
Access to Anti-Biofilm Compounds from Endolichenic Fungi Using a Bioguided Networking Screening
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. General Experimental Procedures
2.3. Fungi and Culturing Conditions
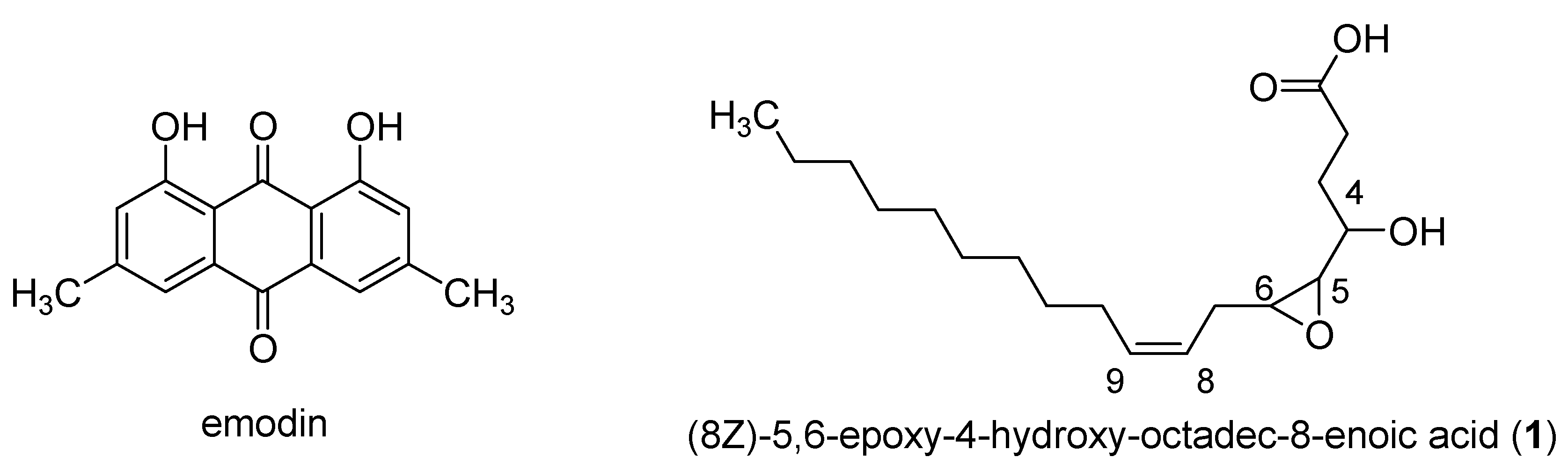
2.4. Extraction and Isolation of Compound 1
2.5. Anti-Maturation and Anti-Biofilm Assays
2.6. Statistical Analysis
2.7. LC-HRMS Analyses
2.8. Data Treatment
3. Results
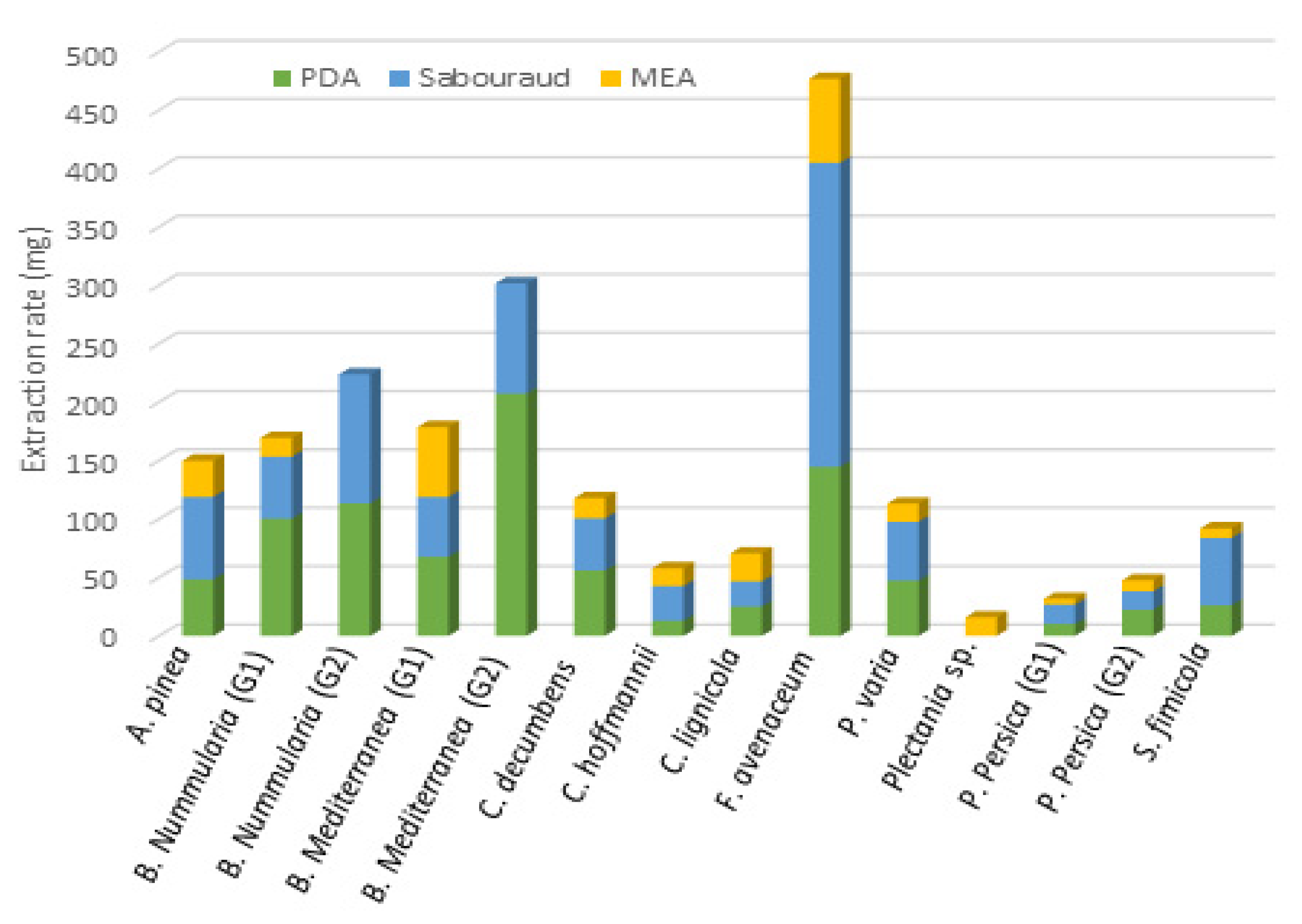
3.1. Extracts Library
3.2. Anti-Biofilm Screening Results
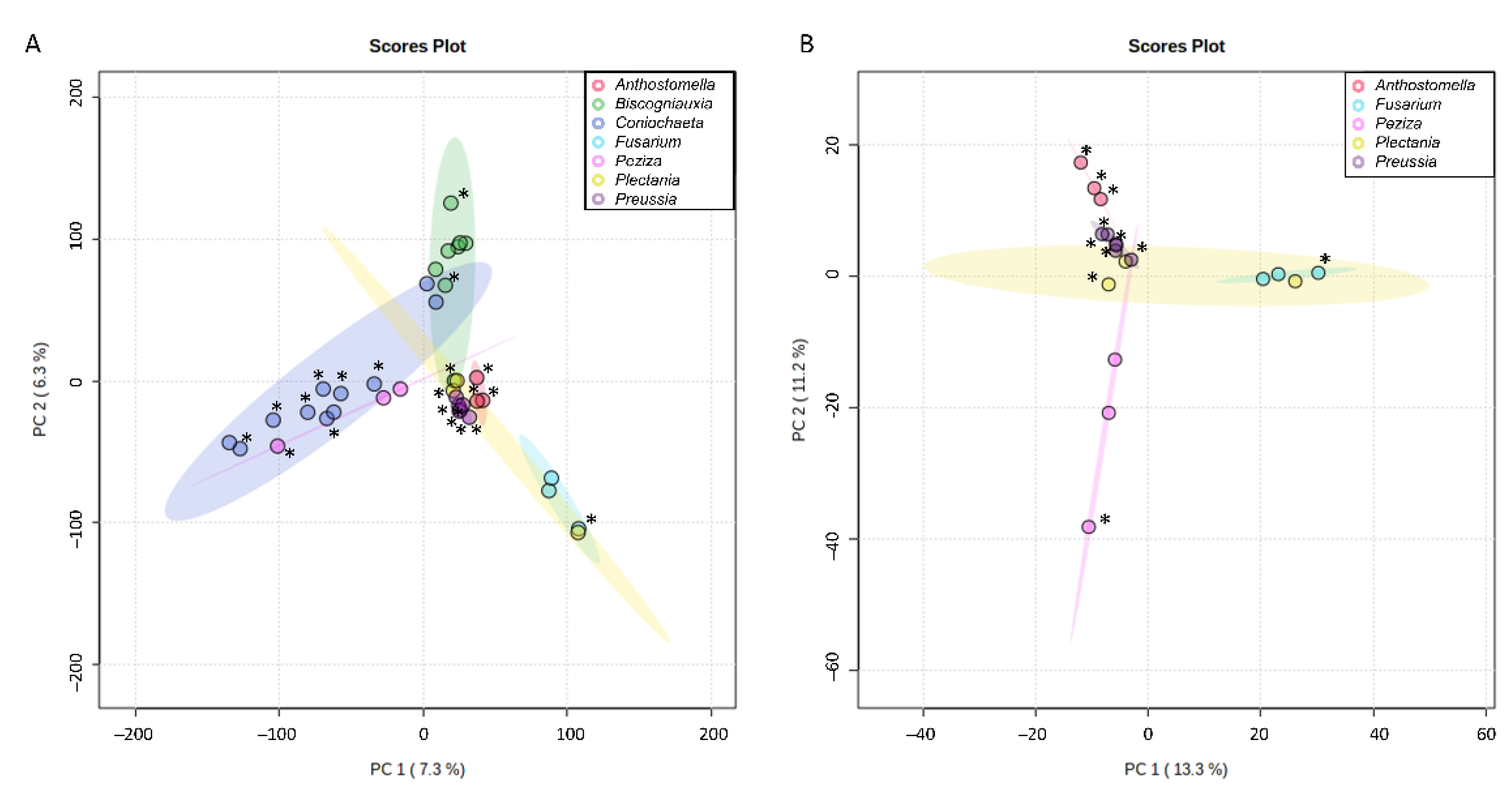
3.3. First Insight into Endolichenic Fungi Metabolomes
3.4. Research for Active Compounds by Biochemometrics and Molecular Networking
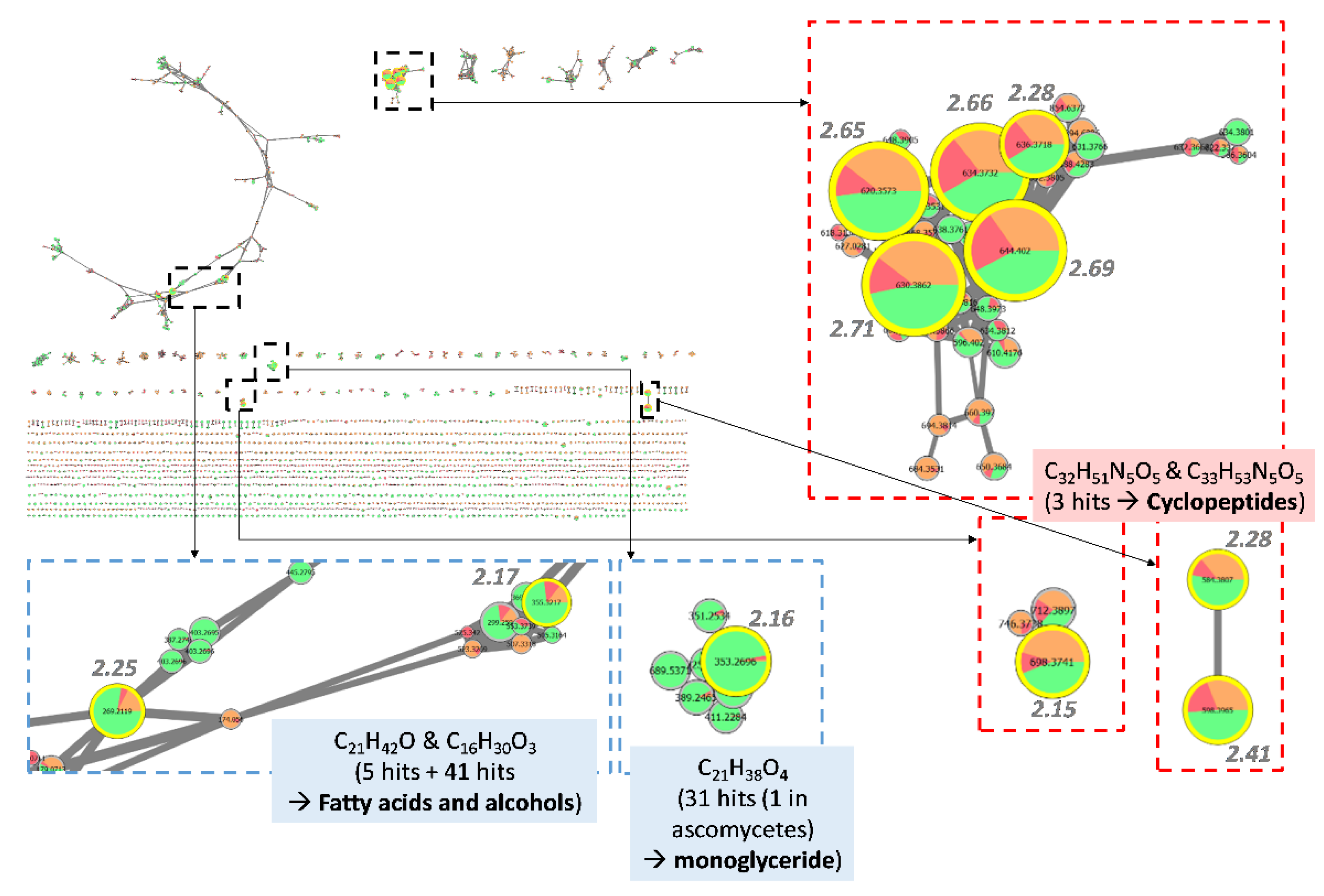
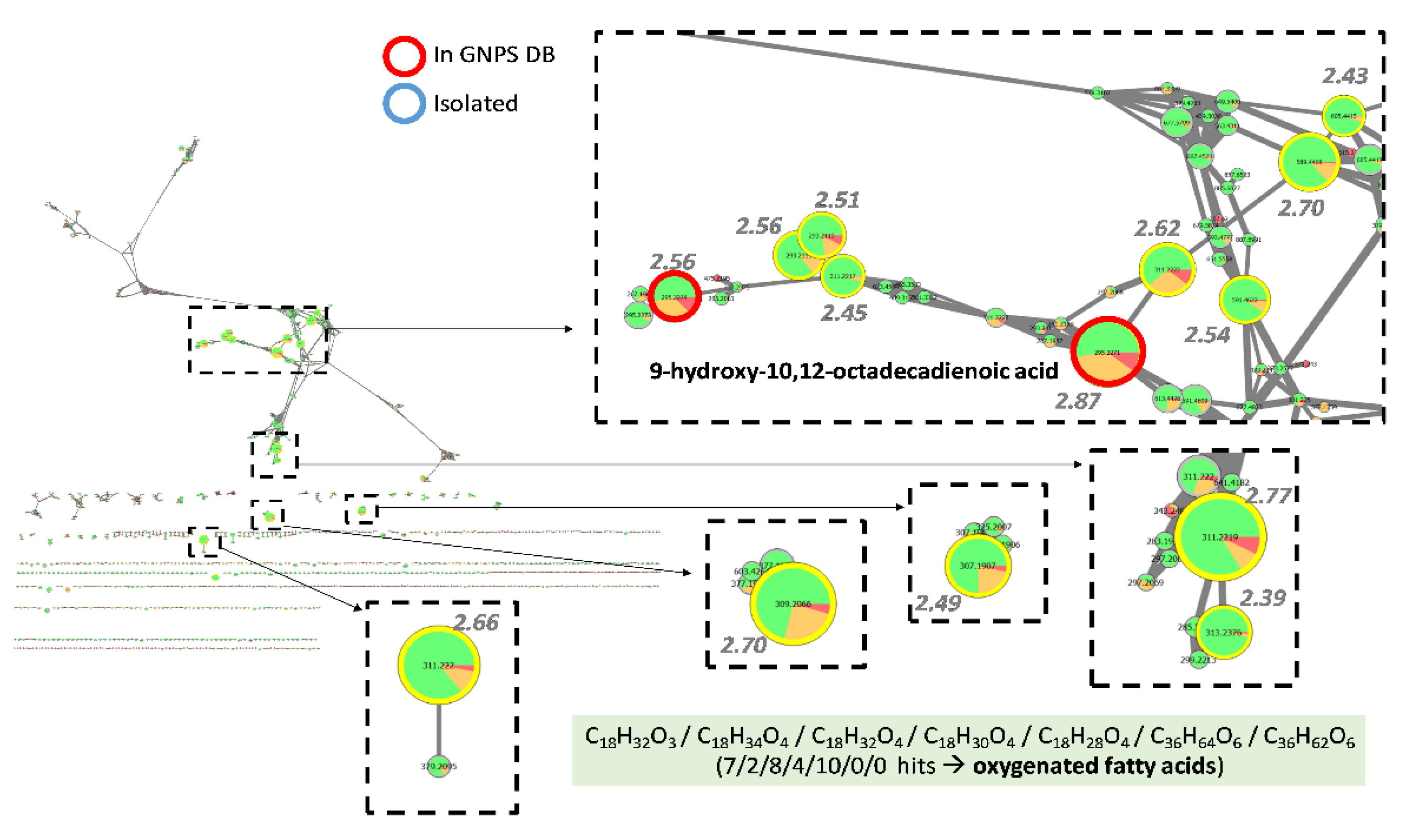
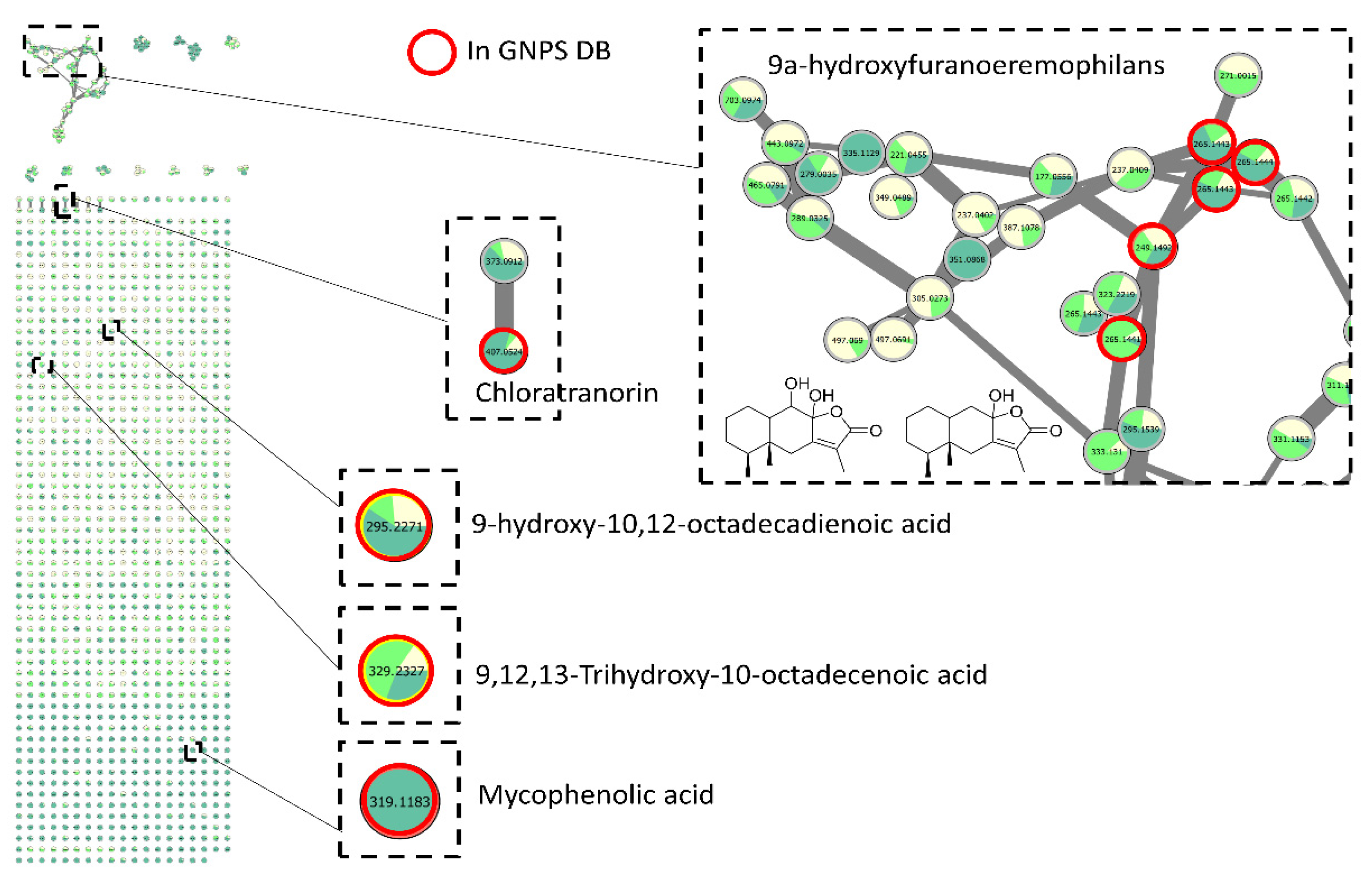
3.4.1. Investigation of Extracts from Coniochaeta Genus
3.4.2. Investigation of Extracts from Biscognauxia Genus
3.4.3. Investigation of Extracts from Preussia Genus
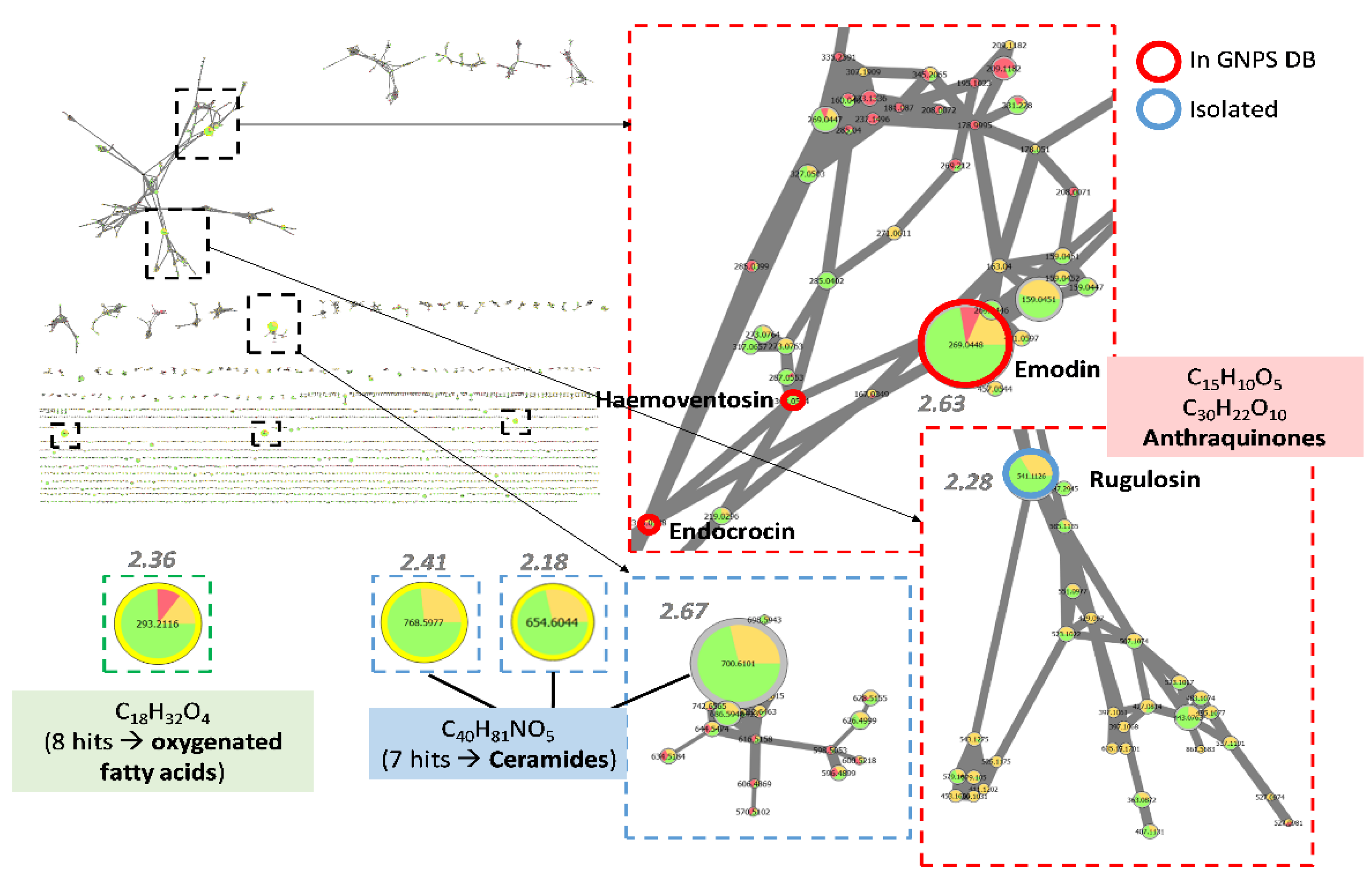
3.4.4. Dereplication of Other Active Extracts from Anthostomella
3.5. Isolation of Active Compounds
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kellogg, J.J.; Raja, H.A. Endolichenic Fungi: A New Source of Rich Bioactive Secondary Metabolites on the Horizon. Phytochem. Rev. 2017, 16, 271–293. [Google Scholar] [CrossRef]
- Singh, B.N.; Upreti, D.K.; Gupta, V.K.; Dai, X.-F.; Jiang, Y. Endolichenic Fungi: A Hidden Reservoir of Next Generation Biopharmaceuticals. Trends Biotechnol. 2017, 35, 808–813. [Google Scholar] [CrossRef]
- Suryanarayanan, T.S.; Thirunavukkarasu, N. Endolichenic Fungi: The Lesser Known Fungal Associates of Lichens. Mycology 2017, 8, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Sardi, J.C.O.; Scorzoni, L.; Bernardi, T.; Fusco-Almeida, A.M.; Mendes Giannini, M.J.S. Candida Species: Current Epidemiology, Pathogenicity, Biofilm Formation, Natural Antifungal Products and New Therapeutic Options. J. Med. Microbiol. 2013, 62, 10–24. [Google Scholar] [CrossRef]
- Bernard, C.; Renaudeau, N.; Mollichella, M.-L.; Quellard, N.; Girardot, M.; Imbert, C. Cutibacterium acnes Protects Candida albicans from the Effect of Micafungin in Biofilms. Int. J. Antimicrob. Agents 2018, 52, 942–946. [Google Scholar] [CrossRef]
- Gupta, P.; Sarkar, S.; Das, B.; Bhattacharjee, S.; Tribedi, P. Biofilm, Pathogenesis and Prevention—A Journey to Break the Wall: A Review. Arch. Microbiol. 2016, 198, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Montelongo-Jauregui, D.; Srinivasan, A.; Ramasubramanian, A.K.; Lopez-Ribot, J.L. An in Vitro Model for Oral Mixed Biofilms of Candida albicans and Streptococcus gordonii in Synthetic Saliva. Front. Microbiol. 2016, 7, 1821. [Google Scholar] [CrossRef]
- Cocuaud, C.; Rodier, M.-H.; Daniault, G.; Imbert, C. Anti-Metabolic Activity of Caspofungin against Candida albicans and Candida parapsilosis Biofilms. J. Antimicrob. Chemother. 2005, 56, 507–512. [Google Scholar] [CrossRef]
- Kuhn, D.M.; George, T.; Chandra, J.; Mukherjee, P.K.; Ghannoum, M.A. Antifungal Susceptibility of Candida Biofilms: Unique Efficacy of Amphotericin B Lipid Formulations and Echinocandins. Antimicrob. Agents Chemother. 2002, 46, 1773–1780. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Long, L.; Kim, H.G.; Ghannoum, M.A. Amphotericin B Lipid Complex Is Efficacious in the Treatment of Candida albicans Biofilms Using a Model of Catheter-Associated Candida Biofilms. Int. J. Antimicrob. Agents 2009, 33, 149–153. [Google Scholar] [CrossRef]
- Toulet, D.; Debarre, C.; Imbert, C. Could Liposomal Amphotericin B (L-AMB) Lock Solutions Be Useful to Inhibit Candida Spp. Biofilms on Silicone Biomaterials? J. Antimicrob. Chemother. 2012, 67, 430–432. [Google Scholar] [CrossRef] [PubMed]
- Girardot, M.; Imbert, C. Natural Sources as Innovative Solutions Against Fungal Biofilms. In Fungal Biofilms and Related Infections; Imbert, C., Ed.; Springer: Cham, Switzerland, 2016; Volume 931, pp. 105–125. ISBN 978-3-319-42359-3. [Google Scholar]
- You, J.; Du, L.; King, J.B.; Hall, B.E.; Cichewicz, R.H. Small-Molecule Suppressors of Candida albicans Biofilm Formation Synergistically Enhance the Antifungal Activity of Amphotericin B against Clinical Candida Isolates. ACS Chem. Biol. 2013, 8, 840–848. [Google Scholar] [CrossRef] [PubMed]
- Millot, M.; Girardot, M.; Dutreix, L.; Mambu, L.; Imbert, C. Antifungal and Anti-Biofilm Activities of Acetone Lichen Extracts against Candida albicans. Molecules 2017, 22, 651. [Google Scholar] [CrossRef] [PubMed]
- Girardot, M.; Millot, M.; Hamion, G.; Billard, J.-L.; Juin, C.; Ntoutoume, G.M.A.N.; Sol, V.; Mambu, L.; Imbert, C. Lichen Polyphenolic Compounds for the Eradication of Candida albicans Biofilms. Front. Cell. Infect. Microbiol. 2021, 11, 698883. [Google Scholar] [CrossRef]
- Lagarde, A.; Jargeat, P.; Roy, M.; Girardot, M.; Imbert, C.; Millot, M.; Mambu, L. Fungal Communities Associated with Evernia prunastri, Ramalina fastigiata and Pleurosticta acetabulum: Three Epiphytic Lichens Potentially Active against Candida Biofilms. Microbiol. Res. 2018, 211, 1–12. [Google Scholar] [CrossRef]
- Dalleau, S.; Cateau, E.; Bergès, T.; Berjeaud, J.-M.; Imbert, C. In Vitro Activity of Terpenes against Candida Biofilms. Int. J. Antimicrob. Agents 2008, 31, 572–576. [Google Scholar] [CrossRef]
- Xia, J.; Psychogios, N.; Young, N.; Wishart, D.S. MetaboAnalyst: A Web Server for Metabolomic Data Analysis and Interpretation. Nucleic Acids Res. 2009, 37, W652–W660. [Google Scholar] [CrossRef]
- Ory, L.; Nazih, E.-H.; Daoud, S.; Mocquard, J.; Bourjot, M.; Margueritte, L.; Delsuc, M.-A.; Bard, J.-M.; Pouchus, Y.F.; Bertrand, S.; et al. Targeting Bioactive Compounds in Natural Extracts—Development of a Comprehensive Workflow Combining Chemical and Biological Data. Anal. Chim. Acta 2019, 1070, 29–42. [Google Scholar] [CrossRef]
- Lagarde, A.; Millot, M.; Pinon, A.; Liagre, B.; Girardot, M.; Imbert, C.; Ouk, T.S.; Jargeat, P.; Mambu, L. Antiproliferative and Antibiofilm Potentials of Endolichenic Fungi Associated with the Lichen Nephroma laevigatum. J. Appl. Microbiol. 2019, 126, 1044–1058. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and Community Curation of Mass Spectrometry Data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [Green Version]
- Allard, P.-M.; Péresse, T.; Bisson, J.; Gindro, K.; Marcourt, L.; Pham, V.C.; Roussi, F.; Litaudon, M.; Wolfender, J.-L. Integration of Molecular Networking and In-Silico MS/MS Fragmentation for Natural Products Dereplication. Anal. Chem. 2016, 88, 3317–3323. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Sanchez, L.M.; Rath, C.M.; Liu, X.; Boudreau, P.D.; Bruns, N.; Glukhov, E.; Wodtke, A.; de Felicio, R.; Fenner, A.; et al. Molecular Networking as a Dereplication Strategy. J. Nat. Prod. 2013, 76, 1686–1699. [Google Scholar] [CrossRef] [PubMed]
- Janeczko, M.; Masłyk, M.; Kubiński, K.; Golczyk, H. Emodin, a Natural Inhibitor of Protein Kinase CK2, Suppresses Growth, Hyphal Development, and Biofilm Formation of Candida albicans: The Effects of Emodin on Candida albicans. Yeast 2017, 34, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Kurobane, I.; Vining, L.C.; Mcinnes, A.G. Biosynthetic Relationships among the Secalonic Acids: Isolation of Emodin, Endocrocin and Secalonic Acids from Pyrenochaeta terrestris and Aspergillus aculeatus. J. Antibiot. 1979, 32, 1256–1266. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-J.; Lin, Y.-C.; Yao, J.-H.; Vrijmoed, L.L.P.; Jones, E.B.G. Two New Metabolites from the Mangrove Endophytic Fungus No. 2524. J. Asian Nat. Prod. Res. 2004, 6, 185–191. [Google Scholar] [CrossRef]
- Nollmann, F.I.; Dauth, C.; Mulley, G.; Kegler, C.; Kaiser, M.; Waterfield, N.R.; Bode, H.B. Insect-Specific Production of New GameXPeptides in Photorhabdus luminescens TTO1, Widespread Natural Products in Entomopathogenic Bacteria. ChemBioChem 2015, 16, 205–208. [Google Scholar] [CrossRef]
- Talontsi, F.M.; Facey, P.; Tatong, M.D.K.; Tofazzal Islam, M.; Frauendorf, H.; Draeger, S.; Von Tiedemann, A.; Laatsch, H. Zoosporicidal Metabolites from an Endophytic Fungus Cryptosporiopsis Sp. of Zanthoxylum leprieurii. Phytochemistry 2012, 83, 87–94. [Google Scholar] [CrossRef]
- Bertrand, S.; Guitton, Y.; Roullier, C. Successes and Pitfalls in Automated Dereplication Strategy Using Liquid Chromatography Coupled to Mass Spectrometry Data: A CASMI 2016 Experience. Phytochem. Lett. 2017, 21, 297–305. [Google Scholar] [CrossRef]
- Bernart, M.W.; Whatley, G.G.; Gerwick, W.H. Unprecedented Oxylipins from the Marine Green Alga Acrosiphonia coalita. J. Nat. Prod. 1993, 56, 245–249. [Google Scholar]
- Yang, Z.; Piironen, V.; Lampi, A.-M. Epoxy and Hydroxy Fatty Acids as Non-Volatile Lipid Oxidation Products in Oat. Food Chem. 2019, 295, 82–93. [Google Scholar] [CrossRef]
- Morrison, E.; Kosiak, B.; Ritieni, A.; Aastveit, A.H.; Uhlig, S.; Bernhoft, A. Mycotoxin Production by Fusarium avenaceum Strains Isolated from Norwegian Grain and the Cytotoxicity of Rice Culture Extracts to Porcine Kidney Epithelial Cells. J. Agric. Food Chem. 2002, 50, 3070–3075. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, J.; Giese, H. Influence of Carbohydrates on Secondary Metabolism in Fusarium avenaceum. Toxins 2013, 5, 1655–1663. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Strobel, G.A.; Feng, T.; Ren, H.; Mends, M.T.; Zhou, Z.; Geary, B. An Endophytic Coniochaeta velutina Producing Broad Spectrum Antimycotics. J. Microbiol. 2015, 53, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Bouhet, J.C.; Pham Van Chuong, P.; Toma, F.; Kirszenbaum, M.; Fromageot, P. Isolation and Characterization of Luteoskyrin and Rugulosin, Two Hepatotoxic Anthraquinonoids from Penicillium islandicum Sopp. and Penicillium rugulosum Thom. J. Agric. Food Chem. 1976, 24, 964–972. [Google Scholar] [CrossRef]
- Hsu, S.-C.; Chung, J.-G. Anticancer Potential of Emodin. BioMedicine 2012, 2, 108–116. [Google Scholar] [CrossRef]
- Jiang, C.; Ge, J.; He, B.; Zeng, B. Glycosphingolipids in Filamentous Fungi: Biological Roles and Potential Applications in Cosmetics and Health Foods. Front. Microbiol. 2021, 12, 690211. [Google Scholar] [CrossRef]
- Vieira, E.; Xisto, M.; Pele, M.; Alviano, D.; Alviano, C.; Barreto-Bergter, E.; De Campos-Takaki, G. Monohexosylceramides from Rhizopus Species Isolated from Brazilian Caatinga: Chemical Characterization and Evaluation of Their Anti-Biofilm and Antibacterial Activities. Molecules 2018, 23, 1331. [Google Scholar] [CrossRef]
- McEvoy, K.; Normile, T.G.; Poeta, M.D. Antifungal Drug Development: Targeting the Fungal Sphingolipid Pathway. J. Fungi 2020, 6, 142. [Google Scholar] [CrossRef]
- Wu, B.; Wiese, J.; Schmaljohann, R.; Imhoff, J. Biscogniauxone, a New Isopyrrolonaphthoquinone Compound from the Fungus Biscogniauxia mediterranea Isolated from Deep-Sea Sediments. Mar. Drugs 2016, 14, 204. [Google Scholar] [CrossRef]
- Cheng, M.-J.; Wu, M.-D.; Yanai, H.; Su, Y.-S.; Chen, I.-S.; Yuan, G.-F.; Hsieh, S.-Y.; Chen, J.-J. Secondary Metabolites from the Endophytic Fungus Biscogniauxia formosana and Their Antimycobacterial Activity. Phytochem. Lett. 2012, 5, 467–472. [Google Scholar] [CrossRef]
- Pomilio, A.; Battista, M.; Vitale, A. Naturally-Occurring Cyclopeptides: Structures and Bioactivity. Curr. Org. Chem. 2006, 10, 2075–2121. [Google Scholar] [CrossRef]
- Wu, W.; Dai, H.; Bao, L.; Ren, B.; Lu, J.; Luo, Y.; Guo, L.; Zhang, L.; Liu, H. Isolation and Structural Elucidation of Proline-Containing Cyclopentapeptides from an Endolichenic Xylaria Sp. J. Nat. Prod. 2011, 74, 1303–1308. [Google Scholar] [CrossRef] [PubMed]
- Hüttel, W. Echinocandins: Structural Diversity, Biosynthesis, and Development of Antimycotics. Appl. Microbiol. Biotechnol. 2021, 105, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Menendez, V.; Martin, J.; Siles, J.A.; Gonzalez-Tejero, M.R.; Reyes, F.; Platas, G.; Tormo, J.R.; Genilloud, O. Biodiversity and Chemotaxonomy of Preussia Isolates from the Iberian Peninsula. Mycol. Prog. 2017, 16, 713–728. [Google Scholar] [CrossRef]
- Kim, Y.-G.; Lee, J.-H.; Park, J.G.; Lee, J. Inhibition of Candida albicans and Staphylococcus aureus Biofilms by Centipede Oil and Linoleic Acid. Biofouling 2020, 36, 126–137. [Google Scholar] [CrossRef]
- Thibane, V.S.; Ells, R.; Hugo, A.; Albertyn, J.; Van Rensburg, W.J.J.; Van Wyk, P.W.J.; Kock, J.L.F.; Pohl, C.H. Polyunsaturated Fatty Acids Cause Apoptosis in C. Albicans and C. Dubliniensis Biofilms. Biochim. Biophys. Acta (BBA) Gen. Subj. 2012, 1820, 1463–1468. [Google Scholar] [CrossRef]
- Estrela, A.; Abraham, W.-R. Fungal Metabolites for the Control of Biofilm Infections. Agriculture 2016, 6, 37. [Google Scholar] [CrossRef]
- Lee, S.; Suh, Y.J.; Yang, S.; Hong, D.G.; Ishigami, A.; Kim, H.; Hur, J.-S.; Chang, S.-C.; Lee, J. Neuroprotective and Anti-Inflammatory Effects of Evernic Acid in an MPTP-Induced Parkinson’s Disease Model. Int. J. Mol. Sci. 2021, 22, 2098. [Google Scholar] [CrossRef]
- Popović, V.; Stojković, D.; Nikolić, M.; Heyerick, A.; Petrović, S.; Soković, M.; Niketić, M. Extracts of Three Laserpitium L. Species and Their Principal Components Laserpitine and Sesquiterpene Lactones Inhibit Microbial Growth and Biofilm Formation by Oral Candida Isolates. Food Funct. 2015, 6, 1205–1211. [Google Scholar] [CrossRef]
- Pompilio, A.; Pomponio, S.; Di Vincenzo, V.; Crocetta, V.; Nicoletti, M.; Piovano, M.; Garbarino, J.A.; Di Bonaventura, G. Antimicrobial and Antibiofilm Activity of Secondary Metabolites of Lichens against Methicillin-Resistant Staphylococcus aureus Strains from Cystic Fibrosis Patients. Future Microbiol. 2013, 8, 281–292. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract References | Fungi | Percent Inhibition Observed at 100 µg/mL | |
|---|---|---|---|
| 24 h | 48 h | ||
| AP_PDA | Anthostomella pinea | 47.8 | 64.2 * |
| AP_SAB | Anthostomella pinea | 39.6 | 65.3 * |
| AP_MEA | Anthostomella pinea | 4.7 | 71.3 * |
| BM_1_PDA | Biscogniauxia mediterranea (G1) | 17.0 | 46.2 |
| BM_1_SAB | Biscogniauxia mediterranea (G1) | 37.0 | 30.7 |
| BM_1_MEA | Biscogniauxia mediterranea (G1) | 25.3 | 30.6 |
| BM_2_PDA | Biscogniauxia mediterranea (G2) | 0 | 25.5 |
| BM_2_SAB | Biscogniauxia mediterranea (G2) | 14.1 | 16.9 |
| BN_1_PDA | Biscogniauxia nummularia (G1) | 0 | 38.5 |
| BN_1_SAB | Biscogniauxia nummularia (G1) | 1.8 | 18.1 |
| BN_1_MEA | Biscogniauxia nummularia (G1) | 11.8 | 40.7 |
| BN_2_PDA | Biscogniauxia nummularia (G2) | 17.0 | 28.3 |
| BN_2_SAB | Biscogniauxia nummularia (G2) | 10.2 | 25.5 |
| CD_PDA | Coniochaeta decumbens | 10.5 | 39.6 |
| CD_SAB | Coniochaeta decumbens | 14.2 | 11.1 |
| CD_MEA | Coniochaeta decumbens | 38.7 | 65.8 * |
| CH_PDA | Coniochaeta hoffmanii | 64.2 * | 45.9 |
| CH_SAB | Coniochaeta hoffmanii | 0 | 18.4 |
| CH_MEA | Coniochaeta hoffmanii | 40.3 | 40.3 |
| CL_PDA | Coniochaeta lignicola | 32.2 | 64.6 * |
| CL_SAB | Coniochaeta lignicola | 26.3 | 46.9 |
| CL_MEA | Coniochaeta lignicola | 19.7 | 48.5 |
| FA_PDA | Fusarium avenaceum | 0 | 19.5 |
| FA_SAB | Fusarium avenaceum | 8.3 | 28.2 |
| FA_MEA | Fusarium avenaceum | 22.0 | 29.5 |
| PV_PDA | Peziza varia | 24.6 | 40.5 |
| PV_SAB | Peziza varia | 29.8 | 35.8 |
| PV_MEA | Peziza varia | 48.7 | 44.8 |
| PSP_MEA | Plectania sp. | 48.7 | 59.4 * |
| PP_1_PDA | Preussia persica (G1) | 37.4 | 55.0 * |
| PP_1_SAB | Preussia persica (G1) | 32.5 | 37.8 |
| PP_1_MEA | Preussia persica (G1) | 25.7 | 46.0 |
| PP_2_PDA | Preussia persica (G2) | 25.6 | 50.6 * |
| PP_2_SAB | Preussia persica (G2) | 28.1 | 54.4 * |
| PP_2_MEA | Preussia persica (G2) | 62.1 * | 64.4 * |
| SF_PDA | Sordaria fimicola | 16.5 | 33.1 |
| SF_SAB | Sordaria fimicola | 33.7 | 23.0 |
| SF_MEA | Sordaria fimicola | 35.8 | 35.4 |
| Extract References | Fungi | Percent Inhibition Observed at 100 µg/mL | |
|---|---|---|---|
| 24 h | 48 h | ||
| AP_PDA | Anthostomella pinea | 64.0 * | 50.7 * |
| AP_SAB | Anthostomella pinea | 8.8 | 59.9 * |
| AP_MEA | Anthostomella pinea | 60.0 * | 68.8 * |
| CD_MEA | Coniochaeta decumbens | 17.4 | 35.9 |
| CL_PDA | Coniochaeta lignicola | 30.4 | 46.2 |
| PSP_MEA | Plectania sp. | 0 | 25.8 |
| PP_1_PDA | Preussia persica (G1) | 0 | 0 |
| PP_2_PDA | Preussia persica (G2) | 0 | 0 |
| PP_2_SAB | Preussia persica (G2) | 0 | 0 |
| PP_2_MEA | Preussia persica (G2) | 63.3 * | 67.9 * |
| Extract References | CI1 | CI2 | CI3 |
|---|---|---|---|
| 48 h | 48 h | 48 h | |
| AP_PDA | 58.6 * | 50.3 * | 29.6 |
| AP_SAB | 52.2 * | 5.0 | 8.2 |
| AP_MEA | 59.5 * | 40.5 | 19.0 |
| PP_2_MEA | 67.9 * | 75.3 * | 56.9 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toure, S.; Millot, M.; Ory, L.; Roullier, C.; Khaldi, Z.; Pichon, V.; Girardot, M.; Imbert, C.; Mambu, L. Access to Anti-Biofilm Compounds from Endolichenic Fungi Using a Bioguided Networking Screening. J. Fungi 2022, 8, 1012. https://doi.org/10.3390/jof8101012
Toure S, Millot M, Ory L, Roullier C, Khaldi Z, Pichon V, Girardot M, Imbert C, Mambu L. Access to Anti-Biofilm Compounds from Endolichenic Fungi Using a Bioguided Networking Screening. Journal of Fungi. 2022; 8(10):1012. https://doi.org/10.3390/jof8101012
Chicago/Turabian StyleToure, Seinde, Marion Millot, Lucie Ory, Catherine Roullier, Zineb Khaldi, Valentin Pichon, Marion Girardot, Christine Imbert, and Lengo Mambu. 2022. "Access to Anti-Biofilm Compounds from Endolichenic Fungi Using a Bioguided Networking Screening" Journal of Fungi 8, no. 10: 1012. https://doi.org/10.3390/jof8101012