

A Molecular Systematics and Taxonomy Research on Trechispora (Hydnodontaceae, Trechisporales): Concentrating on Three New Trechispora Species from East Asia

Abstract
:1. Introduction
2. Materials and Methods
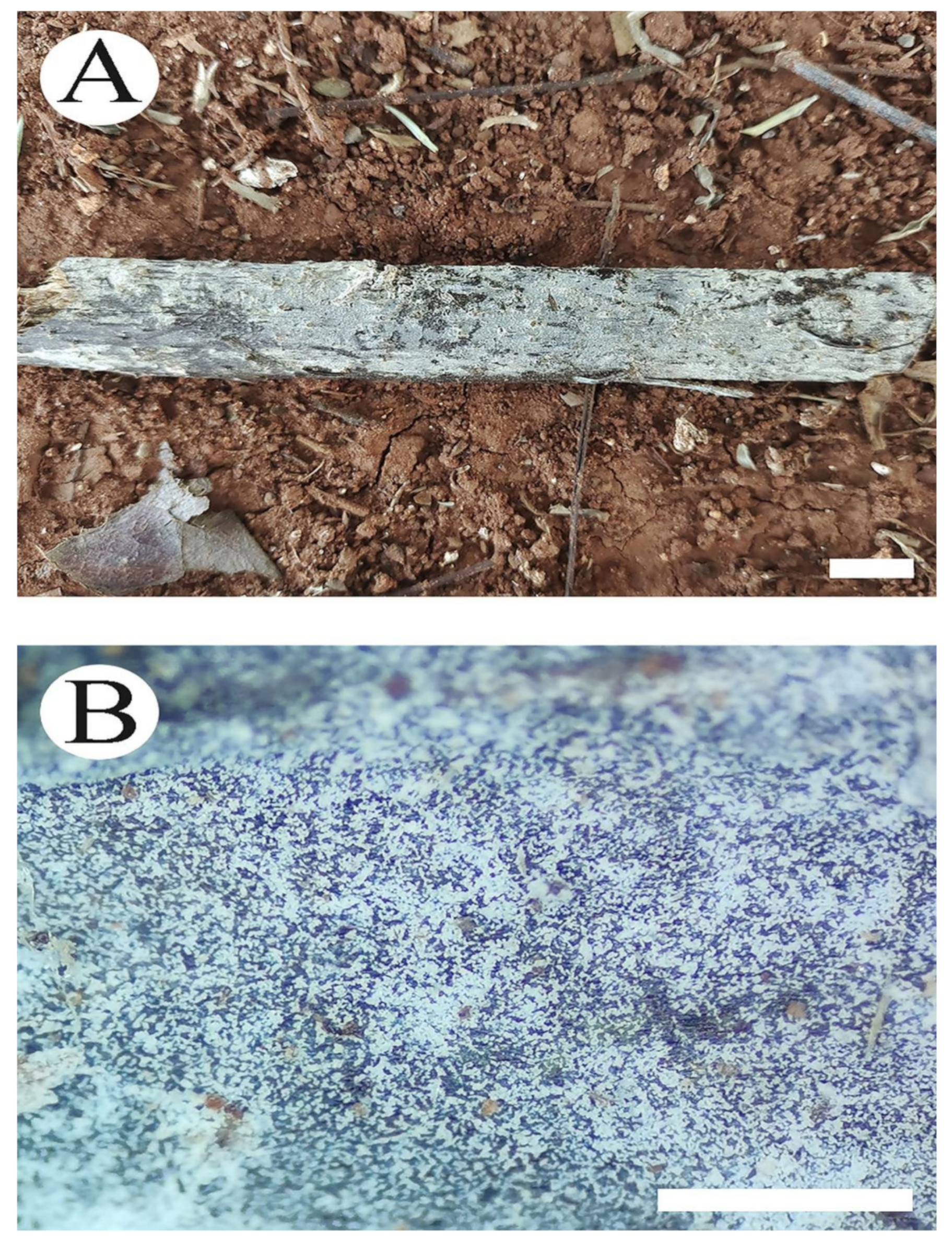
2.1. Sample Collection and Herbarium Specimen Preparation
2.2. Morphology
2.3. Molecular Phylogeny
3. Results
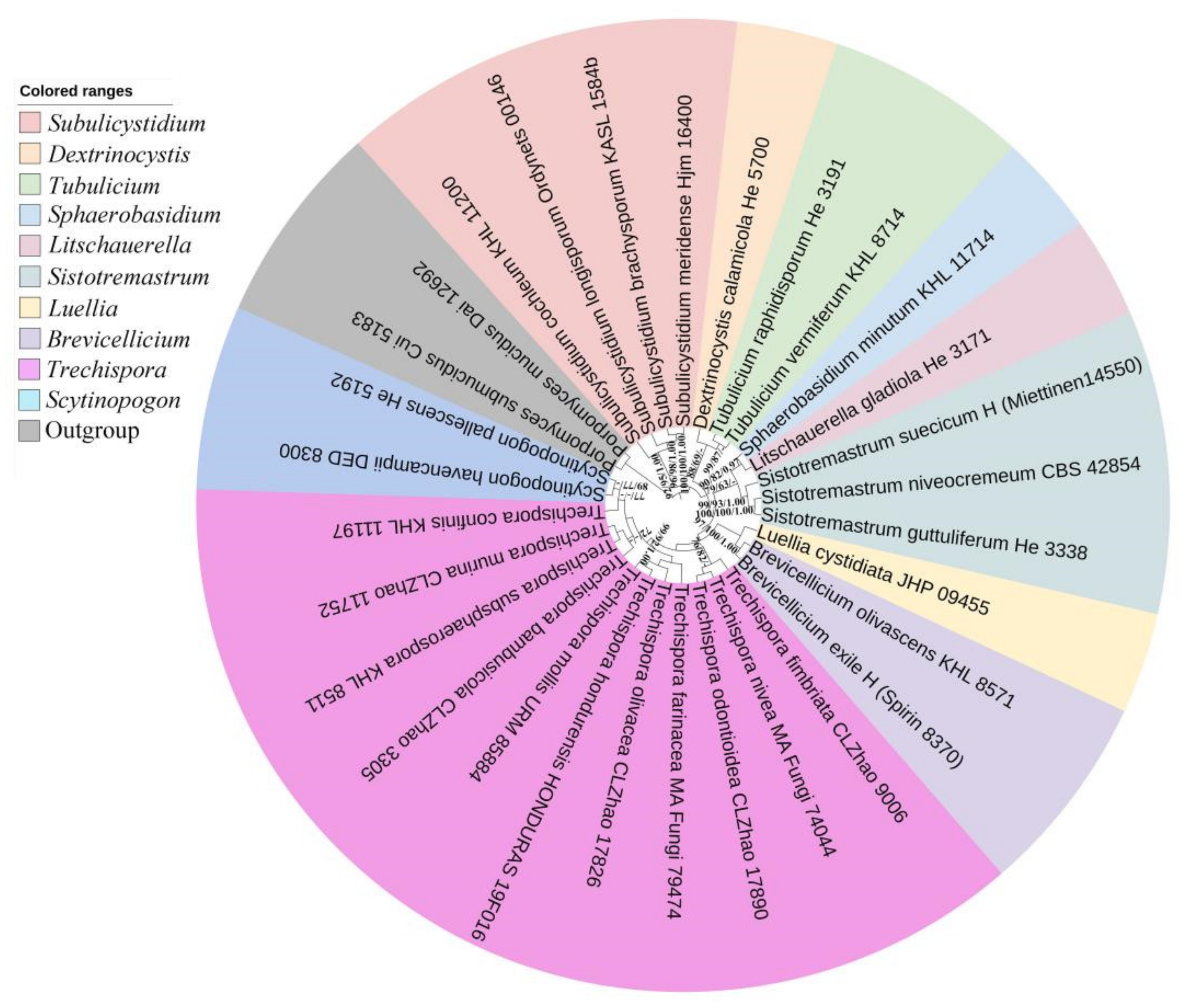
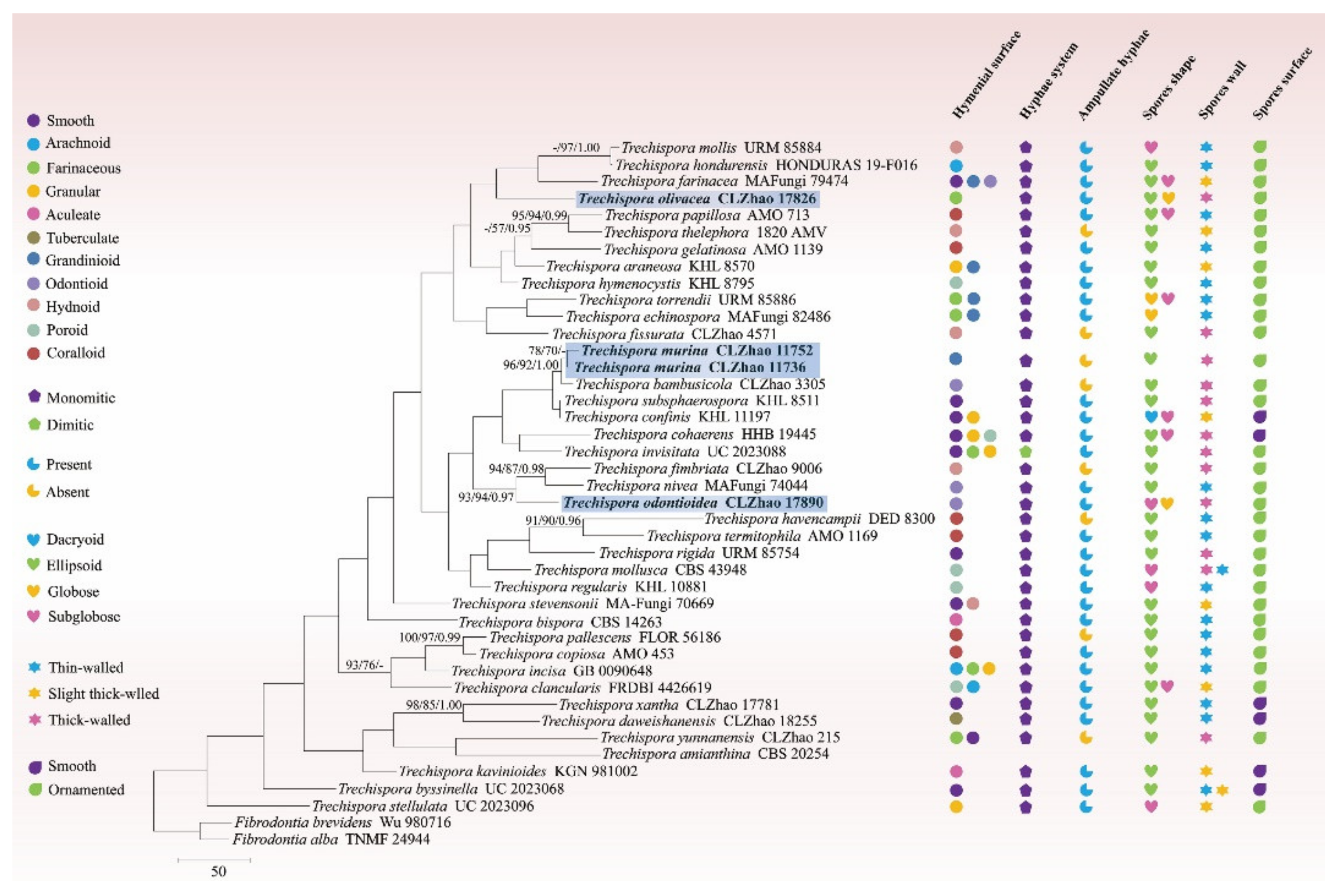
3.1. Molecular Phylogeny
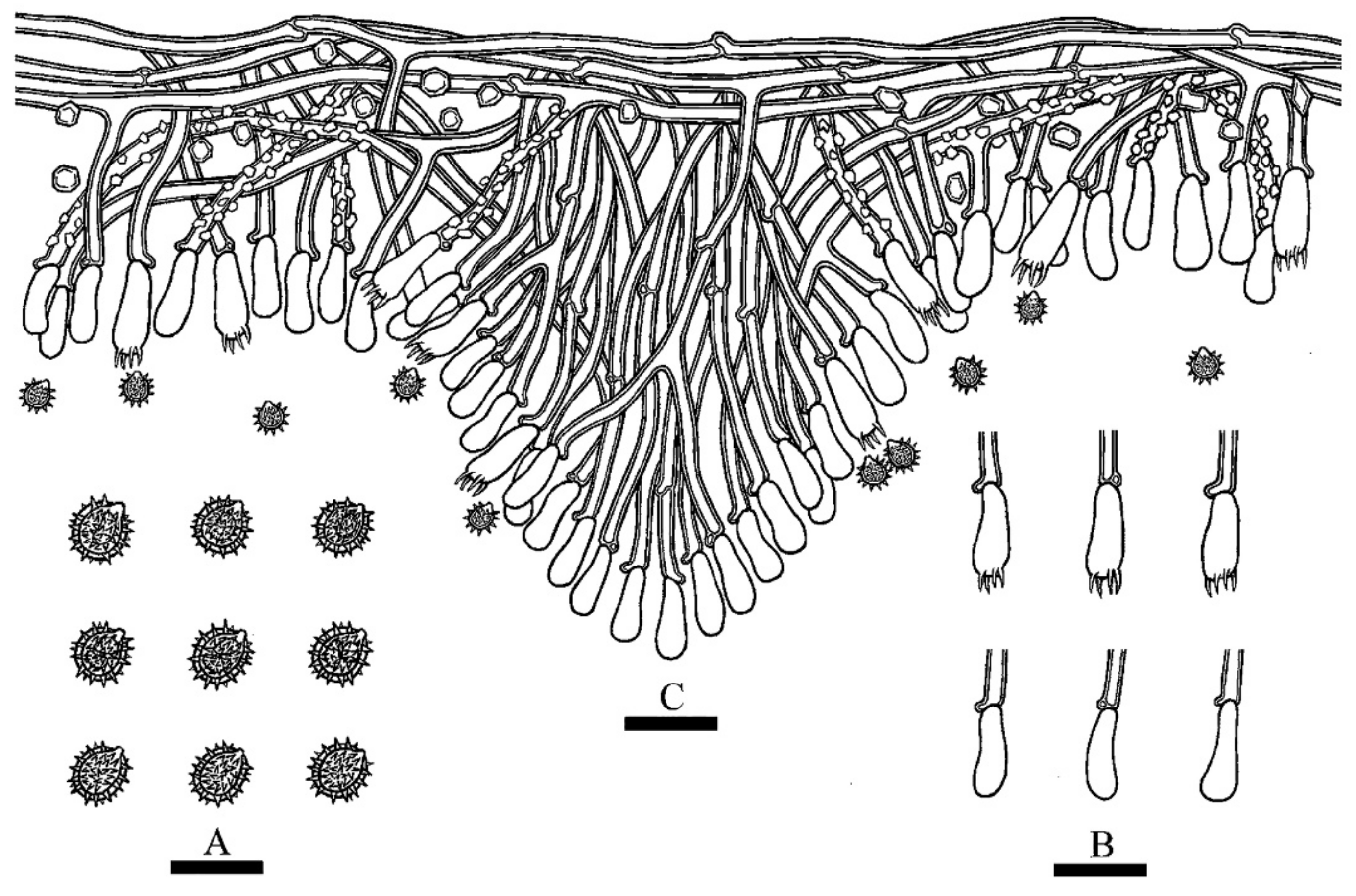
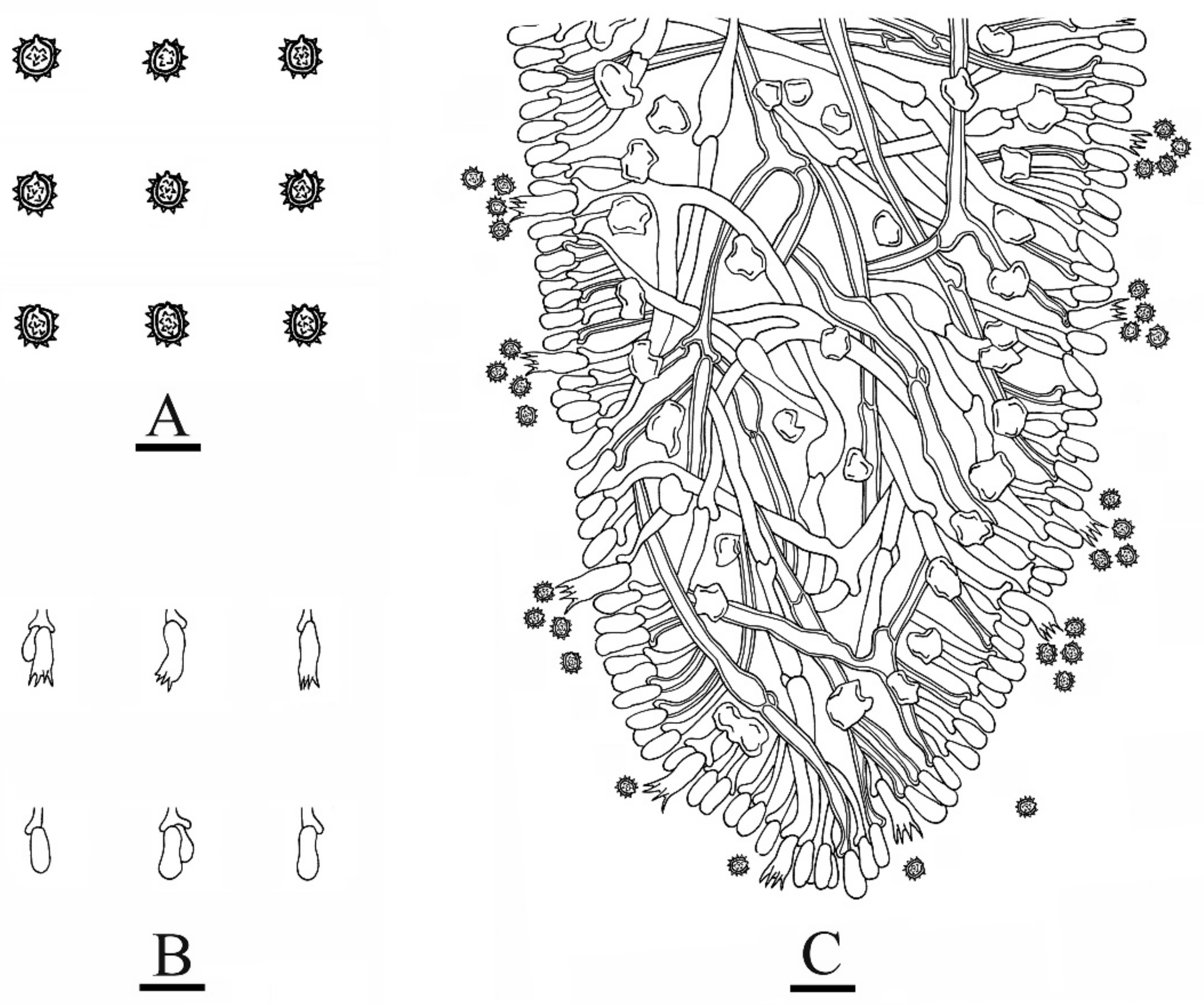
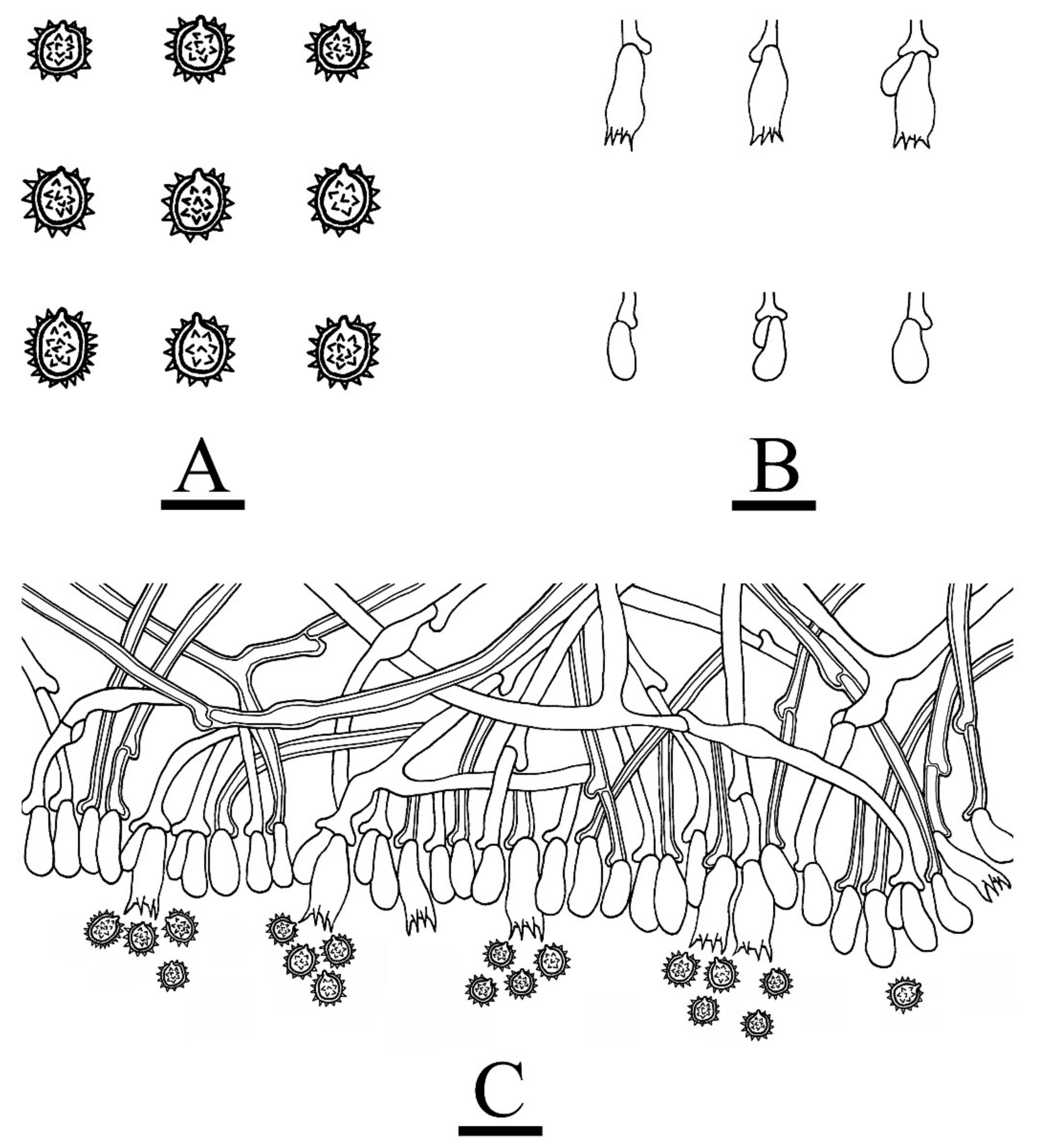
3.2. Taxonomy
4. Discussion
- 1. Basidiospores smooth-------------------------------------------------------------------------------------2
- 1′ Basidiospores aculeate, verrucose or ornamented--------------------------------------------------5
- 2. Ampullate hyphae > 5 μm in width, basidiospores angular----------------T. subsphaerospora
- 2′ Ampullate hyphae < 5 μm in width, basidiospores ellipsoid------------------------------------3
- 3. Basidiospores thick-walled-----------------------------------------------------------------T. cohaerens
- 3′ Basidiospores thin-walled--------------------------------------------------------------------------------4
- 4. Hymenial surface tuberculate-------------------------------------------------------T. daweishanensis
- 4′ Hymenial surface smooth----------------------------------------------------------------------T. xantha
- 5. Hyphal system dimitic------------------------------------------------------------------------T. dimitica
- 5′ Hyphal system monomitic-------------------------------------------------------------------------------6
- 6. Hyphae without ampullate septa----------------------------------------------------------------------7
- 6′ Hyphae with ampullate septa-------------------------------------------------------------------------12
- 7. Basidiospores thin-walled, ovoid to subglobose---------------------------------------T. suberosa
- 7′ Basidiospores thick-walled, ellipsoid-----------------------------------------------------------------8
- 8. Basidiospores > 7 μm in length--------------------------------------------------------T. yunnanensis
- 8′ Basidiospores < 7 μm in length-------------------------------------------------------------------------9
- 9. Basidiomata margin greyish----------------------------------------------------------------T. murina
- 9′ Basidiomata margin white to cream----------------------------------------------------------------10
- 10. Hymenial surface odontioid---------------------------------------------------------T. bambusicola
- 10′ Hymenial surface hydnoid--------------------------------------------------------------------------11
- 11. Hymenophore with blunt aculei--------------------------------------------------------T. fimbriata
- 11′ Hymenophore with sharp aculei--------------------------------------------------------T. fissurata
- 12. Sphaerocysts present, hyphae inflated-------------------------------------------T. hymenocystis
- 12′ Sphaerocysts absent, hyphae uninflated---------------------------------------------------------13
- 13. Ampullate septa > 6 μm in width------------------------------------------------------------------14
- 13′ Ampullate septa < 6 μm in width------------------------------------------------------------------15
- 14. Basidiospores sparsely verrucose-----------------------------------------------T. polygonospora
- 14′ Basidiospores densely aculeate---------------------------------------------------------T. mollusca
- 15. Subhymenium with short-celled hyphae--------------------------------------------------------16
- 15′ Subhymenium with long-celled hyphae---------------------------------------------------------17
- 16. Basidiome thin, ochraceous--------------------------------------------------------------T. farinacea
- 16′ Basidiome thick, dirty white to buff-------------------------------------------------------T. rigida
- 17. Basidiospores thin-walled---------------------------------------------------------------------------18
- 17′ Basidiospores thick-walled--------------------------------------------------------------------------19
- 18. Hymenophore with hydnoid-----------------------------------------------------------------T. nivea
- 18′ Hymenophore without hydnoid------------------------------------------------------T. microspora
- 19. Basidiospores > 5 μm in length---------------------------------------------------------T. praefocata
- 19′ Basidiospores < 5 μm in length---------------------------------------------------------------------20
- 20. Hymenial surface farinaceous with olivaceous--------------------------------------T. olivacea
- 20′ Hymenial surface odontioid with buff--------------------------------------------T. odontioidea
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- James, T.Y.; Stajich, J.E.; Hittinger, C.T.; Rokas, A. Toward a fully resolved fungal tree of life. Annu. Rev. Microbiol. 2020, 74, 291–313. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Bahram, M.; Põlme, S.; Koljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Global diversity and geography of soil fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef] [PubMed]
- Gilbertson, R.L.; Ryvarden, L. North American Polypores 1–2; Fungiflora: Oslo, Norway, 1987; pp. 1–433. [Google Scholar]
- Núñez, M.; Ryvarden, L. East Asian polypores 2. Synop. Fungorum 2001, 14, 165–522. [Google Scholar]
- Bernicchia, A.; Gorjón, S.P. Fungi Europaei 12: Corticiaceae s.l.; Edizioni Candusso: Alassio, Italy, 2010. [Google Scholar]
- Dai, Y.C. Polypore diversity in China with an annotated checklist of Chinese polypores. Mycoscience 2012, 53, 49–80. [Google Scholar] [CrossRef]
- Ryvarden, L.; Melo, I. Poroid fungi of Europe. Synop. Fungorum 2014, 31, 1–455. [Google Scholar]
- Dai, Y.C.; Cui, B.K.; Si, J.; He, S.H.; Hyde, K.D.; Yuan, H.S.; Liu, X.Y.; Zhou, L.W. Dynamics of the worldwide number of fungi with emphasis on fungal diversity in China. Mycol. Prog. 2015, 14, 62. [Google Scholar] [CrossRef]
- Dai, Y.C.; Yang, Z.L.; Cui, B.K.; Wu, G.; Yuan, H.S.; Zhou, L.W.; He, S.H.; Ge, Z.W.; Wu, F.; Wei, Y.L.; et al. Diversity and systematics of the important macrofungi in Chinese forests. Mycosystema 2021, 40, 770–805. [Google Scholar]
- Cui, B.K.; Yuan, H.S.; Zhou, L.W.; He, S.H.; Wei, Y.L. Diversity of wood-decaying fungi in conifer trees of the Greater and Lesser Khinggan Mountains. Biodivers. Sci. 2019, 27, 887–895. [Google Scholar] [CrossRef]
- Wu, F.; Yuan, H.S.; Zhou, L.W.; Yuan, Y.; Cui, B.K.; Dai, Y.C. Polypore diversity in South China. Mycosystema 2020, 39, 653–682. [Google Scholar]
- Luo, K.Y.; Chen, Z.Y.; Zhao, C.L. Phylogenetic and taxonomic analyses of three new wood-inhabiting fungi of Xylodon (Basidiomycota) in a forest ecological system. J. Fungi 2022, 8, 405. [Google Scholar] [CrossRef]
- Qu, M.H.; Wang, D.Q.; Zhao, C.L. A phylogenetic and taxonomic study on Xylodon (Hymenochaetales): Focusing on three new Xylodon species from southern China. J. Fungi 2022, 8, 35. [Google Scholar] [CrossRef]
- Hibbett, D.S.; Binder, M.; Bischoff, J.F.; Blackwell, M.; Cannon, P.F.; Eriksson, O.E.; Huhndorf, S.; James, T. A higher-level phylogenetic classification of the Fungi. Mycol. Res. 2007, 111, 509–547. [Google Scholar] [CrossRef]
- Larsson, K.H. Re-thinking the classification of corticioid fungi. Mycol. Res. 2007, 111, 1040–1063. [Google Scholar] [CrossRef]
- Karsten, P.A. Fragmenta mycologica XXIX. Nova Hedwig. 1890, 29, 147–149. [Google Scholar]
- Liberta, A.E. On Trechispora. Taxon 1966, 15, 317–319. [Google Scholar] [CrossRef]
- Liberta, A.E. The genus Trechispora (Basidiomycetes, Corticiaceae). Can. J. Bot. 1973, 51, 1871–1892. [Google Scholar] [CrossRef]
- Larsson, K.H. Poroid species in Trechispora and the use of calcium oxalate crystals for species identification. Mycol. Res. 1994, 98, 1153–1172. [Google Scholar] [CrossRef]
- Ryvarden, L. A note on the genus Hydnodon Banker. Synop. Fungorum 2002, 15, 31–33. [Google Scholar]
- Trichiès, G.; Schultheis, B. Trechispora antipus sp. nov., une seconde espèce bisporique du genre Trechispora (Basidiomycota, Stereales). Mycotaxon 2002, 82, 453–458. [Google Scholar]
- Miettinen, O.; Larsson, K.H. Trechispora elongata species nova from North Europe. Mycotaxon 2006, 96, 193–198. [Google Scholar]
- Kirk, P.M.; Cannon, P.F.; David, J.C.; Minter, D.W.; Stalpers, J.A. Ainsworth and Bisby’s Dictionary of the Fungi, 10th ed.; CAB International Press: Wallingford, UK, 2008; p. 783. [Google Scholar] [CrossRef]
- Ordynets, A.; Larsson, K.H.; Langer, E. Two new Trechispora species from La Réunion Island. Mycol. Prog. 2015, 14, 113. [Google Scholar] [CrossRef]
- Phookamsak, R.; Hyde, K.D.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Raspe´, O.; Karunarathna, S.C.; Wanasinghe, D.N.; Hongsanan, S.; et al. Fungal diversity notes 929–1035: Taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers. 2019, 95, 1–273. [Google Scholar] [CrossRef]
- Chikowski, R.S.; Larsson, K.H.; Gibertoni, T.B. Taxonomic novelties in Trechispora from Brazil. Mycol. Prog. 2020, 19, 1403–1414. [Google Scholar] [CrossRef]
- Meiras-Ottoni, A.; Larsson, K.H.; Gibertoni, T.B. Additions to Trechispora and the status of Scytinopogon (Trechisporales, Basidiomycota). Mycol. Prog. 2021, 20, 203–222. [Google Scholar] [CrossRef]
- Dai, Y.C. A checklist of polypores in China. Mycosystema 2009, 28, 315–327. [Google Scholar]
- Dai, Y.C. A revised checklist of corticioid and hydnoid fungi in China for 2010. Mycoscience 2011, 52, 69–79. [Google Scholar] [CrossRef]
- Yuan, H.S.; Dai, Y.C. Wood-inhabiting fungi in southern China. 6. Polypores from Guangxi Autonomous region. Ann. Bot. Fenn. 2012, 49, 341–351. [Google Scholar] [CrossRef]
- Xu, T.M.; Chen, Y.H.; Zhao, C.L. Trechispora yunnanensis sp. nov. from China. Phytotaxa 2019, 424, 253–261. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Zhao, M.J.; Su, J.H.; Yang, L.; Deng, H.; Wang, Y.H.; Wu, H.J.; Li, Y.; Wu, H.M.; Wei, X.D.; et al. The use of checklist of fungi in China database in the red list assessment of macrofungi in China. Biodivers. Sci. 2020, 28, 74–98. [Google Scholar] [CrossRef]
- Zhao, W.; Zhao, C.L. The phylogenetic relationship revealed three new wood-inhabiting fungal species from genus Trechispora. Front. Microbiol. 2021, 12, 650195. [Google Scholar] [CrossRef] [PubMed]
- Zong, T.K.; Liu, C.M.; Wu, J.R.; Zhao, C.L. Trechispora daweishanensis and T. xantha spp. nov. (Hydnodontaceae, Trechisporales) found in Yunnan Province of China. Phytotaxa 2021, 479, 147–159. [Google Scholar] [CrossRef]
- Larsson, K.H.; Larsson, E.; Kõljalg, U. High phylogenetic diversity among corticioid homobasidiomycetes. Mycol. Res. 2004, 108, 983–1002. [Google Scholar] [CrossRef]
- Telleria, M.T.; Melo, I.; Dueñas, M.; Larsson, K.H.; Paz, M.M.P. Molecular analyses confifirm Brevicellicium in Trechisporales. IMA Fungus 2013, 4, 21–28. [Google Scholar] [CrossRef]
- Jülich, W. Notes on some Basidiomycetes (Aphyllophorales and Heterobasidiomycetes). Persoonia 1982, 11, 421–428. [Google Scholar]
- Liu, S.L.; Ma, H.X.; He, S.H.; Dai, Y.C. Four new corticioid species in Trechisporales (Basidiomycota) from East Asia and notes on phylogeny of the order. MycoKeys 2019, 48, 97–113. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J. Metheun Hand Book of Colour, 3rd ed.; Metheun London Ltd.: London, UK, 1978; pp. 144–148. [Google Scholar]
- Wu, F.; Zhou, L.W.; Vlasák, J.; Dai, Y.C. Global diversity and systematics of Hymenochaetaceae with poroid hymenophore. Fungal Divers. 2022, 113, 1–192. [Google Scholar] [CrossRef]
- Zhao, C.L.; Wu, Z.Q. Ceriporiopsis kunmingensis sp. nov. (Polyporales, Basidiomycota) evidenced by morphological characters and phylogenetic analysis. Mycol. Prog. 2017, 16, 93–100. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. A Guide Methods Appl. 1990, 18, 315–322. [Google Scholar] [CrossRef]
- Spirin, V.; Volobuev, S.; Viner, I.; Miettinen, O.; Vlasak, J.; Schoutteten, N.; Motato-Vásquez, V.; Kotiranta, H.; Hernawati; Larsson, K.H. On Sistotremastrum and similar-looking taxa (Trechisporales, Basidiomycota). Mycol. Prog. 2021, 20, 453–476. [Google Scholar] [CrossRef]
- Yurchenko, E.; Wu, S.H. Hyphoderma formosanum sp. nov. (Meruliaceae, Basidiomycota) from Taiwan. Sydowia 2014, 66, 19–23. [Google Scholar]
- Wu, F.; Yuan, Y.; Zhao, C.L. Porpomyces submucidus (Hydnodontaceae, Basidiomycota) from tropical China. Phytotaxa 2015, 230, 61–68. [Google Scholar] [CrossRef]
- Desjardin, D.E.; Perry, B.A. A new species of Scytinopogon from the island of Príncipe, Republic of São Tomé and Príncipe, West Africa. Mycosphere 2015, 6, 434–441. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of fifilamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Larsson, K.H.; Parmasto, E.; Fischer, M.; Langer, E.; Nakasone, K.K.; Redhead, S.A. Hymenochaetales: A molecular phylogeny for the hymenochaetoid clade. Mycologia 2006, 98, 926–936. [Google Scholar] [CrossRef] [PubMed]
- Ordynets, A.; Scherf, D.; Pansegrau, F.; Denecke, J.; Langer, E. Short-spored Subulicystidium (Trechisporales, Basidiomycota): High morphological diversity and only partly clear species boundaries. MycoKeys 2018, 35, 41–99. [Google Scholar] [CrossRef]
- Ordynets, A.; Liebisch, R.; Lysenko, L.; Scherf, D.; Volobuev, S.; Saitta, A.; Larsson, K.H.; Yurchenko, E.; Buyck, B.; Bolshakov, S. Morphologically similar but not closely related: The long-spored species of Subulicystidium (Trechisporales, Basidiomycota). Mycol. Prog. 2020, 19, 691–703. [Google Scholar] [CrossRef]
- Vasco-Palacios, A.M.; Lopez-Quintero, C.; Franco-Molano, A.E.; Boekhout, T. Austroboletus amazonicus sp. nov. and Fistulinella campinaranae var. scrobiculata, two commonly occurring boletes from a forest dominated by Pseudomonotes tropenbosii (Dipterocarpaceae) in Colombian Amazonia. Mycologia 2014, 106, 1004–1014. [Google Scholar] [CrossRef] [PubMed]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large data sets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef] [PubMed]
- Swofford, D.L. PAUP *: Phylogenetic Analysis Using Parsimony (* and Other Methods); Version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Felsenstein, J. Confidence intervals on phylogenetics: An approach using bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. The CIPRES science gateway: Enabling high-impact science for phylogenetics researchers with limited resources. Assoc. Comput. Mach. 2012, 39, 1–8. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModeltest V2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. Mrbayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Haelewaters, D.; Dima, B.; Abdel-Hafiz, A.I.; Abdel-Wahab, M.A.; Abul-Ezz, S.R.; Acar, I.; Aguirre-Acosta, E.; Aime, M.C.; Aldemir, S.; Ali, M.; et al. Fungal systematics and evolution: Fuse 6. Sydowia 2020, 72, 231–356. [Google Scholar] [CrossRef]
- Larsson, K.H. New species and combinations in Trechispora (Corticiaceae, Basidiomycotina). Nord. J. Bot. 1996, 16, 83–98. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, C.Q.; Gan, M.S.; Li, Y.; Shao, S.C.; Qin, W.Q.; Deng, W.Q.; Li, T.H. Diversity of Cantharellus (Cantharellales, Basidiomycota) in China with Description of Some New Species and New Records. J. Fungi 2022, 8, 483. [Google Scholar] [CrossRef]
- Song, C.G.; Chen, Y.Y.; Liu, S.; Xu, T.M.; He, X.L.; Wang, D.; Cui, B.K. A Phylogenetic and Taxonomic Study on Phellodon (Bankeraceae, Thelephorales) from China. J. Fungi 2022, 8, 429. [Google Scholar] [CrossRef]
- Hu, J.; Zhao, G.; Tuo, Y.; Rao, G.; Zhang, Z.; Qi, Z.; Yue, L.; Liu, Y.; Zhang, T.; Li, Y.; et al. Morphological and Molecular Evidence Reveal Eight New Species of Gymnopus from Northeast China. J. Fungi 2022, 8, 349. [Google Scholar] [CrossRef]
- Mao, N.; Xu, Y.Y.; Zhao, T.Y.; Lv, J.C.; Fan, L. New Species of Mallocybe and Pseudosperma from North China. J. Fungi 2022, 8, 256. [Google Scholar] [CrossRef]
- Wu, F.; Tohtirjap, A.; Fan, L.F.; Zhou, L.W.; Alvarenga, R.L.M.; Gibertoni, T.B.; Dai, Y.C. Global diversity and updated phylogeny of Auricularia (Auriculariales, Basidiomycota). J. Fungi 2021, 7, 933. [Google Scholar] [CrossRef]
- Guan, Q.X.; Zhao, C.L. Taxonomy and phylogeny of the wood-inhabiting fungal genus Hyphoderma with descriptions of three new species from East Asia. J. Fungi 2021, 7, 308. [Google Scholar] [CrossRef]
- Wang, D.Q.; Zhao, C.L. Morphological and Phylogenetic Evidence for Recognition of Two New Species of Phanerochaete from East Asia. J. Fungi 2021, 7, 1063. [Google Scholar] [CrossRef]
- He, M.Q.; Zhao, R.L.; Hyde, K.D.; Begerow, D.; Kemler, M.; Yurkov, A.; McKenzie, E.H.C.; Raspe, O.; Kakishima, M.; Sanchez-Ramırez, S.; et al. Notes, outline and divergence times of Basidiomycota. Fungal Divers. 2019, 99, 105–367. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Specimen No. | GenBank Accession No. | References | |
|---|---|---|---|---|
| ITS | nLSU | |||
| Brevicellicium exile | H (Spirin 8370) | MT002322 | MT002338 | [43] |
| B. olivascens | KHL 8571 | HE963792 | HE963793 | [36] |
| Dextrinocystis calamicola | He 5700 | MK204534 | MK204547 | [38] |
| Fibrodontia alba | TNMF 24944 | NR153983 | NG060401 | [24] |
| F. brevidens | Wu 9807-16 | KC928276 | KC928277 | [44] |
| Litschauerella gladiola | He 3171 | MK204555 | MK204556 | [38] |
| Luellia cystidiata | JHP 09455 | MW371211 | Unpublished | |
| Porpomyces mucidus | Dai 12692 | KT157833 | KT157838 | [45] |
| P. submucidus | Cui 5183 | KT152143 | KT152145 | [45] |
| Scytinopogon pallescens | He 5192 | MK204553 | [38] | |
| S. havencampii | DED 8300 | KT253946 | KT253947 | [46] |
| Sistotremastrum guttuliferum | He 3338 | MK204540 | MK204552 | [38] |
| S. niveocremeum | CBS 42854 | MH857381 | MH868921 | [47] |
| S. suecicum | H (Miettinen14550) | MT075860 | MT002336 | [43] |
| Sphaerobasidium minutum | KHL 11714 | DQ873652 | DQ873653 | [48] |
| Subulicystidium brachysporum | KASL 1584b | MH041544 | MH041610 | [49] |
| S. cochleum | KHL 11200 | MN207036 | MN207024 | [50] |
| S. longisporum | Ordynets 00146 | MN207039 | MN207032 | [50] |
| S. meridense | Hjm 16400 | MH041538 | MH041604 | [49] |
| Trechispora amianthina | CBS 202.54 | MH857292 | [47] | |
| T. araneosa | KHL 8570 | AF347084 | [35] | |
| T. bambusicola | CLZhao 3305 | MW544022 | MW520172 | [33] |
| T. bispora | CBS 142.63 | MH858241 | [47] | |
| T. byssinella | UC 2023068 | KP814481 | Unpublished | |
| T. clancularis | FRDBI 4426619 | MW487976 | Unpublished | |
| T. cohaerens | HHB 19445 | MW740327 | Unpublished | |
| T. copiosa | AMO 453 | MN701018 | [27] | |
| T. confinis | KHL 11197 | AY463473 | AY586719 | [35] |
| T. daweishanensis | CLZhao 18255 | MW302338 | [34] | |
| T. echinospora | MA Fungi 82486 | JX392853 | [36] | |
| T. farinacea | MA Fungi 79474 | JX392855 | JX392856 | [36] |
| T. fimbriata | CLZhao 9006 | MW544025 | MW520175 | [33] |
| T. fissurata | CLZhao 4571 | MW544027 | [33] | |
| T. gelatinosa | AMO 1139 | MN701021 | [27] | |
| T. havencampii | DED 8300 | NR154418 | [46] | |
| T. hondurensis | HONDURAS 19-F016 | MT571523 | MT636540 | Unpublished |
| T. hymenocystis | KHL 8795 | AF347090 | [35] | |
| T. incisa | GB 0090648 | KU747095 | Unpublished | |
| T. invisitata | UC 2023088 | KP814425 | Unpublished | |
| T. kavinioides | KGN 981002 | AF347086 | [35] | |
| T. mollis | URM 85884 | MK514945 | [26] | |
| T. mollusca | CBS 43948 | MH856428 | [47] | |
| T. murina | CLZhao 11736 | OL615003 | Present study | |
| T. murina | CLZhao 11752 | OL615004 | OL615009 | Present study |
| T. nivea | MA Fungi 74044 | JX392832 | [36] | |
| T.odontioidea | CLZhao 17890 | ON417458 | Present study | |
| T.olivacea | CLZhao 17826 | ON417457 | Present study | |
| T. pallescens | FLOR 56186 | MK458766 | Unpublished | |
| T. papillosa | AMO 713 | MN701022 | [27] | |
| T. regularis | KHL 10881 | AF347087 | [35] | |
| T. rigida | URM 85754 | MT406381 | [26] | |
| T. stellulata | UC 2023096 | KP814450 | Unpublished | |
| T. stevensonii | MA Fungi 70669 | JX392841 | [36] | |
| T. subsphaerospora | KHL 8511 | AF347080 | [35] | |
| T. termitophila | AMO 1169 | MN701028 | [27] | |
| T. thelephora | 1820 AMV | KF937369 | [51] | |
| T. torrendii | URM 85886 | MK515148 | [26] | |
| T. xantha | CLZhao 17781 | MW302340 | [34] | |
| T. yunnanensis | CLZhao 215 | MN654923 | [31] | |
| Tubulicium raphidisporum | He 3191 | MK204537 | MK204545 | [38] |
| T. vermiferum | KHL 8714 | — | AY463477 | [35] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, K.; Zhao, C. A Molecular Systematics and Taxonomy Research on Trechispora (Hydnodontaceae, Trechisporales): Concentrating on Three New Trechispora Species from East Asia. J. Fungi 2022, 8, 1020. https://doi.org/10.3390/jof8101020
Luo K, Zhao C. A Molecular Systematics and Taxonomy Research on Trechispora (Hydnodontaceae, Trechisporales): Concentrating on Three New Trechispora Species from East Asia. Journal of Fungi. 2022; 8(10):1020. https://doi.org/10.3390/jof8101020
Chicago/Turabian StyleLuo, Kaiyue, and Changlin Zhao. 2022. "A Molecular Systematics and Taxonomy Research on Trechispora (Hydnodontaceae, Trechisporales): Concentrating on Three New Trechispora Species from East Asia" Journal of Fungi 8, no. 10: 1020. https://doi.org/10.3390/jof8101020