
Morphological and Phylogenetic Characterizations Reveal Five New Species of Astrothelium (Trypetheliales, Ascomycota) from China
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
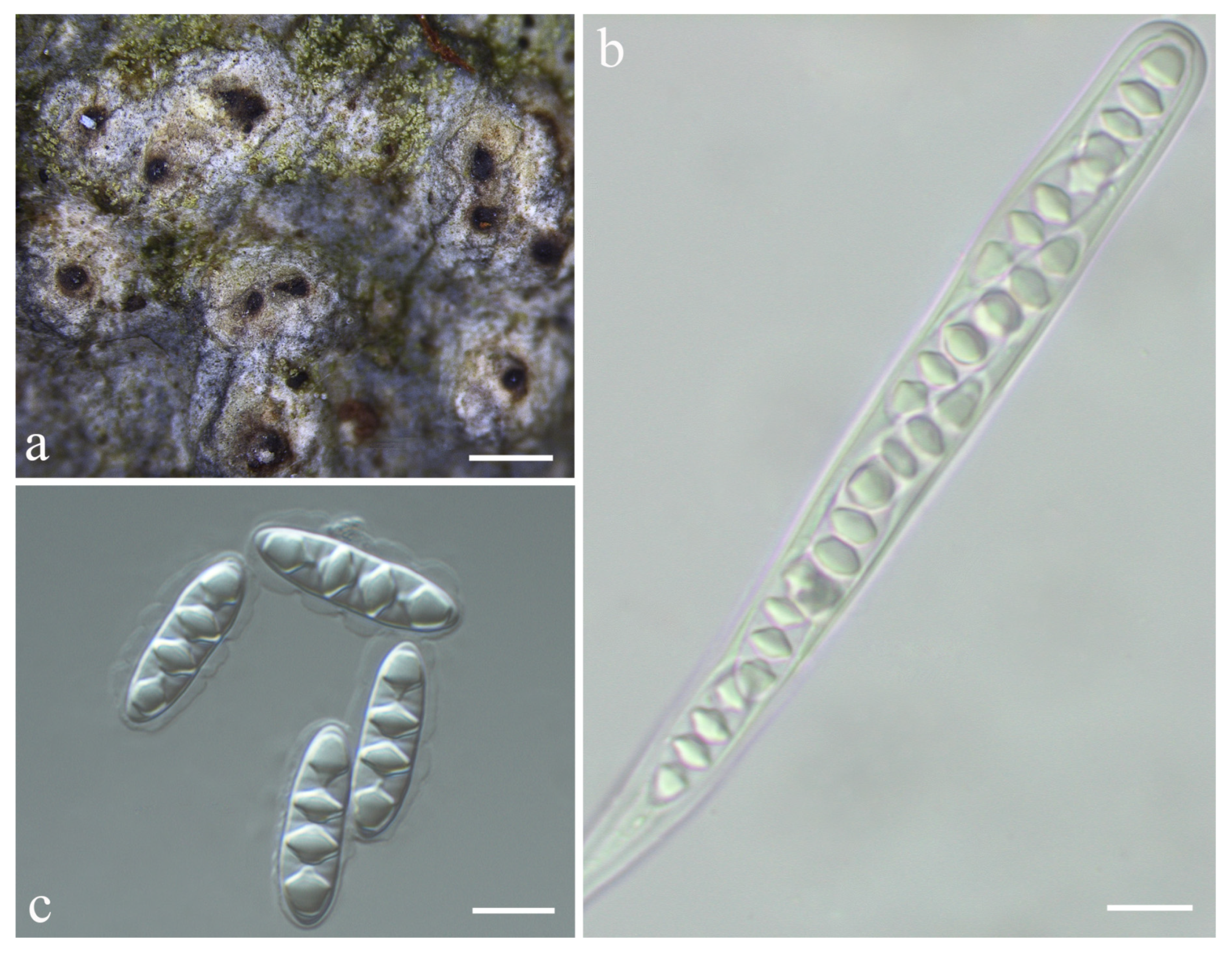{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Material Examined
2.2. Phenotypic Analysis
2.3. Chemical Analysis
2.4. Phylogenetic Analyses
2.4.1. DNA Extraction and PCR Amplification
2.4.2. Phylogenetic Analyses
3. Results
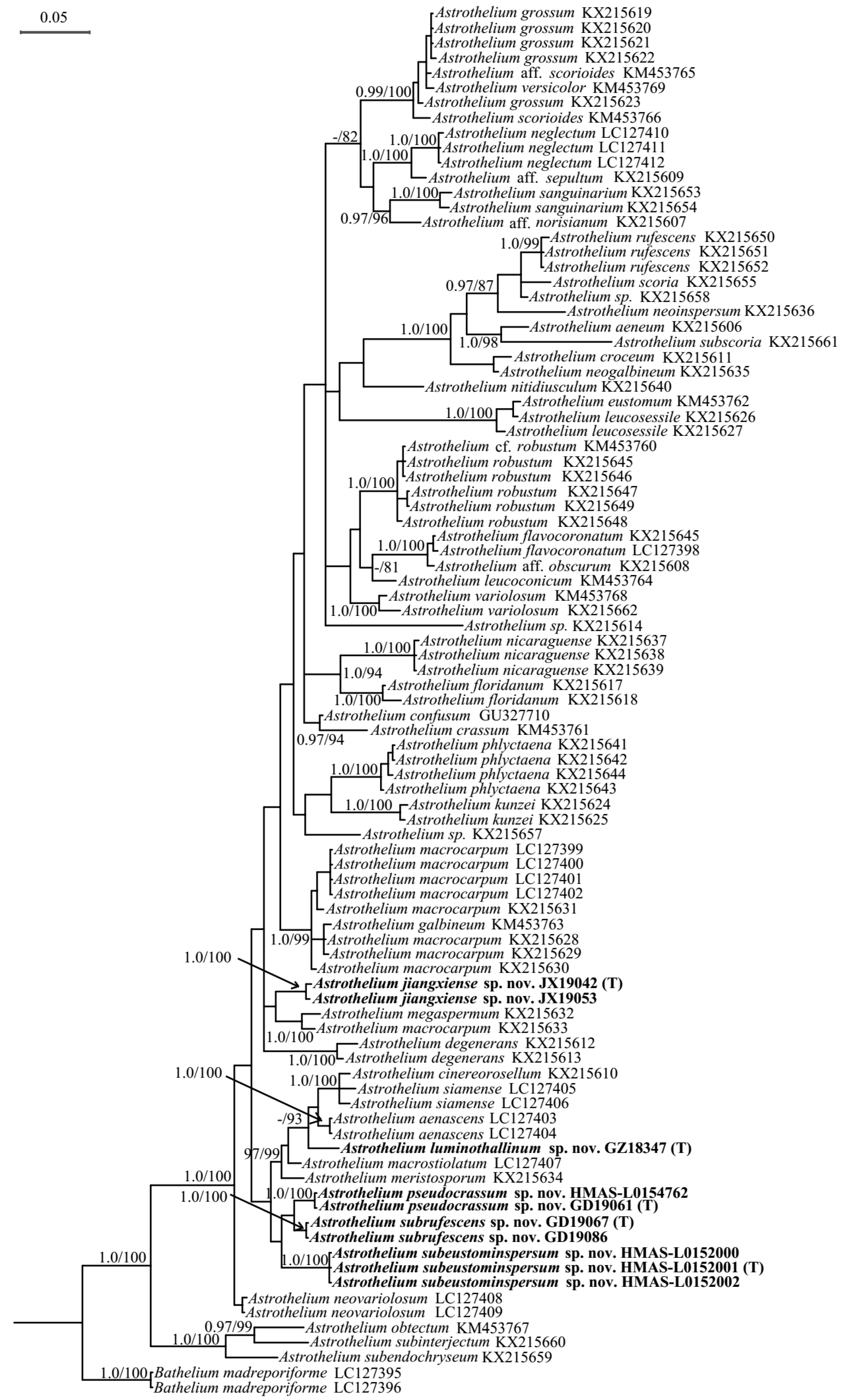
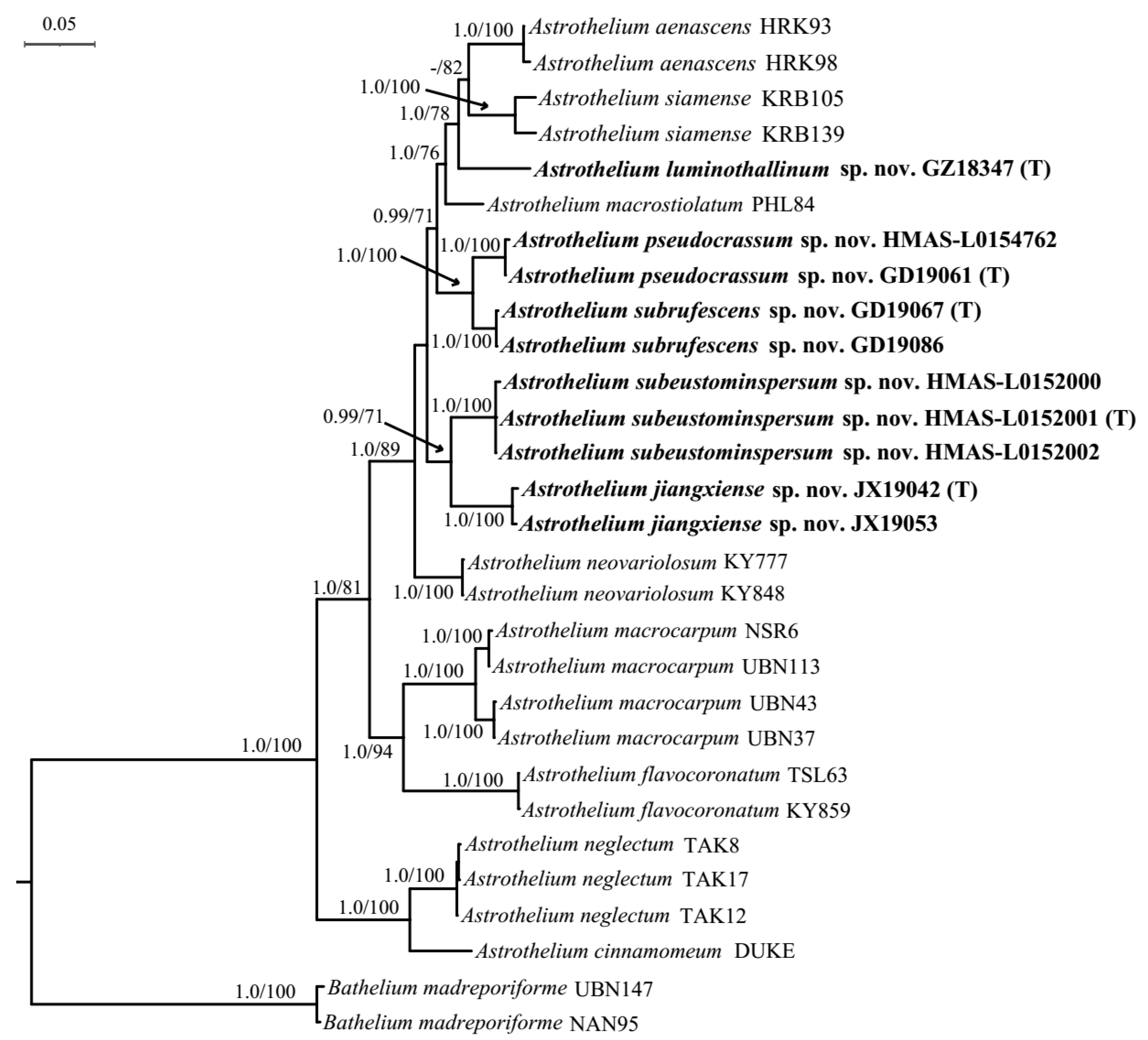
3.1. Phylogenetic Analyses
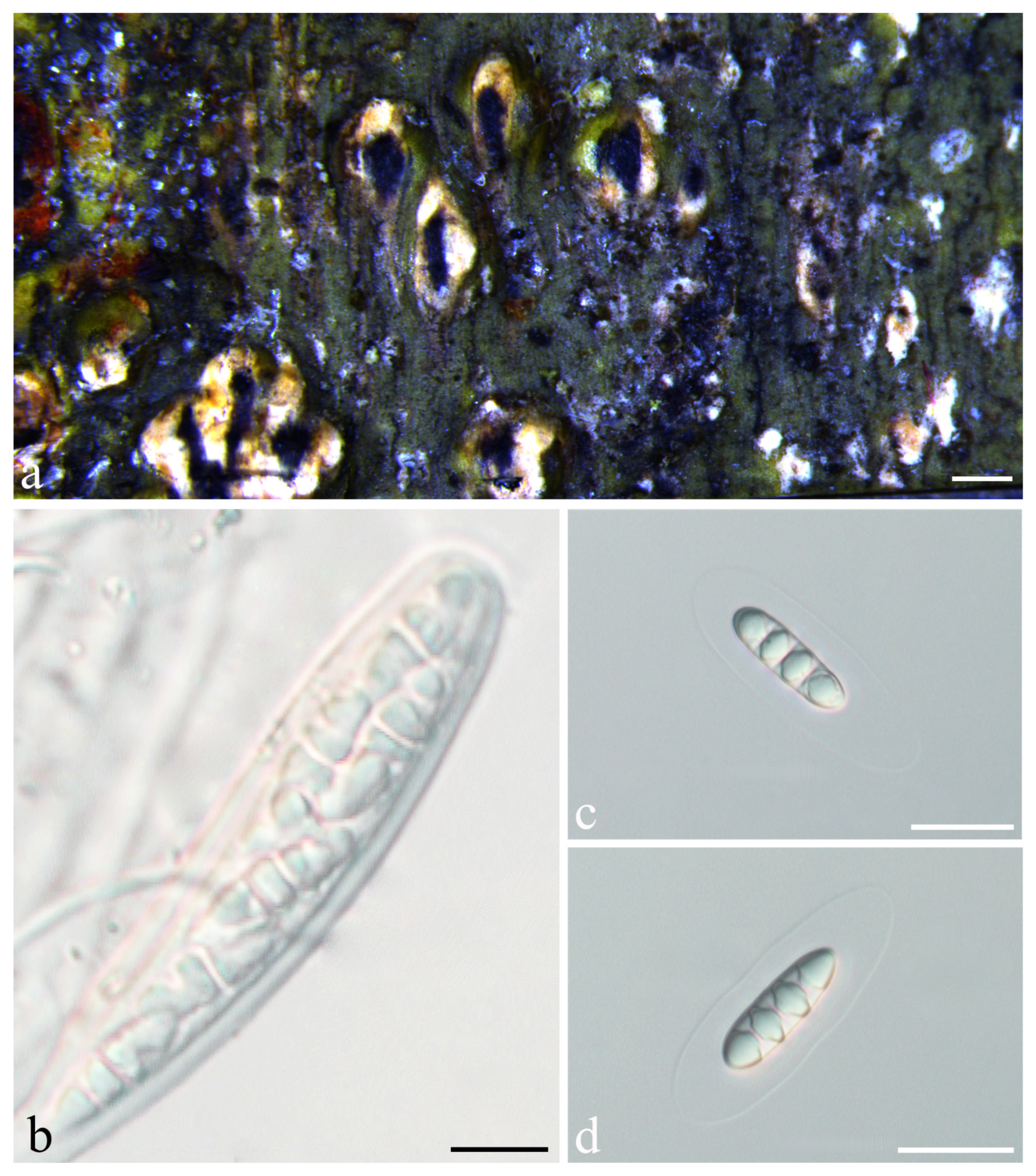
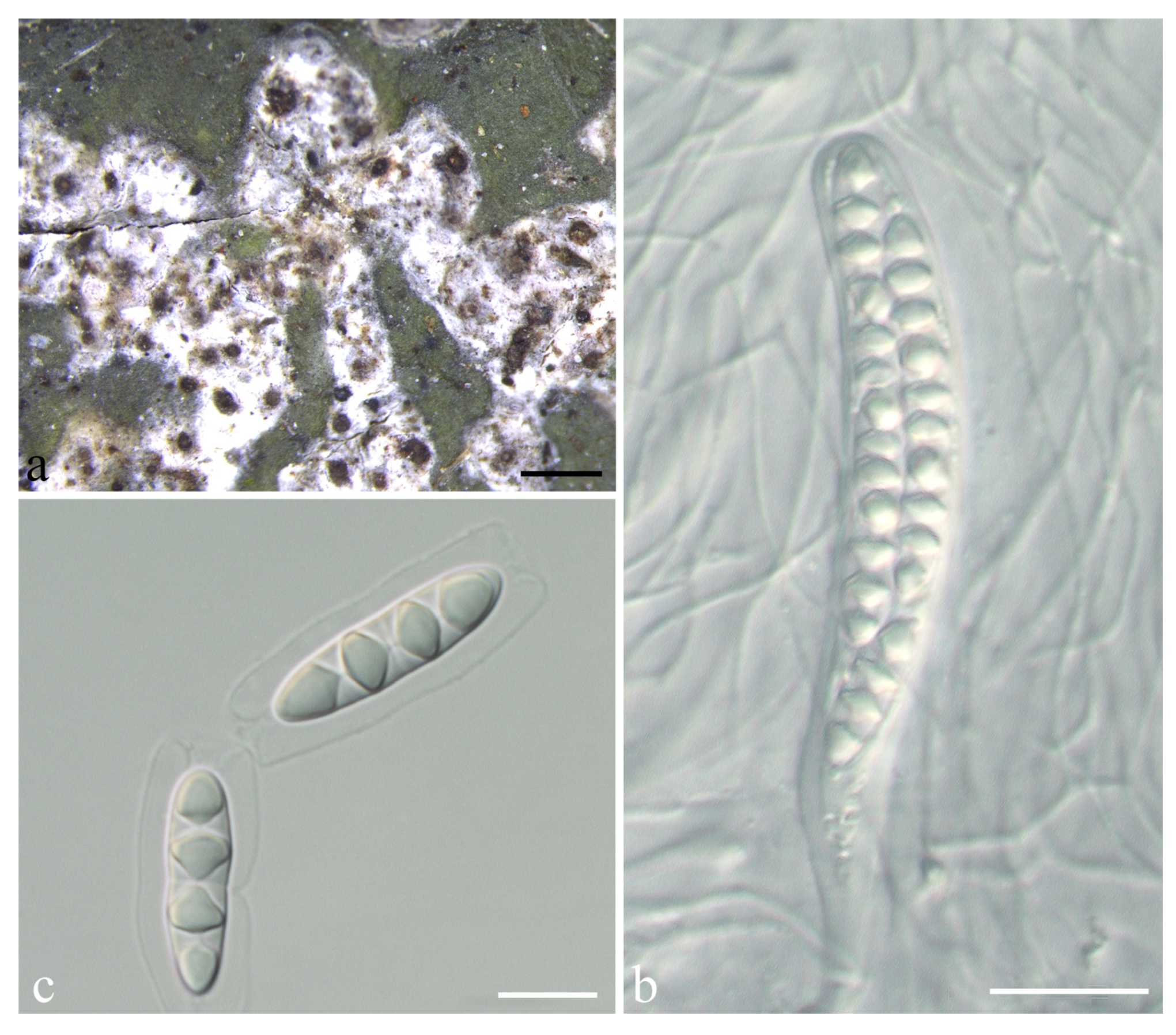
3.2. Taxonomy





4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.C.; Sarma, V.V.; Lücking, R.; Boonmee, S.; Bhat, J.D.; Liu, N.G.; et al. Refined families of Dothideomycetes: Orders and families incertae sedis in Dothideomycetes. Fungal Divers. 2020, 105, 17–318. [Google Scholar] [CrossRef]
- Hyde, K.D.; Jones, E.B.G.; Liu, J.K.; Ariyawansa, H.; Boehm, E.; Boonmee, S.; Braun, U.; Chomnunti, P.; Crous, P.W.; Dai, D.Q.; et al. Families of Dothideomycetes. Fungal Divers. 2013, 63, 1–313. [Google Scholar] [CrossRef]
- Nelsen, M.P.; Lücking, R.; Grube, M.; Mbatchou, J.S.; Muggia, L.; Plata, E.R.; Lumbsch, H.T. Unravelling the phylogenetic relationships of lichenised fungi in Dothideomyceta. Stud. Mycol. 2009, 64, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.C. Working Keys to the Lichen-Forming Fungi of Puerto Rico. In Tropical Lichen Workshop; Catholic University of Puerto Rico: Ponce, Puerto Rico, 1989. [Google Scholar]
- Harris, R.C. More Florida Lichens. In Including the 10ȼ Tour of the Pyrenolichens; The New York Botanical Garden: New York, NY, USA, 1995; pp. 1–182. [Google Scholar]
- Aptroot, A.; Lücking, R.; Sipman, H.J.M.; Umaña, L.; Chaves, J.L. Pyrenocarpous lichens with bitunicate asci. A first assessment of the lichen biodiversity inventory in Costa Rica. Bibl. Lichenol. 2008, 97, 1–162. [Google Scholar]
- Del Prado, R.; Schmitt, I.; Kautz, S.; Palice, Z.; Lücking, R.; Lumbsch, H.T. Molecular data place Trypetheliaceae in Dothideomycetes. Mycol. Res. 2006, 110, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Sweetwood, G.; Lücking, R.; Nelsen, M.P.; Aptroot, A. Ascospore ontogeny and discharge in megalosporous Trypetheliaceae and Graphidaceae (Ascomycota: Dothideomycetes and Lecanoromycetes) suggest phylogenetic relationships and ecological constraints. Lichenologist 2012, 44, 277–296. [Google Scholar] [CrossRef]
- Nelsen, M.P.; Lücking, R.; Aptroot, A.; Andrew, C.J.; Cáceres, M.; Plata, E.R.; Gueidan, C.; Silva Canêz, L.; Knight, A.; Ludwig, L.R.; et al. Elucidating phylogenetic relationships and genus-level classification within the fungal family Trypetheliaceae (Ascomycota: Dothideomycetes). Taxon 2014, 63, 974–992. [Google Scholar] [CrossRef]
- Lücking, R.; Nelsen, M.P.; Aptroot, A.; De Klee, R.B.; Bawingan, P.A.; Benatti, M.N.; Binh, N.Q.; Bungartz, F.; Cáceres, M.E.S.; Silva Canêz, L.; et al. A phylogenetic framework for reassessing generic concepts and species delimitation in the lichenized family Trypetheliaceae (Ascomycota: Dothideomycetes). Lichenologist 2016, 48, 739–762. [Google Scholar] [CrossRef]
- Lücking, R.; Nelsen, M.P.; Aptroot, A.; Benatti, M.N.; Binh, N.Q.; Gueidan, C.; Guitiérrez, M.C.; Jungbluth, P.; Lumbsch, H.T.; Marcelli, M.P.; et al. A pot-pourri of new species of Trypetheliaceae resulting from molecular phylogenetic studies. Lichenologist 2016, 48, 639–660. [Google Scholar] [CrossRef]
- Aptroot, A.; Lücking, R. A revisionary synopsis of the Trypetheliaceae (Ascomycota: Trypetheliales). Lichenologist 2016, 48, 763–982. [Google Scholar] [CrossRef]
- Lücking, R.; Hodkinson, B.P.; Leavitt, S.D. The 2016 classification of lichenized fungi in the Ascomycota & Basidiomycota—Approaching one thousand genera. Bryologist 2017, 119, 361–416. [Google Scholar]
- Cáceres, M.E.S.; Aptroot, A. Lichens from the Brazilian Amazon, with special reference to the genus Astrothelium. Bryologist 2017, 120, 165–181. [Google Scholar] [CrossRef]
- Aptroot, A.; Weerakoon, G. Three new species and ten new records of Trypetheliaceae (Ascomycota) from Sri Lanka. Cryptogam. Mycol. 2018, 39, 373–377. [Google Scholar] [CrossRef]
- Aptroot, A.; Sipman, H.J.M.; Barreto, F.M.O.; Nunes, A.D.; Cáceres, M.E.S. Ten new species and 34 new country records of Trypetheliaceae. Lichenologist 2019, 51, 27–43. [Google Scholar] [CrossRef]
- Jiang, S.H.; Zhang, C.; Yao, Z.T.; Liu, H.J. A species new to science and a new Chinese record of Astrothelium (Typetheliaceae, Ascomycota). Mycosystema 2021, 40, 31–39. [Google Scholar]
- Harris, R.C. The family Trypetheliaceae (Loculoascomycetes: Lichenized Melanommatales) in Amazonian Brazil. Suppl. Acta Amaz. 1984, 14, 55–80. [Google Scholar] [CrossRef]
- Awasthi, D.D. A key to the microlichens of India, Nepal and Sri Lanka. Bibl. Lichenol. 1991, 40, 1–340. [Google Scholar]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Dictionary of the Fungi, 10th ed.; Wallingford, CAB International: Wallingford, UK, 2008. [Google Scholar]
- Aptroot, A. Trypetheliaceae. Flora Aust. 2009, 57, 535–552. [Google Scholar]
- Medina, E.S.; Aptroot, A.; Lücking, R. Aspidothelium silverstonei and Astrothelium fuscosporum, two new corticolous lichen species from Colombia. Cryptogam. Mycol. 2017, 38, 253–258. [Google Scholar] [CrossRef]
- Aptroot, A.; Seaward, M.R.D. Annotated checklist of Hongkong Lichens. Trop. Bryol. 1999, 17, 57–101. [Google Scholar]
- Zahlbruckner, A. Flechten der Insel Formosa in Feddes, Lichenes in Feddes, Repertorium sp. Feddes Repert. 1933, 31, 194–224. [Google Scholar] [CrossRef]
- Elix, J.A. A Catalogue of Standardized Thin-Layer Chromatographic Data and Biosynthetic Relationships for Lichen Substances, 3rd ed.; Australian National University: Canberra, Australia, 2014. [Google Scholar]
- Orange, A.; James, P.W.; White, F.J. Microchemical Methods for the Identification of Lichens; British Lichen Society: London, UK, 2010. [Google Scholar]
- Rogers, S.O.; Bendich, A.J. Extraction of DNA from Plant Tissues. In Plant Molecular Biology Manual; Gelvin, S.B., Schilperoort, R.A., Verma, D.P.S., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1988; pp. 1–10. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.J.; Taylor, J.L. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. A Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Luangsuphabool, T.; Lumbsch, H.T.; Aptroot, A.; Piapukiew, J.; Sangvichien, E. Five new species and one new record of Astrothelium (Trypetheliaceae, Ascomycota) from Thailand. Lichenologist 2016, 48, 727–737. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.H.; Lücking, R.; Liu, H.J.; Wei, X.L.; Xavier-Leite, A.B.; Portilla, C.V.; Ren, Q.; Wei, J.C. Twelve new species reveal cryptic diversification in foliicolous lichens of Strigula s.lat. (Strigulales, Ascomycota). J. Fungi 2022, 8, 2. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Kelly, L.J.; Hollingsworth, P.M.; Coppins, B.J.; Ellis, C.J.; Harrold, P.; Tosh, J.; Yahr, R. DNA barcoding of lichenized fungi demonstrates high identification success in a floristic context. New Phytol. 2011, 191, 288–300. [Google Scholar] [CrossRef]
- Aptroot, A. World key to the species of Pyrenulaceae and Trypetheliaceae. Arch. Lichenol. 2021, 29, 1–91. [Google Scholar]
- Flakus, A.; Kukwa, M.; Aptroot, A. Trypetheliaceae of Bolivia: An updated checklist with descriptions of twenty-four new species. Lichenologist 2016, 48, 661–692. [Google Scholar] [CrossRef]
- Aptroot, A.; Ertz, D.; Etayo, J.A.S.; Gueidan, C.; Mercado-Díaz, J.A.; Schumm, F.; Weerakoon, G. Forty-six new species of Trypetheliaceae from the tropics. Lichenologist 2016, 48, 609–638. [Google Scholar] [CrossRef]
- Junior, I.O.; Aptroot, A.; Marcela, E.S.C. Lichens from Monte Pascoal, Bahia, Brazil, with some new pyrenocarpous species and a key to the Pyrenula species from Brazil. Bryologist 2021, 124, 552–568. [Google Scholar] [CrossRef]
- Zhu, H. The tropical forests of Southern China and conservation of biodiversity. Bot. Rev. 2017, 83, 87–105. [Google Scholar] [CrossRef]
- Aptroot, A.; Sparrius, L.B. New microlichens from Taiwan. Fungal Divers. 2003, 14, 1–50. [Google Scholar]
- Aptroot, A. Pyrenocarpous lichens and related nonlichenized ascomycetes from Taiwan. J. Hattort. Bot. Lab. 2003, 93, 155–173. [Google Scholar]
- Aptroot, A.; Ferraro, L.I.; Lai, M.J.; Sipman, H.J.M.; Sparrius, L.B. Foliicolous lichens and their lichenicolous ascomycetes from Yunnan and Taiwan. Mycotaxon 2003, 88, 41–47. [Google Scholar]
- Aptroot, A.; Sipman, H.J.M. New Hong Kong lichens, ascomycetes and lichenicolous fungi. J. Hattort. Bot. Lab. 2001, 91, 317–343. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, S.-H.; Zhang, C.; Xue, X.-D.; Aptroot, A.; Wei, J.-C.; Wei, X.-L. Morphological and Phylogenetic Characterizations Reveal Five New Species of Astrothelium (Trypetheliales, Ascomycota) from China. J. Fungi 2022, 8, 994. https://doi.org/10.3390/jof8100994
Jiang S-H, Zhang C, Xue X-D, Aptroot A, Wei J-C, Wei X-L. Morphological and Phylogenetic Characterizations Reveal Five New Species of Astrothelium (Trypetheliales, Ascomycota) from China. Journal of Fungi. 2022; 8(10):994. https://doi.org/10.3390/jof8100994
Chicago/Turabian StyleJiang, Shu-Hua, Chao Zhang, Xian-Dong Xue, André Aptroot, Jiang-Chun Wei, and Xin-Li Wei. 2022. "Morphological and Phylogenetic Characterizations Reveal Five New Species of Astrothelium (Trypetheliales, Ascomycota) from China" Journal of Fungi 8, no. 10: 994. https://doi.org/10.3390/jof8100994