hernández-Rodríguez, M.; Oria-de-Rueda, J.A.; Martín-Pinto, P. Post-fire fungal succession in a Mediterranean ecosystem dominated by Cistus ladanifer L. For. Ecol. Manag. 2013, 289, 48–57. [Google Scholar] [CrossRef]
- Hiiesalu, I.; Bahram, M.; Tedersoo, L. Plant species richness and productivity determine the diversity of soil fungal guilds in temperate coniferous forest and bog habitats. Mol. Ecol. 2017, 26, 4846–4858. [Google Scholar] [CrossRef]
- Rudolph, S.; Maciá-Vicente, J.G.; Lotz-Winter, H.; Schleuning, M.; Piepenbring, M. Temporal variation of fungal diversity in a mosaic landscape in Germany. Stud. Mycol. 2018, 89, 95–104. [Google Scholar] [CrossRef]
- O’Hanlon, R.; Harrington, T.J. Macrofungal diversity and ecology in four Irish forest types. Fungal Ecol. 2012, 5, 499–508. [Google Scholar] [CrossRef]
- Ruiz-Almenara, C.; Gándara, E.; Gómez-Hernández, M. Comparison of diversity and composition of macrofungal species between intensive mushroom harvesting and non-harvesting areas in Oaxaca, Mexico. PeerJ 2019, 7, e8325. [Google Scholar] [CrossRef] [PubMed]
- Adeniyi, M.; Adeyemi, Y.; Odeyemi, Y.; Odeyemi, O. Ecology, diversity and seasonal distribution of wild mushrooms in a Nigerian tropical forest reserve. Biodiversitas J. Biol. Divers 2018, 19, 285–295. [Google Scholar] [CrossRef]
- Antonin, V. Monograph of Marasmius, Gloiocephala, Palaeocephala and Setulipes in Tropical Africa. Fungus Fl Trop Afr. 2007, 1, 177. [Google Scholar]
- Hama, O.; Maes, E.; Guissou, M.; Ibrahim, D.; Barrage, M.; Parra, L.; Raspe, O.; De Kesel, A. Agaricus subsaharianus, une nouvelle espèce comestible et consomméeau Niger, au Burkina Faso et en Tanzanie. Crypto. Mycol. 2010, 31, 221–234. [Google Scholar]
- Pegler, D.N. Studies on African Agaricales: II *. Kew Bull. 1969, 23, 219–249. [Google Scholar] [CrossRef]
- Morris, B. An annotated check-list of the macrofungi of Malawi. Kirkia 1990, 13, 323–364. [Google Scholar]
- Pegler, D.N. Studies on African Agaricales: 1. Kew Bull. 1968, 21, 499–533. [Google Scholar] [CrossRef]
- Pegler, D.; Rayner, R. A contribution to the Agaric flora of Kenya. Kew Bull. 1969, 23, 347–412. [Google Scholar] [CrossRef]
- Rammeloo, J.; Walleyn, R. The edible fungi of Africa South of the Sahara: A literature survey. Scr. Bot. Belg. 1993, 5, 62. [Google Scholar]
- Ryvarden, L.; Piearce, G.D.; Masuka, A.J. An Introduction to the Larger Fungi of South Central Africa; Baobab Books: Harare, Zimbabwe, 1994. [Google Scholar]
- Singer, R. Marasmius. Fl. Icon. Champ. Congo 1965, 14, 253–278. [Google Scholar]
- Põlme, S.; Abarenkov, K.; Nilsson, R.H.; Lindahl, B.D.; Clemmensen, K.E.; Kauserud, H.; Nguyen, N.; Kjøller, R.; Bates, S.T.; Baldrian, P.; et al. FungalTraits: A user-friendly traits database of fungi and fungus-like stramenopiles. Fungal Divers 2020, 105, 1–16. [Google Scholar] [CrossRef]
- Gerhardt, E.; Vila, J.; Llimona, X. Mushroom from Spain and Europe: Manual for Identification; Omega SA Editions: Barcelona, Spain, 2000; ISBN 84-282-1120-5. [Google Scholar]
- Moreno, G.; Manjón, J.L. Guide to Fungi of the Iberian Peninsula; Omega SA Editions: Barcelona, Spain, 2010. [Google Scholar]
- Reeuwijk, L. Procedures for Soil Analysis, 6th ed.; International Soil Reference and Information Centre: Wageningen, The Netherlands, 2002. [Google Scholar]
- Walkley, A.; Black, I.A. An examination of the digestion method for determining soil organic matter and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 34, 29–38. [Google Scholar] [CrossRef]
- Kim, J.; Kreller, C.R.; Greenberg, M.M. Preparation and Analysis of Oligonucleotides Containing the C4‘-Oxidized Abasic Site and Related Mechanistic Probes. J. Org. Chem. 2005, 70, 8122–8129. [Google Scholar] [CrossRef]
- Kim, H.T. Soil Sampling, Preparation and Analysis; CRC Press: Boca Raton, FL, USA, 1996; pp. 139–145. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; Team, R.C. Nlme: Linear and Nonlinear Mixed Effects Models. R Packag. Version 3.1–126. 2016. Available online: http://CRAN>R-project.org/package=nlme (accessed on 1 August 2022).
- Anderson, M.J. A new method for non parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Zeleke, G.; Dejene, T.; Tadesse, W.; Agúndez, D.; Martín-Pinto, P. Ethnomycological Knowledge of Three Ethnic Groups in Ethiopia. Forests 2020, 11, 875. [Google Scholar] [CrossRef]
- Sitotaw, R.; Mulat, A.; Abate, D. Morphological and molecular studies on Termitomyces species of Menge District, Asossa Zone, Northwest Ethiopia. Sci. Technol. Arts Res. J. 2015, 4, 49–57. [Google Scholar] [CrossRef]
- Faye, M.D.; Weber, J.C.; Abasse, T.A.; Boureima, M.; Larwanou, M.; Bationo, A.B.; Diallo, B.O.; Sigué, H.; Dakouo, J.M.; Samaké, O.; et al. Farmers’ preferences for tree functions and species in the west African sahel. For. Trees Livelihoods 2011, 20, 113–136. [Google Scholar] [CrossRef]
- Sundriyal, M.; Sundriyal, R. Wild edible plants of the Sikkim Himalaya: Nutritive values of selected species. Econ. Bot. 2001, 55, 377–390. [Google Scholar] [CrossRef]
- Abate, D. Wild mushrooms and mushroom cultivation efforts in Ethiopia. WSMBMP Bull. 2014, 11, 1–3. [Google Scholar]
- Yehuala, K. Contribution of Small Scale Mushroom Production for Food Security in the Amhara. In Proceedings of the Experience Sharing and Revitalization Workshop of Amhara Region Food Security Network, Bahir Dar, Ethiopia, 26–27 August 2010. [Google Scholar]
- Alem, D.; Dejene, T.; Oria-de-Rueda, J.A.; Martín-Pinto, P. Survey of macrofungal diversity and analysis of edaphic factors influencing the fungal community of church forests in Dry Afromontane areas of Northern Ethiopia. For. Ecol. Manag. 2021, 496, 119391. [Google Scholar] [CrossRef]
- Mediavilla, O.; Oria-de-Rueda, J.A.; Martín-Pinto, P. Changes in sporocarp production and vegetation following wildfire in a Mediterranean Forest Ecosystem dominated by Pinus nigra in Northern Spain. For. Ecol. Manag. 2014, 331, 85–92. [Google Scholar] [CrossRef]
- Deacon, J. Fungal Biology, 4th ed.; Blackwell Publishing Oxford Press: Malden, MA, USA, 2006. [Google Scholar]
- Powers, J.S.; Montgomery, R.A.; Adair, E.C.; Brearley, F.Q.; DeWalt, S.J.; Castanho, C.T.; Chave, J.; Deinert, E.; Ganzhorn, J.U.; Gilbert, M.E.; et al. Decomposition in tropical forests: A pan-tropical study of the effects of litter type, litter placement and mesofaunal exclusion across a precipitation gradient. J. Ecol. 2009, 97, 801–811. [Google Scholar] [CrossRef]
- Wardle, D.A.; Bardgett, R.; Klironomos, J.; Setala, H.; van der Putten, W.; Wall, D. Ecological Linkages Between Aboveground and Belowground Biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef]
- Cairney, J.W.G.; Bastias, B.A. Influences of fire on forest soil fungal communitiesThis article is one of a selection of papers published in the Special Forum on Towards Sustainable Forestry—The Living Soil: Soil Biodiversity and Ecosystem Function. Can. J. For. Res. 2007, 37, 207–215. [Google Scholar] [CrossRef]
- Toivanen, T.; Markkanen, A.; Kotiaho, J.S.; Halme, P. The effect of forest fuel harvesting on the fungal diversity of clear-cuts. Biomass Bioenergy 2012, 39, 84–93. [Google Scholar] [CrossRef]
- Elizabeth, M.-E.; Felipe, R.-S.; Maribel, I.-M. Conocimiento Popular Acerca De La K’Allampa De Pino (Suillus luteus (L) Roussel) En La Localidad De Alalay, Mizque (Cochabamba, Bolivia): Un Ejemplode Dilogo De Saberes. Revista Etnobiologia 2018, 16, 76–86. [Google Scholar]
- Gómez-Hernández, M.; Williams-Linera, G. Diversity of macromycetes determined by tree species, vegetation structure, and microenvironment in tropical cloud forests in Veracruz, Mexico. Botany 2011, 89, 203–216. [Google Scholar] [CrossRef]
- Read, D.; Perez-Moreno, J. Mycorrhizas and nutrient cycling in ecosystems—A journey towards relevance? New Phytol. 2003, 157, 475–492. [Google Scholar] [CrossRef]
- Kirk, P.; Cannon, P.; Minter, D.; Stalpers, J. Dictionary of the Fungi, 10th ed.; The Centre for Agriculture and Bioscience International (CABI): Wallingford, UK, 2008. [Google Scholar]
- Crowther, T.W.; Stanton, D.W.G.; Thomas, S.M.; A’Bear, A.D.; Hiscox, J.; Jones, T.H.; Voříšková, J.; Baldrian, P.; Boddy, L. Top-down control of soil fungal community composition by a globally distributed keystone consumer. Ecology 2013, 94, 2518–2528. [Google Scholar] [CrossRef]
- Koide, R.T.; Fernandez, C.; Malcolm, G. Determining place and process: Functional traits of ectomycorrhizal fungi that affect both community structure and ecosystem function. New Phytol. 2014, 201, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Cozzolino, V.; Di Meo, V.; Monda, H.; Spaccini, R.; Piccolo, A. The molecular characteristics of compost affect plant growth, arbuscular mycorrhizal fungi, and soil microbial community composition. Biol. Fertil. Soils 2016, 52, 15–29. [Google Scholar] [CrossRef]
- Harrington, T.J. Relationships between macrofungi and vegetation in the burren. Biol. Environ. 2003, 103, 147–159. [Google Scholar] [CrossRef]
- Kranabetter, J.M.; Durall, D.M.; MacKenzie, W.H. Diversity and species distribution of ectomycorrhizal fungi along productivity gradients of a southern boreal forest. Mycorrhiza 2009, 19, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Trudell, S.A.; Edmonds, R.L. Macrofungus communities correlate with moisture and nitrogen abundance in two old-growth conifer forests, Olympic National Park, Washington, USA. Can. J. Bot. 2004, 82, 781–800. [Google Scholar] [CrossRef]
- Toljander, J.F.; Eberhardt, U.; Toljander, Y.K.; Paul, L.R.; Taylor, A.F.S. Species composition of an ectomycorrhizal fungal community along a local nutrient gradient in a boreal forest. New Phytol. 2006, 170, 873–884. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest | Geographical Coordinates | Altitude (m asl) | MAP (mm) | MAT (°C) | Vegetation Types | Reference |
|---|---|---|---|---|---|---|
| Taragedam | 12°06′59″–12°07′25″ N and 37°46′14″–37°47′02″ E | 2062–2457 | 1300 | 20.4 | Natural forests | [48] |
| Alemsaga | 11°54′30″–11°56′00″ N and 37°55′00″–37°57′00″ E | 2100–2470 | 1484 | 16.4 | Natural forests | [49] |
| Banja | 10°57′17″–11°03′05″ N and 36°39′09″–36°48′25″ E | 1870–2570 | 1215.3 | 17.7 | Natural forests | [50] |
| Wondo Genet | 7°06′–7°07′ N and 38°37′–38°42′ E | 1600–2580 | 1210 | 20 | Natural forests and Pinus patula and Eucalyptus grandis plantation forests | [51] |
| Menagesha Suba | 8°56′–9°03′ N and 38°28′–38°36′ E | 2200–3385 | 1100 | 16 | Pinus radiata plantation forests | [52] |
| Species | Family | T | N | P |
|---|---|---|---|---|
| Agaricus augustus Fr. | Agaricaceae | S | x | |
| Agaricus bitorquis (Quél.) Sacc. | Agaricaceae | S | x | |
| Agaricus campestris L. | Agaricaceae | S | x | |
| Agaricus campestroides Heinem. and Gooss. -Font. | Agaricaceae | S | x | x |
| Agaricus moelleri Wasser | Agaricaceae | S | x | |
| Agaricus murinaceus Bull. | Agaricaceae | S | x | |
| Agaricus subedulis Heinem. | Agaricaceae | S | x | x |
| Ampulloclitocybe clavipes (Pers.) Redhead, Lutzoni, Moncalvo and Vilgalys | Hygrophoraceae | S | x | |
| Armillaria heimii Pegler | Physalacriaceae | P | x | |
| Auricularia auricula-judae (Bull.) Quél. | Auriculariaceae | S | x | |
| Calvatia cyathiformis (Bosc) Morgan | Lycoperdaceae | S | x | |
| Calvatia gigantea (Batsch) Lloyd | Lycoperdaceae | S | x | |
| Calvatia subtomentosa Dissing and M. Lange | Lycoperdaceae | S | x | |
| Clitocybe carolinensis H.E. Bigelow and Hesler | Tricholomataceae | S | x | |
| Clitocybe cistophila Bon and Contu | Tricholomataceae | S | x | |
| Clitocybe foetens Melot | Tricholomataceae | S | x | |
| Clitocybe fragrans (With.) P. Kumm. | Tricholomataceae | S | x | |
| Clitocybe geotropa (Bull. ex DC.) Quél. | Tricholomataceae | S | x | |
| Coprinellus domesticus (Bolton) Vilgalys, Hopple and Jacq. Johnson | Psathyrellaceae | S | x | x |
| Coprinopsis nivea (Pers.) Redhead, Vilgalys and Moncalvo | Psathyrellaceae | S | x | |
| Coprinus comatus (O.F. Müll.) Pers. | Agaricaceae | S | x | |
| Coprinus lagopus (Fr.) Fr. | Psathyrellaceae | S | x | |
| Coprinus micaceus (Bull.) Fr. | Psathyrellaceae | S | x | |
| Coprinus niveus (Pers.) Fr. | Psathyrellaceae | S | x | |
| Craterellus ignicolor (R.H. Petersen) Dahlman, Danell and Spatafora | Hydnaceae | S | x | |
| Crepidotus applanatus (Pers.) P. Kumm. | Crepidotaceae | S | x | |
| Crepidotus mollis (Schaeff.) Staude | Crepidotaceae | S | x | |
| Gymnopilus pampeanus (Speg.) Singer | Strophariaceae | S | x | |
| Hygrocybe chlorophana (Fr.) Wünsche | Hygrophoraceae | S | x | |
| Hygrophoropsis aurantiaca (Wulfen) Maire | Hygrophoropsidaceae | S | x | x |
| Laetiporus sulphureus (Bull.) Murrill | Laetiporaceae | S | x | |
| Lentinellus cochleatus (Pers.) P. Karst. | Auriscalpiaceae | S | x | |
| Lepista sordida (Schumach.) Singer | Tricholomataceae | S | x | |
| Lepista sordida var. lilacea (Quél.) Bon | Tricholomataceae | S | x | |
| Leucoagaricus americanus (Peck) Vellinga | Agaricaceae | S | x | |
| Leucoagaricus holosericeus (J.J. Planer) M.M. Moser | Agaricaceae | S | x | x |
| Leucoagaricus leucothites (Vittad.) Wasser | Agaricaceae | S | x | x |
| Leucoagaricus purpureolilacinus Huijsman | Agaricaceae | S | x | |
| Leucoagaricus rubrotinctus (Peck) Singer | Agaricaceae | S | x | x |
| Leucocoprinus birnbaumii (Corda) Singer | Agaricaceae | S | x | |
| Leucocoprinus cepistipes (Sowerby) Pat. | Agaricaceae | S | x | x |
| Lycoperdon perlatum Pers. | Lycoperdaceae | S | x | |
| Lycoperdon umbrinum Pers. | Lycoperdaceae | S | x | |
| Macrolepiota africana (R. Heim) Heinem. | Agaricaceae | S | x | |
| Macrolepiota procera (Scop.) Singer | Agaricaceae | S | x | |
| Morchella cf americana Clowez and C. Matherly | Morchellaceae | S | x | |
| Morchella anatolica Isiloglu, Spooner, Alli and Solak | Morchellaceae | S | x | |
| Omphalotus illudens (Schwein.) Bresinsky and Besl | Omphalotaceae | S | x | |
| Pholiota aurivella (Batsch) P. Kumm. | Strophariaceae | S | x | |
| Pleurotus luteoalbus Beeli | Pleurotaceae | S | x | |
| Pleurotus populinus O. Hilber and O.K. Mill. | Pleurotaceae | S | x | |
| Pleurotus pulmonarius (Fr.) Quél. | Pleurotaceae | S | x | |
| Polyporus badius (Pers.) Schwein. | Polyporaceae | S | x | |
| Polyporus tenuiculus (P. Beauv.) Fr. | Polyporaceae | S | x | |
| Polyporus tuberaster (Jacq. ex Pers.) Fr. | Polyporaceae | S | x | |
| Schizophyllum commune Fr. | Schizophyllaceae | S | x | |
| Suillus luteus (L.) Roussel | Suillaceae | EM | x | |
| Termitomyces clypeatus R. Heim | Lyophyllaceae | S | x | |
| Termitomyces microcarpus (Berk. and Broome) R. Heim | Lyophyllaceae | S | x | |
| Termitomyces robustus (Beeli) R. Heim | Lyophyllaceae | S | x | |
| Termitomyces schimperi (Pat.) R. Heim | Lyophyllaceae | S | x | |
| Tricholoma portentosum (Fr.) Quél. | Tricholomataceae | EM | x | |
| Tricholoma saponaceum (Fr.) P. Kumm. | Tricholomataceae | EM | x | |
| Tylopilus niger (Heinem. and Gooss. -Font.) Wolfe | Boletaceae | EM | x |
| Sources | Contribution | Variables | Pseudo-F | p-Values |
|---|---|---|---|---|
| Edaphic variables | 32.33% | OM | 0.2824 | 0.001 |
| N | 0.3057 | 0.001 | ||
| P | 0.4184 | 0.001 | ||
| OC | 0.3122 | 0.001 | ||
| Climate | 60.76% | Elevation | 0.0762 | 0.088 |
| Tmin | 0.5217 | 0.001 | ||
| Spatial factors | 64.19% | Latitude | 0.7777 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kewessa, G.; Dejene, T.; Alem, D.; Tolera, M.; Martín-Pinto, P. Forest Type and Site Conditions Influence the Diversity and Biomass of Edible Macrofungal Species in Ethiopia. J. Fungi 2022, 8, 1023. https://doi.org/10.3390/jof8101023
Kewessa G, Dejene T, Alem D, Tolera M, Martín-Pinto P. Forest Type and Site Conditions Influence the Diversity and Biomass of Edible Macrofungal Species in Ethiopia. Journal of Fungi. 2022; 8(10):1023. https://doi.org/10.3390/jof8101023
Chicago/Turabian StyleKewessa, Gonfa, Tatek Dejene, Demelash Alem, Motuma Tolera, and Pablo Martín-Pinto. 2022. "Forest Type and Site Conditions Influence the Diversity and Biomass of Edible Macrofungal Species in Ethiopia" Journal of Fungi 8, no. 10: 1023. https://doi.org/10.3390/jof8101023