
A Putative D-Arabinono-1,4-lactone Oxidase, MoAlo1, Is Required for Fungal Growth, Conidiogenesis, and Pathogenicity in Magnaporthe oryzae

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Culture Conditions
2.2. Generation of the MoALO1 Null Mutant and Complementation Strain in M. oryzae
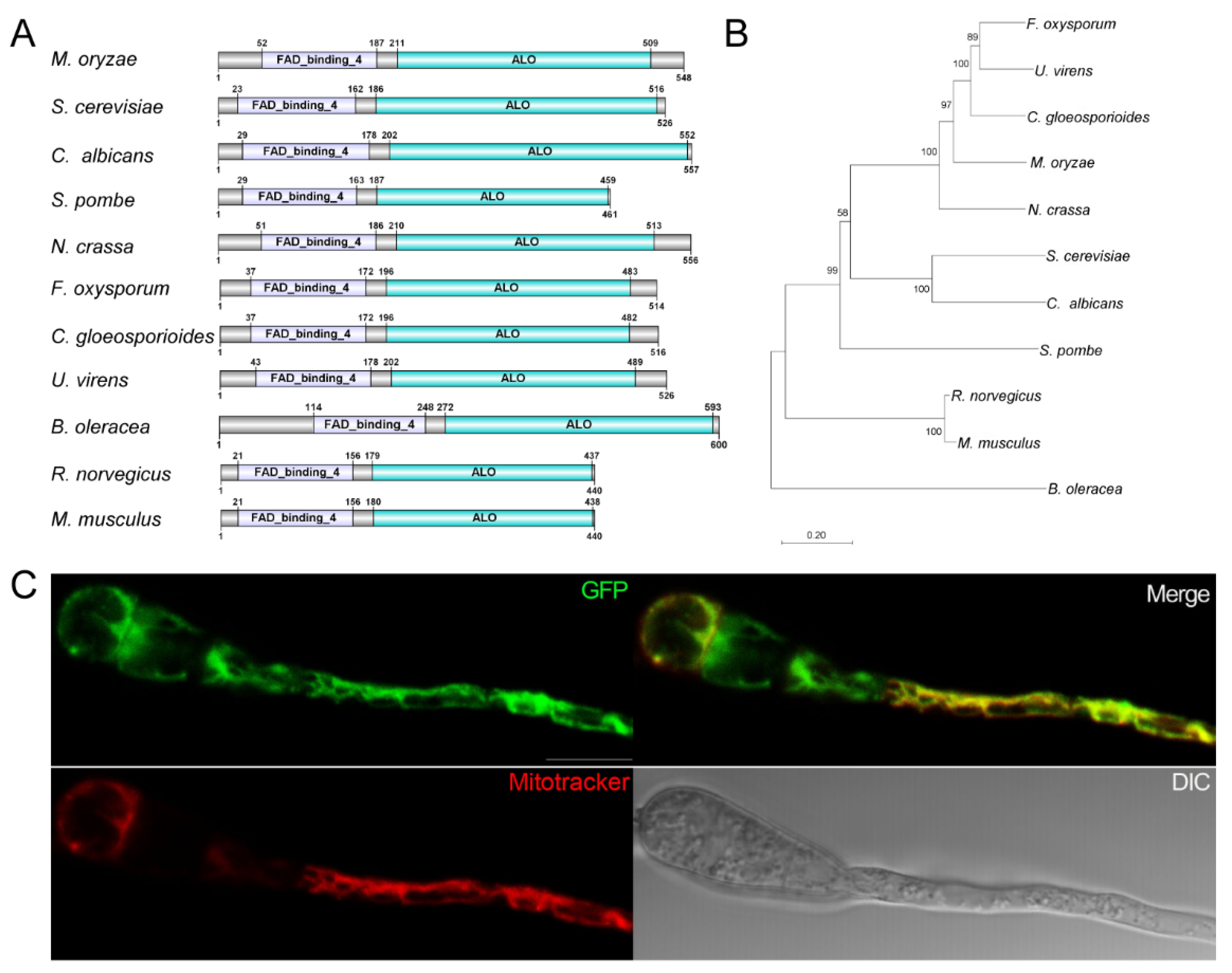
2.3. Cellular Localization of MoAlo1 in M. oryzae
2.4. The Growth, Conidiation, Glycogen Distribution, and Oxidative Stress of M. oryzae
2.5. Pathogenicity and Penetration Assays
3. Results
3.1. Identification and Subcellular Localization of MoAlo1
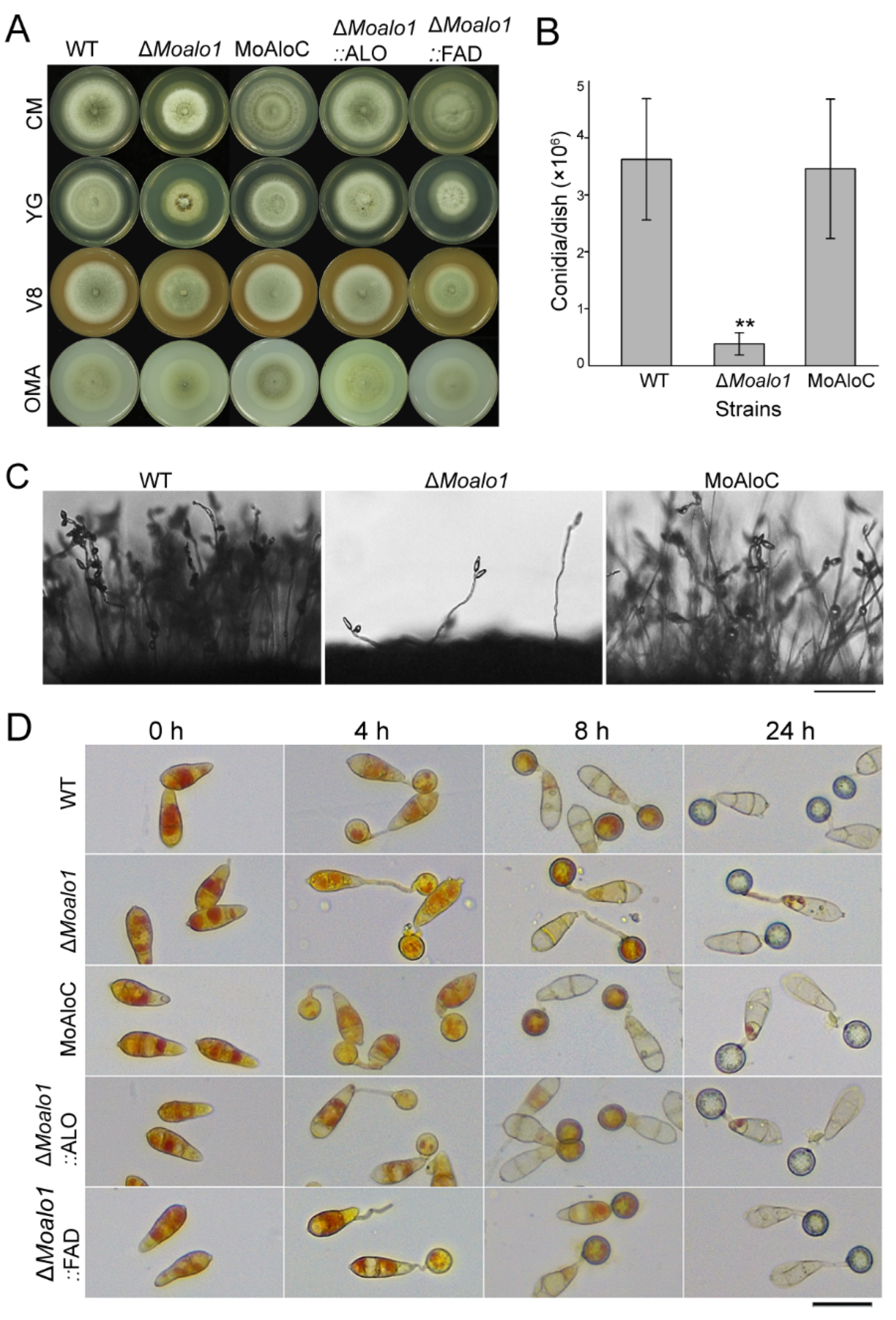
3.2. MoAlo1 Is Important for Vegetative Growth and Conidiogenesis
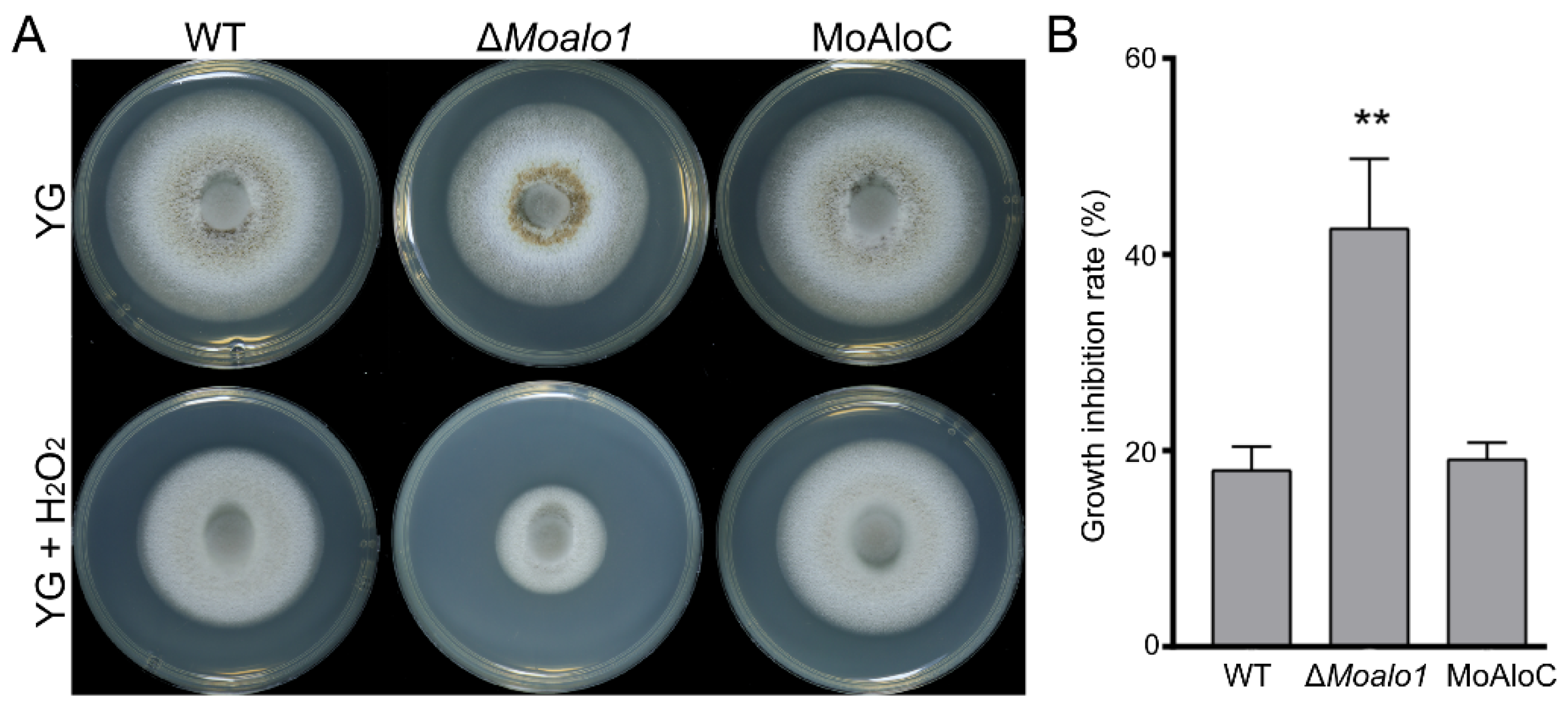
3.3. MoAlo1 Plays a Role in Oxidative Stress
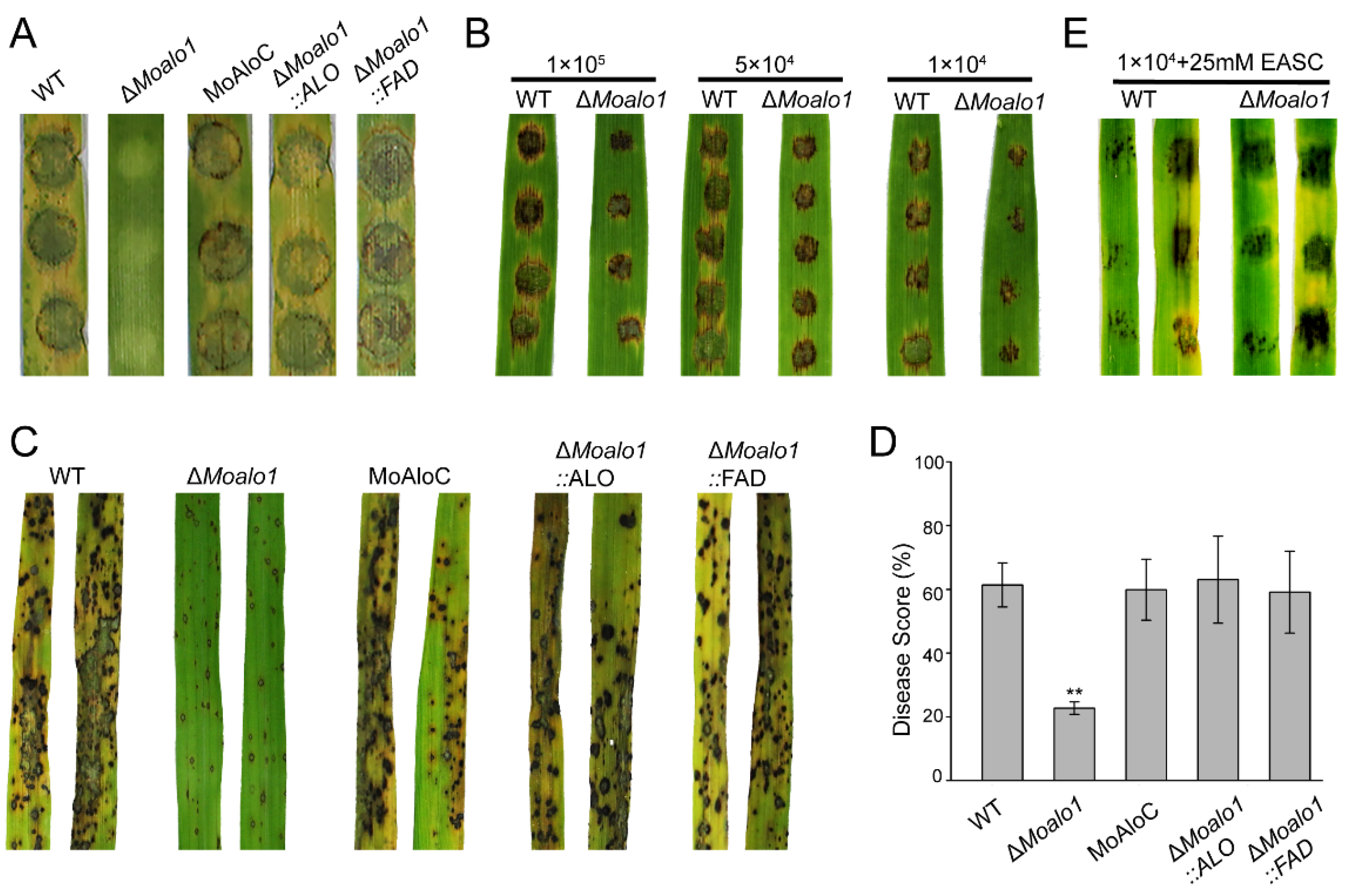
3.4. MoAlo1 Is Required for Pathogenicity
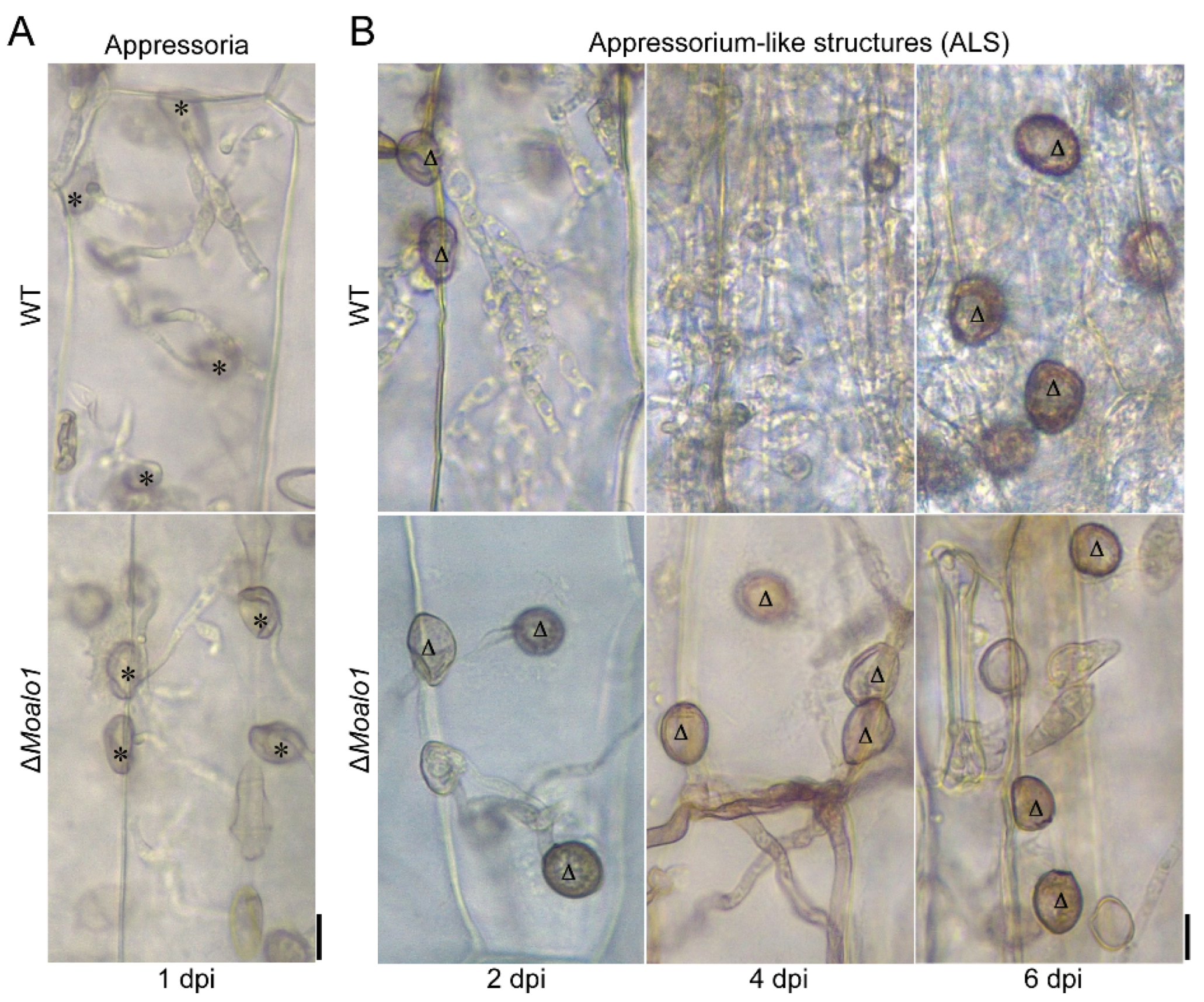
3.5. MoAlo1 Play Pleiotropic Roles in Penetration
3.6. The ALO Domain and FAD_Binding_4 Domain Can Restore the Defects in the MoALO1 Null Mutant
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′-3′) |
|---|---|
| MoALO1 up (F) | AGGCTAACTGACACTCTAGACATTTCCCGTTAGGCGTC |
| MoALO1 up (R) | TTCAATATCATCTTCTGGTAAGATCGGACGTCAAG |
| MoALO1 down (F) | CGTCACCGAGATTTAGCGGAACTTTGCAGAGAAT |
| MoALO1 down (R) | CGACGGCCAGTGCCAAGCTTGGCATTGCTGCTGATAAACT |
| BAR-F | CACCATCGTCAACCACTACATC |
| BAR-R | GCGACGAGCCAGGGATA |
| MoALO1-L-F | ATGGTCTTCCTGAGCTTG |
| MoALO1-S-F | AAATGTCGCGGACGGCATTCC |
| MoALO1-S-R | CCGAAACAAATCCGGGTCC |
| qTUBLIN-F | GTAGTTCAGGTCACCGTATGAG |
| qTUBLIN-R | CCATCCCGAGCTTGTTGATA |
| qBAR-F | CACCATCGTCAACCACTACATC |
| qBAR-R | GCGACGAGCCAGGGATA |
| MoALO1C-H3-F | CAATCACAATGGCCGGATCCATGCGACGTAACAAATCAAG |
| MoALO1C-H3-R | CCCTTGCTCACCATCCCGGGTTTTCTGCCTCCGCAGTCCC |
| FAD-F | CAATCACAATGGCCGGATCCATGGGAGCTGTACCTGCAGC |
| FAD-R | CCCTTGCTCACCATCCCGGGACTGCAGGTCCGCGTCGTCC |
| ALO-F | CAATCACAATGGCCGGATCCATGGAGCTTCCGGGCCGTGC |
| ALO-R | CCCTTGCTCACCATCCCGGGTCGACGATGCCATGGCCCAG |
References
- Ebbole, D.J. Magnaporthe as a model for understanding host-pathogen interactions. Annu. Rev. Phytopathol. 2007, 45, 437–456. [Google Scholar] [CrossRef]
- Howard, T.M.B.; Richard, J. In vitro development of penetration structures in the rice blast fungus Magnaporthe grisea. Can. J. Bot. 1990, 68, 329–342. [Google Scholar]
- Tucker, S.L.; Talbot, N.J. Surface attachment and pre-penetration stage development by plant pathogenic fungi. Annu. Rev. Phytopathol. 2001, 39, 385–417. [Google Scholar] [CrossRef]
- Skamnioti, P.; Gurr, S.J. Magnaporthe grisea cutinase2 mediates appressorium differentiation and host penetration and is required for full virulence. Plant Cell 2007, 19, 2674–2689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talbot, N.J. On the trail of a cereal killer: Exploring the biology of Magnaporthe grisea. Annu. Rev. Microbiol. 2003, 57, 177–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CONKLIN, P.L.; BARTH, C. Ascorbic acid, a familiar small molecule intertwined in the response of plants to ozone, pathogens, and the onset of senescence. Plant Cell Environ. 2004, 27, 959–970. [Google Scholar] [CrossRef]
- Wheeler, G.L.; Jones, M.A.; Smirnoff, N. The biosynthetic pathway of vitamin C in higher plants. Nature 1998, 393, 365–369. [Google Scholar] [CrossRef]
- Burns, J.J. Missing step in man, monkey and guinea pig required for the biosynthesis of L-ascorbic acid. Nature 1957, 180, 553. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, C.R.; Chatterjee, I.B. L-ascorbic acid synthesis in birds: Phylogenetic trend. Science 1969, 164, 435–436. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, I.B. Evolution and the biosynthesis of ascorbic acid. Science 1973, 182, 1271–1272. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.Y.; Seib, P.A.; Kramer, K.J.; Van Galen, D.A. Synthesis and properties of D-erythroascorbic acid and its vitamin C activity in the tobacco hornworm (Manduca sexta). J. Agric. Food Chem. 1993, 41, 1391–1396. [Google Scholar] [CrossRef]
- Brush, J.S.; May, H.E. A kinetic study of the mechanism of action of L-gulonolactone oxidase. J. Biol. Chem. 1966, 241, 2907–2912. [Google Scholar] [CrossRef]
- Puskás, F.; Braun, L.; Csala, M.; Kardon, T.; Marcolongo, P.; Benedetti, A.; Mandl, J.; Bánhegyi, G. Gulonolactone oxidase activity-dependent intravesicular glutathione oxidation in rat liver microsomes. FEBS Lett. 1998, 430, 293–296. [Google Scholar] [CrossRef] [Green Version]
- Lorence, A.; Chevone, B.I.; Mendes, P.; Nessler, C.L. myo-inositol oxygenase offers a possible entry point into plant ascorbate biosynthesis. Plant Physiol. 2004, 134, 1200–1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huh, W.K.; Lee, B.H.; Kim, S.T.; Kim, Y.R.; Rhie, G.E.; Baek, Y.W.; Hwang, C.S.; Lee, J.S.; Kang, S.O. D-Erythroascorbic acid is an important antioxidant molecule in Saccharomyces cerevisiae. Mol. Microbiol. 1998, 30, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Huh, W.K.; Kim, S.T.; Kim, H.; Jeong, G.; Kang, S.O. Deficiency of d-Erythroascorbic Acid Attenuates Hyphal Growth and Virulence of Candida albicans. Infect. Immun. 2001, 69, 3939–3946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwak, M.-K.; Song, S.-H.; Ku, M.; Kang, S.-O. Candida albicans erythroascorbate peroxidase regulates intracellular methylglyoxal and reactive oxygen species independently of d-erythroascorbic acid. FEBS Lett. 2015, 589, 1863–1871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huh, W.K.; Song, Y.B.; Lee, Y.S.; Ha, C.W.; Kim, S.T.; Kang, S.O. D-Erythroascorbic acid activates cyanide-resistant respiration in Candida albicans. Biochem. Biophys. Res. Commun. 2008, 369, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Baroja-Mazo, A.; del Valle, P.; Rúa, J.; de Cima, S.; Busto, F.; de Arriaga, D.; Smirnoff, N. Characterisation and biosynthesis of D-erythroascorbic acid in Phycomyces blakesleeanus. Fungal Genet. Biol. 2005, 42, 390–402. [Google Scholar] [CrossRef]
- Talbot, N.J.; Kershaw, M.J.; Wakley, G.E.; De Vries, O.; Wessels, J.; Hamer, J.E. MPG1 Encodes a Fungal Hydrophobin Involved in Surface Interactions during Infection-Related Development of Magnaporthe grisea. Plant Cell 1996, 8, 985–999. [Google Scholar] [CrossRef]
- Liu, X.H.; Chen, S.M.; Gao, H.; Ning, G.A.; Shi, H.B.; Wang, Y.; Dong, B.; Qi, Y.Y.; Zhang, D.M.; Lu, G.-d.; et al. The small GTPase MoYpt7 is required for membrane fusion in autophagy and pathogenicity of Magnaporthe oryzae. Environ. Microbiol. 2015, 17, 4495–4510. [Google Scholar] [CrossRef]
- Lu, J.; Cao, H.; Zhang, L.; Huang, P.; Lin, F. Systematic analysis of Zn2Cys6 transcription factors required for development and pathogenicity by high-throughput gene knockout in the rice blast fungus. PLoS Pathog. 2014, 10, e1004432. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Shi, H.; Liang, S.; Ning, G.; Xu, N.; Lu, J.; Liu, X.; Lin, F. MoARG1, MoARG5,6 and MoARG7 involved in arginine biosynthesis are essential for growth, conidiogenesis, sexual reproduction, and pathogenicity in Magnaporthe oryzae. Microbiol. Res. 2015, 180, 11–22. [Google Scholar] [CrossRef]
- Liu, X.-H.; Ning, G.-A.; Huang, L.-Y.; Zhao, Y.-H.; Dong, B.; Lu, J.-P.; Lin, F.-C. Calpains are involved in asexual and sexual development, cell wall integrity and pathogenicity of the rice blast fungus. Sci. Rep. 2016, 6, 31204. [Google Scholar] [CrossRef]
- Ostergaard, J.; Persiau, G.; Davey, M.W.; Bauw, G.; Van Montagu, M. Isolation of a cDNA coding for L-galactono-gamma-lactone dehydrogenase, an enzyme involved in the biosynthesis of ascorbic acid in plants. Purification, characterization, cDNA cloning, and expression in yeast. J. Biol. Chem. 1997, 272, 30009–30016. [Google Scholar] [CrossRef] [Green Version]
- Koshizaka, T.; Nishikimi, M.; Ozawa, T.; Yagi, K. Isolation and sequence analysis of a complementary DNA encoding rat liver L-gulono-gamma-lactone oxidase, a key enzyme for L-ascorbic acid biosynthesis. J. Biol. Chem. 1988, 263, 1619–1621. [Google Scholar] [CrossRef]
- Levine, M.; Rumsey, S.C.; Daruwala, R.; Park, J.B.; Wang, Y. Criteria and recommendations for vitamin C intake. JAMA 1999, 281, 1415–1423. [Google Scholar] [CrossRef]
- Hancock, R.D.; Galpin, J.R.; Viola, R. Biosynthesis of L-ascorbic acid (vitamin C) by Saccharomyces cerevisiae. FEMS Microbiol. Lett. 2000, 186, 245–250. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, S.R.; Prathalingam, S.R.; Taylor, M.C.; Horn, D.; Kelly, J.M. Vitamin C biosynthesis in trypanosomes: A role for the glycosome. Proc. Natl. Acad. Sci. USA 2005, 102, 11645–11650. [Google Scholar] [CrossRef] [Green Version]
- Keates, S.E.; Loewus, F.A.; Helms, G.L.; Zink, D.L. 5-O-(alpha-D-galactopyranosyl)-D-glycero-pent-2-enono-1,4-lactone: Characterization in the oxalate-producing fungus, Sclerotinia sclerotiorum. Phytochemistry 1998, 49, 2397. [Google Scholar] [CrossRef]
- Berthold, D.A.; Siedow, J.N. Partial purification of the cyanide-resistant alternative oxidase of skunk cabbage (Symplocarpus foetidus) mitochondria. Plant Physiol. 1993, 101, 113–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Zhao, H.; Luo, C.; Du, L.; Cheng, J.; Xie, J.; Jiang, D.; Fu, Y. CmAim24 Is Essential for Mitochondrial Morphology, Conidiogenesis, and Mycoparasitism in Coniothyrium minitans. Appl. Environ. Microbiol. 2020, 86, e02291-19. [Google Scholar] [CrossRef] [PubMed]
- Bartoli, C.G.; Pastori, G.M.; Foyer, C.H. Ascorbate biosynthesis in mitochondria is linked to the electron transport chain between complexes III and IV. Plant Physiol. 2000, 123, 335–343. [Google Scholar] [CrossRef] [Green Version]
- Manhas, R.; Anand, S.; Tripathi, P.; Madhubala, R. Deletion of Vitamin C biosynthesis enzyme, Arabino-1, 4-lactone oxidase in Leishmania donovani results in increased pro-inflammatory responses from host immune cells. Mol. Microbiol. 2014, 91, 1227–1239. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.J.; Ferrari, M.A.; Roach, D.H.; Money, N.P. Penetration of hard substrates by a fungus employing enormous turgor pressures. Proc. Natl. Acad. Sci. USA 1992, 88, 11281–11284. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Zheng, W.; Zheng, S.; Zhang, D.; Sang, W.; Chen, X.; Li, G.; Lu, G.; Wang, Z.; Andrianopoulos, A. Rac1 Is Required for Pathogenicity and Chm1-Dependent Conidiogenesis in Rice Fungal Pathogen Magnaporthe grisea. PLoS Pathog. 2008, 4, e1000202. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.Z.; Ramos-Pamplona, M.; Naqvi, N.I. Autophagy-assisted glycogen catabolism regulates asexual differentiation in Magnaporthe oryzae. Autophagy 2009, 5, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, C.; Saklani, B.K.; Singh, P.K.; Asthana, R.K.; Sharma, T.R. Variation in the LRR region of Pi54 protein alters its interaction with the AvrPi54 protein revealed by in silico analysis. PLoS ONE 2019, 14, e0224088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.Y.; Yu, Q.; Dong, B.; Zhang, Y.; Liu, X.H.; Lin, F.C.; Liang, S. MoLEU1, MoLEU2, and MoLEU4 regulated by MoLEU3 are involved in leucine biosynthesis, fungal development, and pathogenicity in Magnaporthe oryzae. Environ. Microbiol. Rep. 2019, 11, 784–796. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.M.; Liang, S.; Shi, H.B.; Lu, J.P.; Dong, B.; Liao, Q.S.; Lin, F.C.; Liu, X.H. VPS9 domain-containing proteins are essential for autophagy and endocytosis in Pyricularia oryzae. Environ. Microbiol. 2018, 20, 1516–1530. [Google Scholar] [CrossRef]
- Liu, W.; Xie, S.; Zhao, X.; Chen, X.; Zheng, W.; Lu, G.; Xu, J.R.; Wang, Z. A homeobox gene is essential for conidiogenesis of the rice blast fungus Magnaporthe oryzae. Mol. Plant Microbe Interact. 2010, 23, 366–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arrigoni, O.; De Tullio, M.C. Ascorbic acid: Much more than just an antioxidant. Biochim. Biophys. Acta (BBA) Gen. Subj. 2002, 1569, 1–9. [Google Scholar] [CrossRef]
- Loewus, F.A.; Saito, K.; Suto, R.K.; Maring, E. Conversion of D-arabinose to D-erythroascorbic acid and oxalic acid in Sclerotinia sclerotiorum. Biochem. Biophys. Res. Commun. 1995, 212, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Qin, C.; Lin, L.; Zeng, X.; Zhao, Y.; He, S.; Lu, S.; Guo, Z. Overexpression of Yeast Arabinono-1,4-Lactone Oxidase Gene (ALO) Increases Tolerance to Oxidative Stress and Al Toxicity in Transgenic Tobacco Plants. Plant Mol. Biol. Rep. 2015, 33, 806–818. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, M.-H.; Huang, L.-Y.; Sun, L.-X.; Qian, H.; Wei, Y.-Y.; Liang, S.; Zhu, X.-M.; Li, L.; Lu, J.-P.; Lin, F.-C.; et al. A Putative D-Arabinono-1,4-lactone Oxidase, MoAlo1, Is Required for Fungal Growth, Conidiogenesis, and Pathogenicity in Magnaporthe oryzae. J. Fungi 2022, 8, 72. https://doi.org/10.3390/jof8010072
Wu M-H, Huang L-Y, Sun L-X, Qian H, Wei Y-Y, Liang S, Zhu X-M, Li L, Lu J-P, Lin F-C, et al. A Putative D-Arabinono-1,4-lactone Oxidase, MoAlo1, Is Required for Fungal Growth, Conidiogenesis, and Pathogenicity in Magnaporthe oryzae. Journal of Fungi. 2022; 8(1):72. https://doi.org/10.3390/jof8010072
Chicago/Turabian StyleWu, Ming-Hua, Lu-Yao Huang, Li-Xiao Sun, Hui Qian, Yun-Yun Wei, Shuang Liang, Xue-Ming Zhu, Lin Li, Jian-Ping Lu, Fu-Cheng Lin, and et al. 2022. "A Putative D-Arabinono-1,4-lactone Oxidase, MoAlo1, Is Required for Fungal Growth, Conidiogenesis, and Pathogenicity in Magnaporthe oryzae" Journal of Fungi 8, no. 1: 72. https://doi.org/10.3390/jof8010072