
Local Environmental Conditions Promote High Turnover Diversity of Benthic Deep-Sea Fungi in the Ross Sea (Antarctica)

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
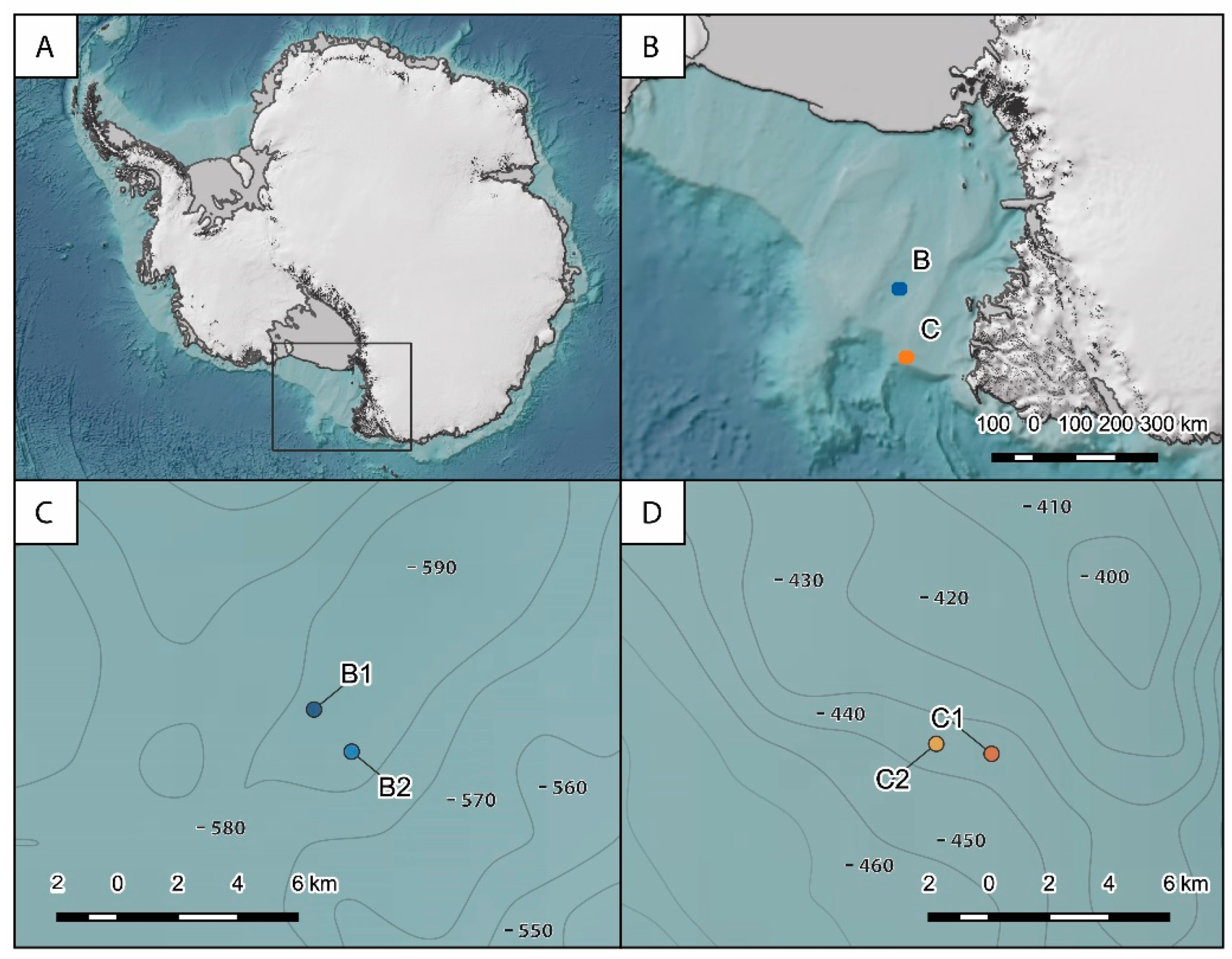
2.1. Study Area and Sampling Strategy
2.2. Trophic Conditions
2.3. DNA Extraction and Purification for Molecular Analysis
2.4. Fungal Abundance Estimated by Quantitative Real-Time PCR (qPCR)
2.5. Fungal Diversity and Assemblage Composition
2.6. Data Analyses
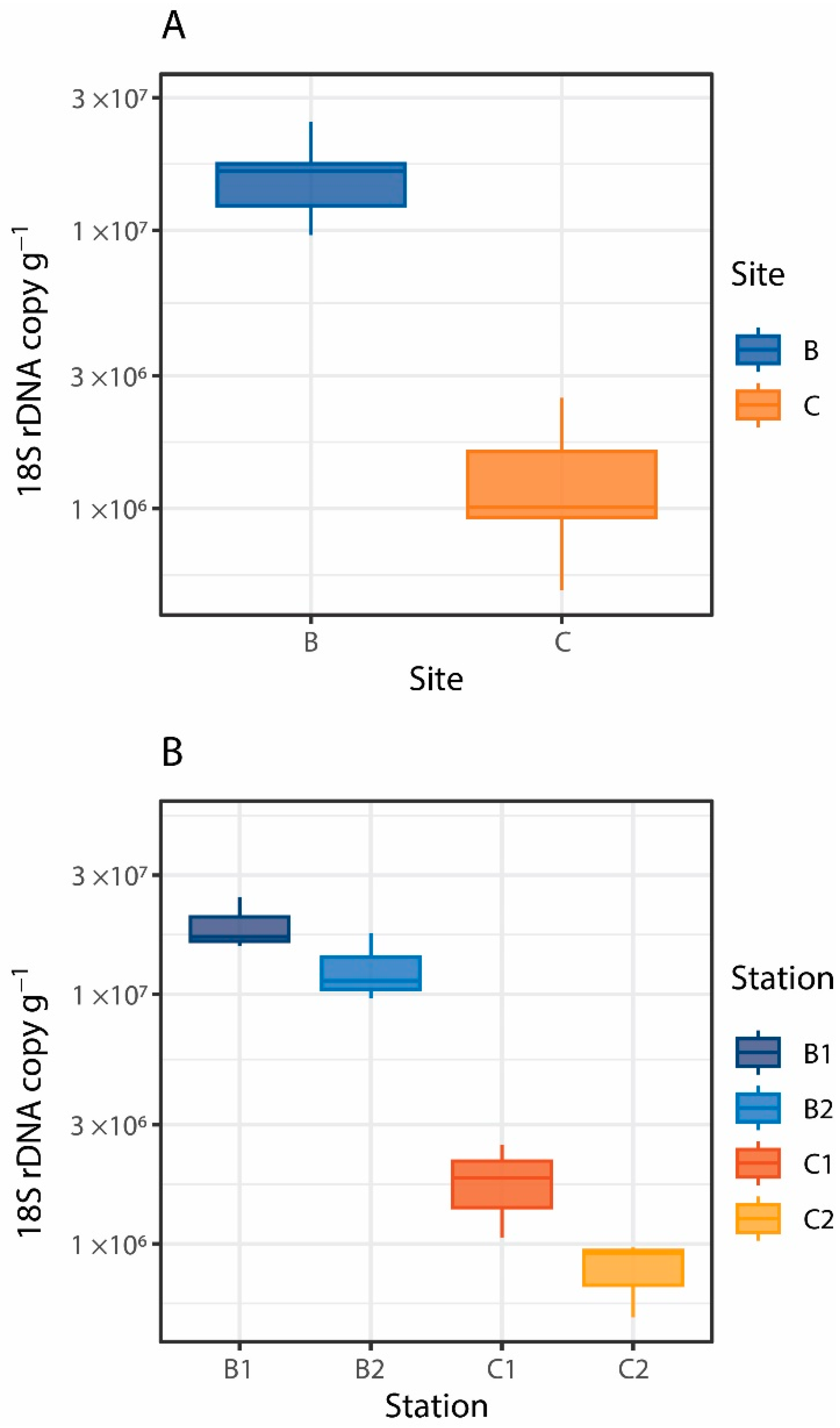
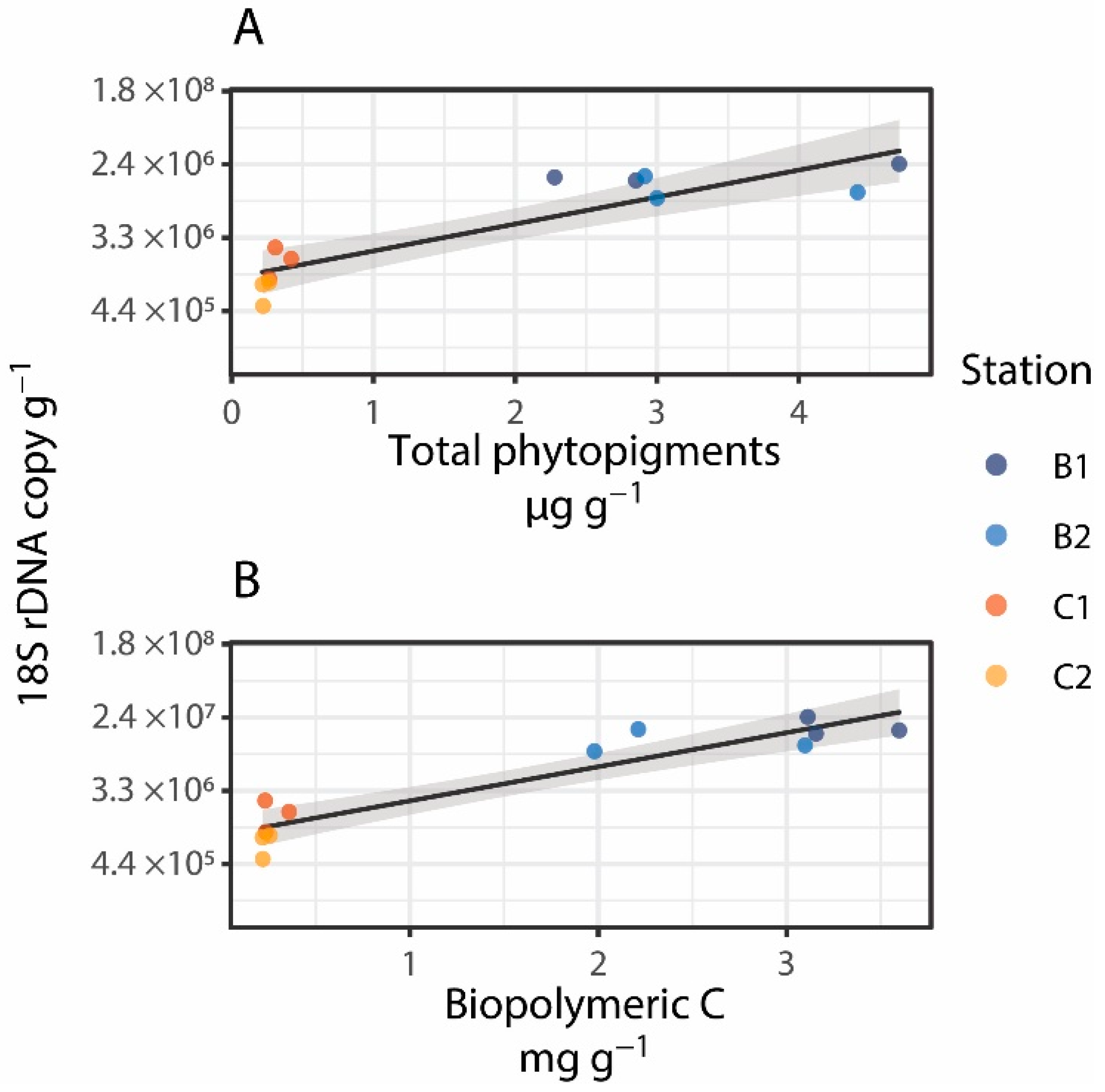
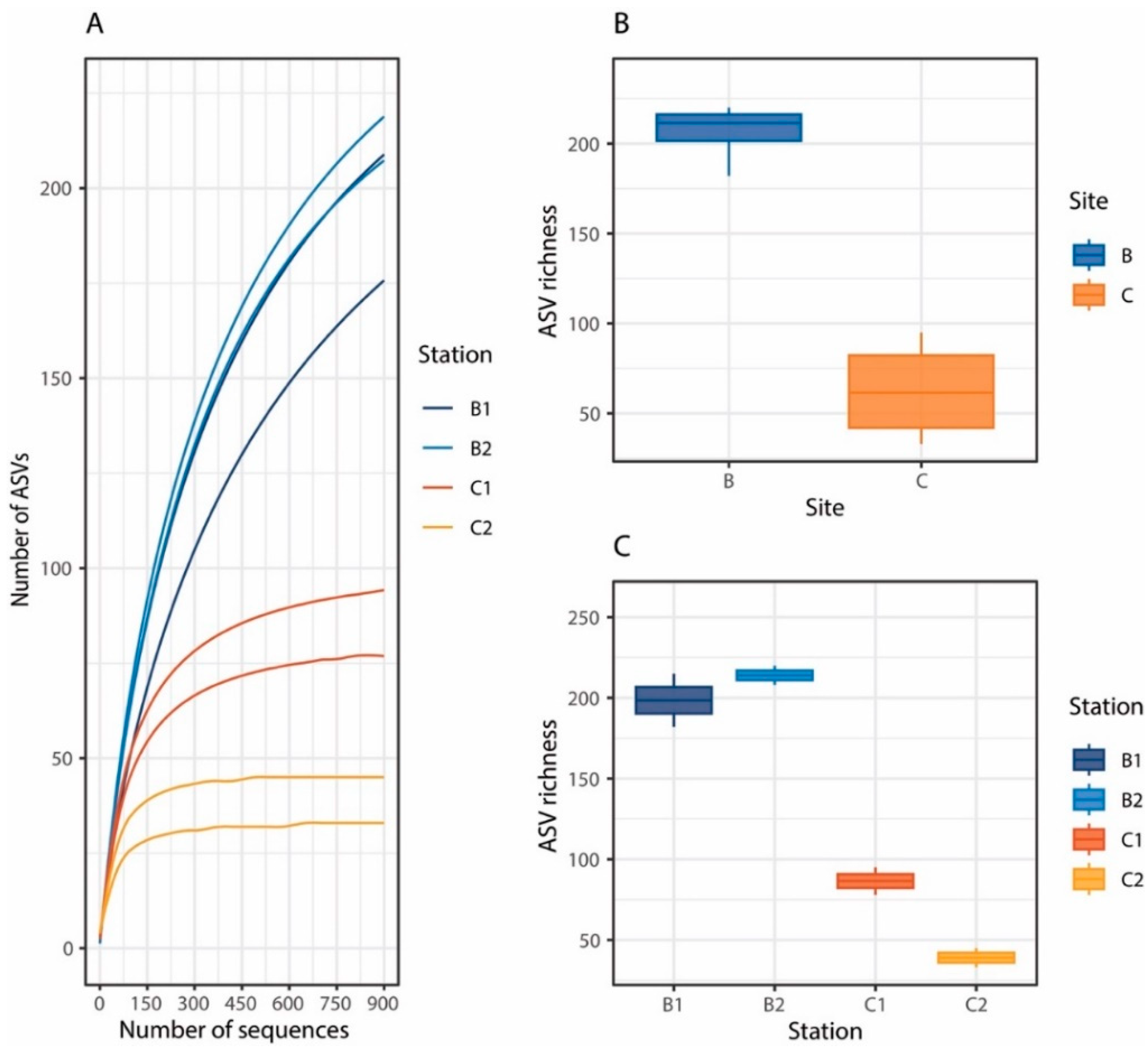
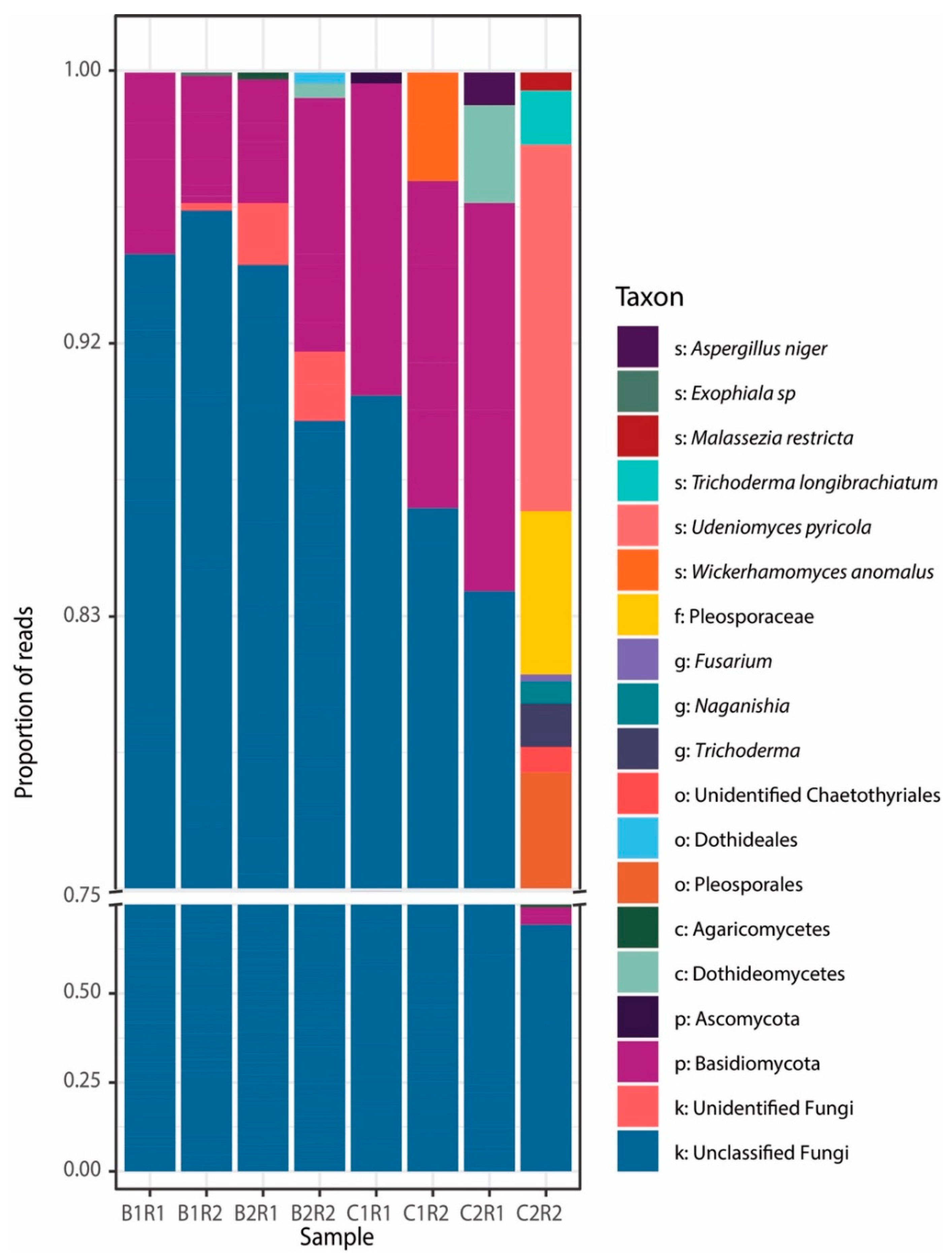
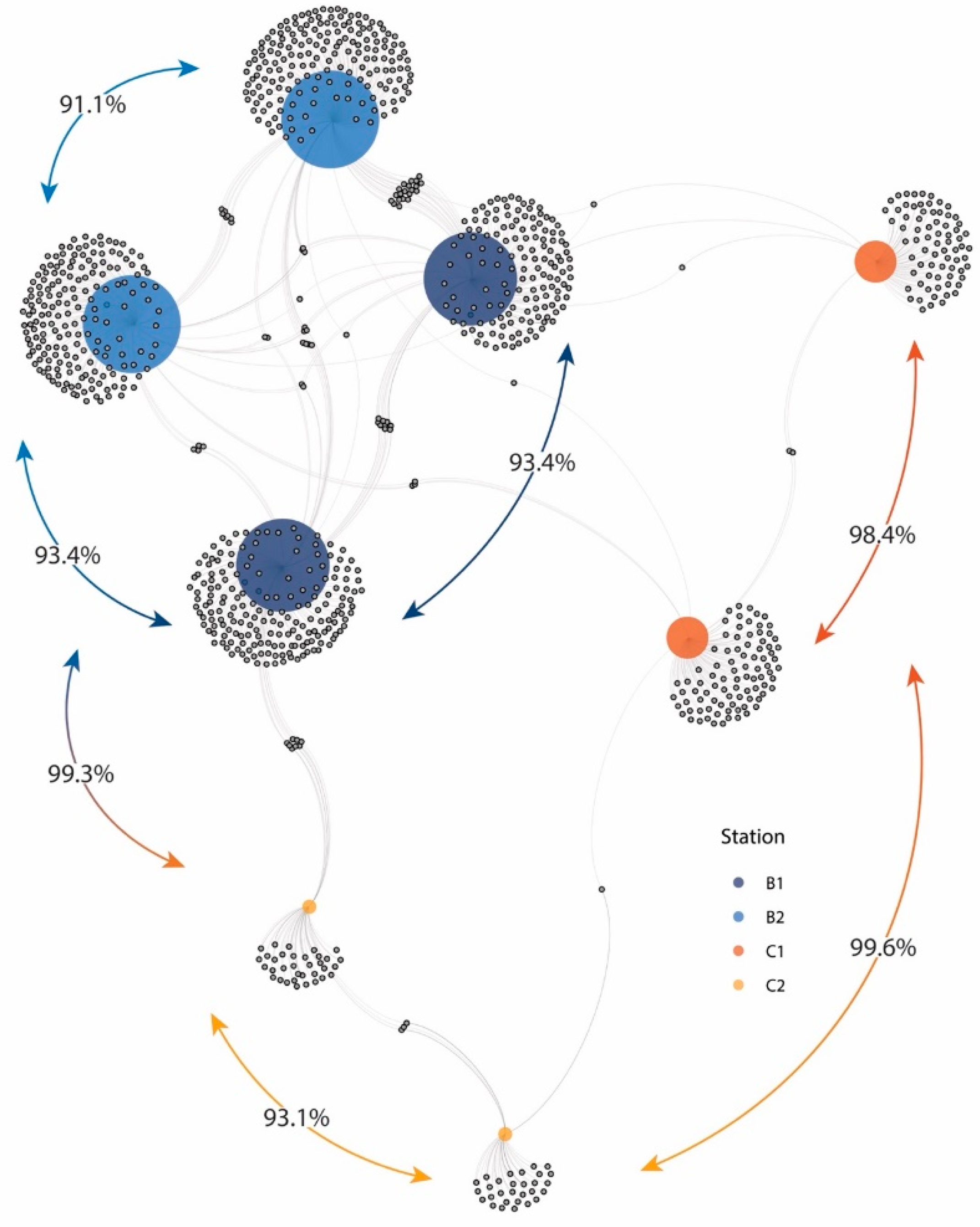
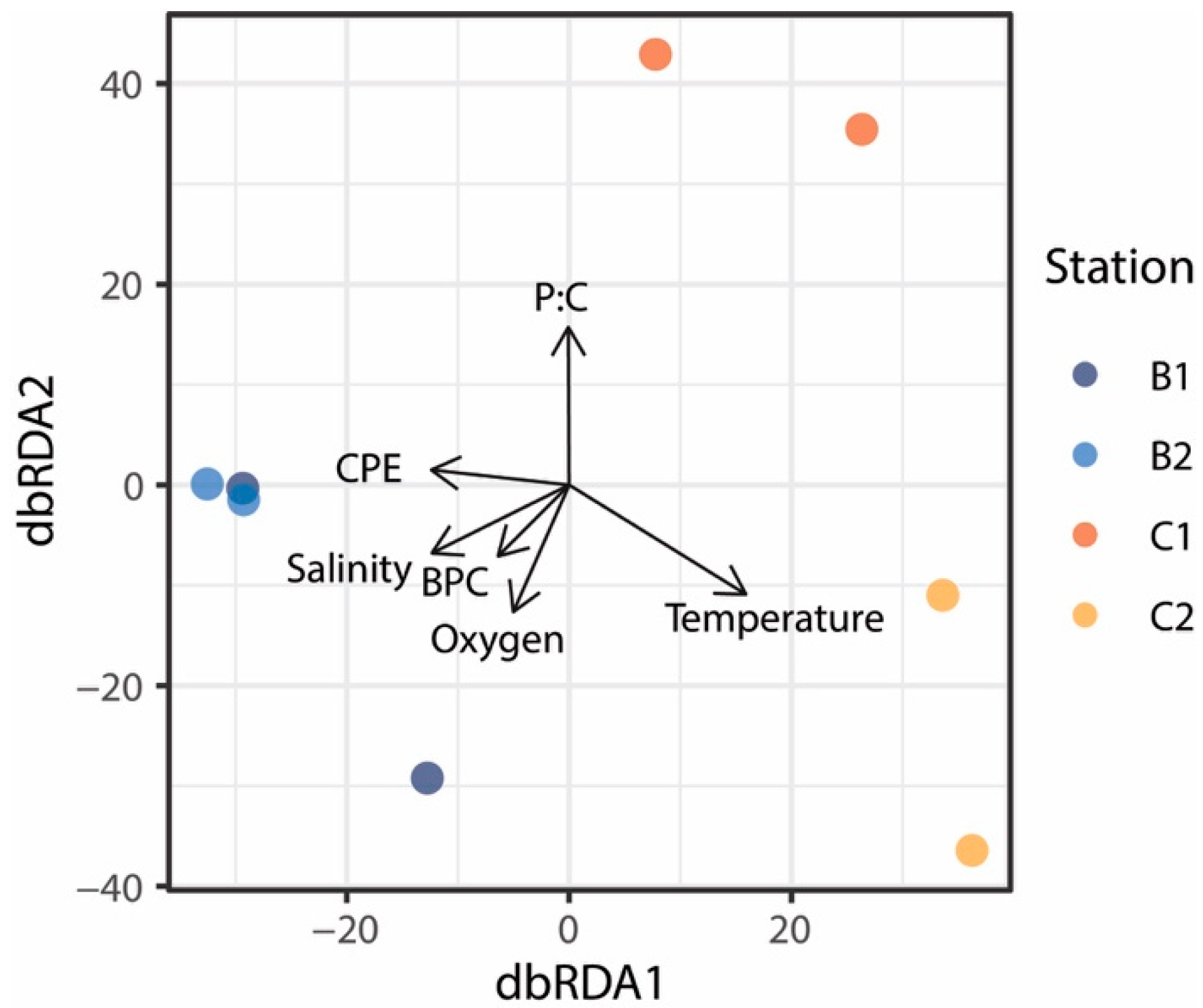
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arrigo, K.R.; van Dijken, G.L.; Bushinsky, S. Primary production in the Southern Ocean, 1997–2006. J. Geophys. Res. Ocean 2008, 113, 1–27. [Google Scholar] [CrossRef]
- Arrigo, K.R.; van Dijken, G.; Long, M. Coastal Southern Ocean: A strong anthropogenic CO2 sink. Geophys. Res. Lett. 2008, 35, 1–6. [Google Scholar]
- Arrigo, K.R.; van Dijken, G.L. Phytoplankton dynamics within 37 Antarctic coastal polynya systems. J. Geophys. Res. Ocean 2003, 108, 3271. [Google Scholar] [CrossRef]
- Arrigo, K.R.; Van Dijken, G.L. Interannual variation in air-sea CO2 flux in the Ross Sea, Antarctica: A model analysis. J. Geophys. Res. Ocean 2007, 112, 1–16. [Google Scholar]
- Smith, W.O.; Nelson, D.M. Importance of ice edge phytoplankton production in the Southern Ocean. Bioscience 1986, 36, 251–257. [Google Scholar] [CrossRef]
- Smith, W.O.; Gordon, L.I. Hyperproductivity of the Ross Sea (Antarctica) polynya during austral spring. Geophys. Res. Lett. 1997, 24, 233–236. [Google Scholar] [CrossRef]
- Sweeney, C. The annual cycle of surface water CO2 and O2 in the Ross Sea: A model for gas exchange on the continental shelves of Antarctica. Biogeochem. Ross Sea 2003, 78, 295–312. [Google Scholar]
- Arrigo, K.R.; Van Dijken, G.L. Annual changes in sea-ice, chlorophyll α, and primary production in the Ross Sea, Antarctica. Deep. Res. Part II Top. Stud. Oceanogr. 2004, 51, 117–138. [Google Scholar] [CrossRef]
- Brandt, A.; De Broyer, C.; De Mesel, I.; Ellingsen, K.E.; Gooday, A.J.; Hilbig, B.; Linse, K.; Thomson, M.R.A.; Tyler, P.A. The biodiversity of the deep Southern Ocean benthos. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 39–66. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, H.J. Antarctic Marine Biodiversity—What Do We Know About the Distribution of Life in the Southern Ocean? PLoS ONE 2010, 5, e11683. [Google Scholar] [CrossRef] [Green Version]
- Danovaro, R.; Snelgrove, P.V.R.; Tyler, P. Challenging the paradigms of deep-sea ecology. Trends Ecol. Evol. 2014, 29, 465–475. [Google Scholar] [CrossRef]
- Bernhard, J.M.; Kormas, K.; Pachiadaki, M.G.; Rocke, E.; Beaudoin, D.J.; Morrison, C.; Visscher, P.T.; Cobban, A.; Starczak, V.R.; Edgcomb, V.P. Benthic protists and fungi of Mediterranean deep hypsersaline anoxic basin redoxcline sediments. Front. Microbiol. 2014, 5, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgcomb, V.P.; Pachiadaki, M.G.; Mara, P.; Kormas, K.A.; Leadbetter, E.R.; Bernhard, J.M. Gene expression profiling of microbial activities and interactions in sediments under haloclines of E. Mediterranean deep hypersaline anoxic basins. ISME J. 2016, 10, 2643–2657. [Google Scholar] [CrossRef] [Green Version]
- Barone, G.; Varrella, S.; Tangherlini, M.; Rastelli, E.; Dell’Anno, A.; Danovaro, R.; Corinaldesi, C. Marine fungi: Biotechnological perspectives from deep-hypersaline anoxic basins. Diversity 2019, 11, 113. [Google Scholar] [CrossRef] [Green Version]
- Nagahama, T.; Takahashi, E.; Nagano, Y.; Abdel-Wahab, M.A.; Miyazaki, M. Molecular evidence that deep-branching fungi are major fungal components in deep-sea methane cold-seep sediments. Environ. Microbiol. 2011, 13, 2359–2370. [Google Scholar] [CrossRef]
- Thaler, A.D.; Van Dover, C.L.; Vilgalys, R. Ascomycete phylotypes recovered from a Gulf of Mexico methane seep are identical to an uncultured deep-sea fungal clade from the Pacific. Fungal Ecol. 2012, 5, 270–273. [Google Scholar] [CrossRef]
- Burgaud, G.; Arzur, D.; Durand, L.; Cambon-Bonavita, M.A.; Barbier, G. Marine culturable yeasts in deep-sea hydrothermal vents: Species richness and association with fauna. FEMS Microbiol. Ecol. 2010, 73, 121–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgaud, G.; Le Calvez, T.; Arzur, D.; Vandenkoornhuyse, P.; Barbier, G. Diversity of culturable marine filamentous fungi from deep-sea hydrothermal vents. Environ. Microbiol. 2009, 11, 1588–1600. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Guo, S.; Pang, K.L.; Luo, Z.H. Fungi associated with chimney and sulfide samples from a South Mid-Atlantic Ridge hydrothermal site: Distribution, diversity and abundance. Deep. Res. Part I Oceanogr. Res. Pap. 2017, 123, 48–55. [Google Scholar] [CrossRef]
- Edgcomb, V.P.; Beaudoin, D.; Gast, R.; Biddle, J.F.; Teske, A. Marine subsurface eukaryotes: The fungal majority. Environ. Microbiol. 2011, 13, 172–183. [Google Scholar] [CrossRef]
- Orsi, W.; Biddle, J.F.; Edgcomb, V. Deep Sequencing of Subseafloor Eukaryotic rRNA Reveals Active Fungi across Marine Subsurface Provinces. PLoS ONE 2013, 8, e56335. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, N.F.; Zhang, Y.Q.; Liu, H.Y.; Yu, L.Y. Diversity and distribution of fungal communities in the marine sediments of Kongsfjorden, Svalbard (High Arctic). Sci. Rep. 2015, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Pachiadaki, M.G.; Rédou, V.; Beaudoin, D.J.; Burgaud, G.; Edgcomb, V.P. Fungal and prokaryotic activities in the Marine subsurface biosphere at Peru Margin and canterbury basin inferred from RNA-based analyses and microscopy. Front. Microbiol. 2016, 7, 846. [Google Scholar] [CrossRef] [PubMed]
- Barone, G.; Rastelli, E.; Corinaldesi, C.; Tangherlini, M.; Danovaro, R.; Dell’Anno, A. Benthic deep-sea fungi in submarine canyons of the Mediterranean Sea. Prog. Oceanogr. 2018, 168, 57–64. [Google Scholar] [CrossRef]
- Ogaki, M.B.; Coelho, L.C.; Vieira, R.; Neto, A.A.; Zani, C.L.; Alves, T.M.A.; Junior, P.A.S.; Murta, S.M.F.; Barbosa, E.C.; Oliveira, J.G.; et al. Cultivable fungi present in deep-sea sediments of Antarctica: Taxonomy, diversity, and bioprospecting of bioactive compounds. Extremophiles 2020, 24, 227–238. [Google Scholar] [CrossRef]
- Ogaki, M.B.; Pinto, O.H.B.; Vieira, R.; Neto, A.A.; Convey, P.; Carvalho-Silva, M.; Rosa, C.A.; Câmara, P.E.A.S.; Rosa, L.H. Fungi Present in Antarctic Deep-Sea Sediments assessed Using DNA Metabarcoding. Microb. Ecol. 2021, 82, 157–164. [Google Scholar] [CrossRef]
- Rosa, L.H. Fungi of Antarctica Diversity, Ecology and Biotechnological Applications; Springer International Publishing: Cham, Germany, 2019; ISBN 978-3-030-18367-7. [Google Scholar]
- Varrella, S.; Barone, G.; Tangherlini, M.; Rastelli, E.; Dell’anno, A.; Corinaldesi, C. Diversity, ecological role and biotechnological potential of antarctic marine fungi. J. Fungi 2021, 7, 391. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, M. The fungi: 1, 2, 3… 5.1 million species? Am. J. Bot. 2011, 98, 426–438. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Rossman, A.Y. Where are all the undescribed fungi? Phytopathology 1997, 87, 888–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffries, T.C.; Curlevski, N.J.; Brown, M.V.; Harrison, D.P.; Doblin, M.A.; Petrou, K.; Ralph, P.J.; Seymour, J.R. Partitioning of fungal assemblages across different marine habitats. Environ. Microbiol. Rep. 2016, 8, 235–238. [Google Scholar] [CrossRef]
- Vargas-Gastélum, L.; Riquelme, M. The mycobiota of the deep sea: What omics can offer. Life 2020, 10, 292. [Google Scholar] [CrossRef]
- Tisthammer, K.H.; Cobian, G.M.; Amend, A.S. Global biogeography of marine fungi is shaped by the environment. Fungal Ecol. 2016, 19, 39–46. [Google Scholar] [CrossRef]
- Li, W.; Wang, M.M.; Wang, X.G.; Cheng, X.L.; Guo, J.J.; Bian, X.M.; Cai, L. Fungal communities in sediments of subtropical Chinese seas as estimated by DNA metabarcoding. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef]
- Orsi, W.D.; Vuillemin, A.; Coskun, Ö.K.; Rodriguez, P.; Oertel, Y.; Niggemann, J.; Mohrholz, V.; Gomez-Saez, G.V. Carbon assimilating fungi from surface ocean to subseafloor revealed by coupled phylogenetic and stable isotope analysis. ISME J. 2021, 15, 1–17. [Google Scholar] [CrossRef]
- Hyde, K.D.; Jones, E.B.G.; Leaño, E.; Pointing, S.B.; Poonyth, A.D.; Vrijmoed, L.L.P. Role of fungi in marine ecosystems. Biodivers. Conserv. 1998, 7, 1147–1161. [Google Scholar] [CrossRef]
- Cathrine, S.J.; Raghukumar, C. Anaerobic denitrification in fungi from the coastal marine sediments off Goa, India. Mycol. Res. 2009, 113, 100–109. [Google Scholar] [CrossRef]
- Jebaraj, C.S.; Raghukumar, C.; Behnke, A.; Stoeck, T. Fungal diversity in oxygen-depleted regions of the Arabian Sea revealed by targeted environmental sequencing combined with cultivation. FEMS Microbiol. Ecol. 2010, 71, 399–412. [Google Scholar] [CrossRef] [Green Version]
- Dell’Anno, A.; Danovaro, R. Ecology: Extracellular DNA plays a key role in deep-sea ecosystem functioning. Science 2005, 309, 2179. [Google Scholar] [CrossRef]
- Dell’Anno, A.; Pusceddu, A.; Corinaldesi, C.; Canals, M.; Heussner, S.; Thomsen, L.; Danovaro, R. Trophic state of benthic deep-sea ecosystems from two different continental margins off Iberia. Biogeosciences 2013, 10, 2945–2957. [Google Scholar] [CrossRef] [Green Version]
- Grinhut, T.; Hadar, Y.; Chen, Y. Degradation and transformation of humic substances by saprotrophic fungi: Processes and mechanisms. Fungal Biol. Rev. 2007, 21, 179–189. [Google Scholar] [CrossRef]
- Raghukumar, S. Animals in Coastal Benthic Ecosystem and Aquaculture Systems. In Fungi in Coastal and Oceanic Marine Ecosystems; Raghukumar, S., Ed.; Springer International Publishing: Cham, Germany, 2017; pp. 163–183. ISBN 978-3-319-54304-8. [Google Scholar]
- Doney, S.C.; Ruckelshaus, M.; Emmett Duffy, J.; Barry, J.P.; Chan, F.; English, C.A.; Galindo, H.M.; Grebmeier, J.M.; Hollowed, A.B.; Knowlton, N.; et al. Climate change impacts on marine ecosystems. Ann. Rev. Mar. Sci. 2012, 4, 11–37. [Google Scholar] [CrossRef] [Green Version]
- Garciá Molinos, J.; Halpern, B.S.; Schoeman, D.S.; Brown, C.J.; Kiessling, W.; Moore, P.J.; Pandolfi, J.M.; Poloczanska, E.S.; Richardson, A.J.; Burrows, M.T. Climate velocity and the future global redistribution of marine biodiversity. Nat. Clim. Chang. 2016, 6, 83–88. [Google Scholar] [CrossRef]
- Silvano, A.; Foppert, A.; Rintoul, S.R.; Holland, P.R.; Tamura, T.; Kimura, N.; Castagno, P.; Falco, P.; Budillon, G.; Haumann, F.A.; et al. Recent recovery of Antarctic Bottom Water formation in the Ross Sea driven by climate anomalies. Nat. Geosci. 2020, 13, 780–786. [Google Scholar] [CrossRef]
- Learman, D.R.; Henson, M.W.; Thrash, J.C.; Temperton, B.; Brannock, P.M.; Santos, S.R.; Mahon, A.R.; Halanych, K.M. Biogeochemical and microbial variation across 5500 km of Antarctic surface sediment implicates organic matter as a driver of benthic community structure. Front. Microbiol. 2016, 7, 284. [Google Scholar] [CrossRef]
- Sweetman, A.K.; Thurber, A.R.; Smith, C.R.; Levin, L.A.; Mora, C.; Wei, C.L.; Gooday, A.J.; Jones, D.O.B.; Rex, M.; Yasuhara, M.; et al. Major impacts of climate change on deep-sea benthic ecosystems. Elementa 2017, 5, 4. [Google Scholar] [CrossRef]
- Smith, W.O.; Ainley, D.G.; Arrigo, K.R.; Dinniman, M.S. The oceanography and ecology of the ross sea. Ann. Rev. Mar. Sci. 2014, 6, 469–487. [Google Scholar] [CrossRef] [Green Version]
- Fabiano, M.; Danovaro, R. Enzymatic activity, bacterial distribution, and organic matter composition in sediments of the Ross Sea (Antarctica). Appl. Environ. Microbiol. 1998, 64, 3838–3845. [Google Scholar] [CrossRef] [Green Version]
- Dell’Anno, A.; Mei, M.L.; Pusceddu, A.; Danovaro, R. Assessing the trophic state and eutrophication of coastal marine systems: A new approach based on the biochemical composition of sediment organic matter. Mar. Pollut. Bull. 2002, 44, 611–622. [Google Scholar] [CrossRef]
- Pusceddu, A.; Dell’Anno, A.; Fabiano, M.; Danovaro, R.; Dell’Anno, A.; Fabiano, M.; Danovaro, R. Quantity and bioavailability of sediment organic matter as signatures of benthic trophic status. Mar. Ecol. Prog. Ser. 2009, 375, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Danovaro, R. Methods for the Study of Deep-Sea Sediments, Their Functioning and Biodiversity; Danovaro, R., Ed.; CRC Press: Boca Raton, FL, USA, 2010; ISBN 9781439811382. [Google Scholar]
- Taylor, J.D.; Cunliffe, M. Multi-year assessment of coastal planktonic fungi reveals environmental drivers of diversity and abundance. ISME J. 2016, 10, 2118–2128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chemidlin Prévost-Bouré, N.; Christen, R.; Dequiedt, S.; Mougel, C.; Lelièvre, M.; Jolivet, C.; Shahbazkia, H.R.; Guillou, L.; Arrouays, D.; Ranjard, L. Validation and application of a PCR primer set to quantify fungal communities in the soil environment by real-time quantitative PCR. PLoS ONE 2011, 6, e24166. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes–application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Rivers, A.R.; Weber, K.C.; Gardner, T.G.; Liu, S.; Armstrong, S.D. ITSxpress: Software to rapidly trim internally transcribed spacer sequences with quality scores for marker gene analysis. F1000Research 2018, 7, 1418. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Accuracy of taxonomy prediction for 16S rRNA and fungal ITS sequences. PeerJ 2018, 2018, e4652. [Google Scholar] [CrossRef]
- Hughes, J.B.; Hellmann, J.J. The application of rarefaction techniques to molecular inventories of microbial diversity. Methods Enzymol. 2005, 397, 292–308. [Google Scholar] [PubMed]
- Gihring, T.M.; Green, S.J.; Schadt, C.W. Massively parallel rRNA gene sequencing exacerbates the potential for biased community diversity comparisons due to variable library sizes. Environ. Microbiol. 2012, 14, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- Anderson, M.J. Permutation tests for univariate or multivariate analysis of variance and regression. Can. J. Fish. Aquat. Sci. 2001, 58, 626–639. [Google Scholar] [CrossRef]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An open source software for exploring and manipulating networks. In Proceedings of the Third International AAAI Conference on Weblogs and Social Media, San Jose, CA, USA, 17–20 May 2009. [Google Scholar]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- McArdle, B.H.; Anderson, M.J. Fitting Multivariate Models to Community Data: A Comment on Distance-Based Redundancy Analysis. Ecology 2001, 82, 290. [Google Scholar] [CrossRef]
- Bergamasco, A.; Defendi, V.; Zambianchi, E.; Spezie, G. Evidence of dense water overflow on the Ross Sea shelf-break. Antarct. Sci. 2002, 14, 271–277. [Google Scholar] [CrossRef]
- Gardner, W.D.; Richardson, M.J.; Smith, W.O. Seasonal patterns of water column particulate organic carbon and fluxes in the ross sea, antarctica. Deep. Res. Part II Top. Stud. Oceanogr. 2000, 47, 3423–3449. [Google Scholar] [CrossRef]
- Frignani, M.; Giglio, F.; Accornero, A.; Langone, L.; Ravaioli, M. Sediment characteristics at selected sites of the Ross Sea continental shelf: Does the sedimentary record reflect water column fluxes? Antarct. Sci. 2003, 15, 133–139. [Google Scholar] [CrossRef]
- Danovaro, R.; Molari, M.; Corinaldesi, C.; Dell’Anno, A. Macroecological drivers of archaea and bacteria in benthic deep-sea ecosystems. Sci. Adv. 2016, 2, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, C.L.; Rowe, G.T.; Briones, E.E.; Boetius, A.; Soltwedel, T.; Caley, M.J.; Soliman, Y.; Huettmann, F.; Qu, F.; Yu, Z.; et al. Global patterns and predictions of seafloor biomass using random forests. PLoS ONE 2010, 5, e15323. [Google Scholar] [CrossRef]
- Xu, W.; Pang, K.L.; Luo, Z.H. High Fungal Diversity and Abundance Recovered in the Deep-Sea Sediments of the Pacific Ocean. Microb. Ecol. 2014, 68, 688–698. [Google Scholar] [CrossRef]
- Rämä, T.; Hassett, B.T.; Bubnova, E. Arctic marine fungi: From filaments and flagella to operational taxonomic units and beyond. Bot. Mar. 2017, 60, 433–452. [Google Scholar] [CrossRef]
- Duarte, A.W.F.; dos Santos, J.A.; Vianna, M.V.; Vieira, J.M.F.; Mallagutti, V.H.; Inforsato, F.J.; Wentzel, L.C.P.; Lario, L.D.; Rodrigues, A.; Pagnocca, F.C.; et al. Cold-adapted enzymes produced by fungi from terrestrial and marine Antarctic environments. Crit. Rev. Biotechnol. 2018, 38, 600–619. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.; Xue, C.; Tian, L.; Xu, M.; Chen, J.; Deng, Z.; Proksch, P.; Lin, W. Asperelines A−F, Peptaibols from the Marine-Derived Fungus Trichoderma asperellum. J. Nat. Prod. 2009, 72, 1036–1044. [Google Scholar] [CrossRef]
- Gonçalves, V.N.; Vaz, A.B.M.; Rosa, C.A.; Rosa, L.H. Diversity and distribution of fungal communities in lakes of Antarctica. FEMS Microbiol. Ecol. 2012, 82, 459–471. [Google Scholar] [CrossRef] [Green Version]
- Godinho, V.M.; Furbino, L.E.; Santiago, I.F.; Pellizzari, F.M.; Yokoya, N.S.; Pupo, D.; Alves, T.M.A.; S Junior, P.A.; Romanha, A.J.; Zani, C.L.; et al. Diversity and bioprospecting of fungal communities associated with endemic and cold-adapted macroalgae in Antarctica. ISME J. 2013, 7, 1434–1451. [Google Scholar] [CrossRef] [PubMed]
- Furbino, L.E.; Godinho, V.M.; Santiago, I.F.; Pellizari, F.M.; Alves, T.M.A.A.; Zani, C.L.; Junior, P.A.S.S.; Romanha, A.J.; Carvalho, A.G.O.O.; Gil, L.H.V.G.V.G.; et al. Diversity Patterns, Ecology and Biological Activities of Fungal Communities Associated with the Endemic Macroalgae Across the Antarctic Peninsula. Microb. Ecol. 2014, 67, 775–787. [Google Scholar] [CrossRef]
- Wang, F.; Sheng, J.; Chen, Y.; Xu, J. Microbial diversity and dominant bacteria causing spoilage during storage and processing of the Antarctic krill, Euphausia superba. Polar Biol. 2021, 44, 163–171. [Google Scholar] [CrossRef]
- Gonçalves, V.N.; Vitoreli, G.A.; de Menezes, G.C.A.; Mendes, C.R.B.; Secchi, E.R.; Rosa, C.A.; Rosa, L.H. Taxonomy, phylogeny and ecology of cultivable fungi present in seawater gradients across the Northern Antarctica Peninsula. Extremophiles 2017, 21, 1005–1015. [Google Scholar] [CrossRef] [PubMed]
- Richards, T.A.; Talbot, N.J. Horizontal gene transfer in osmotrophs: Playing with public goods. Nat. Rev. Microbiol. 2013, 11, 720–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, T.A.; Leonard, G.; Mahé, F.; Del Campo, J.; Romac, S.; Jones, M.D.M.; Maguire, F.; Dunthorn, M.; De Vargas, C.; Massana, R.; et al. Molecular diversity and distribution of marine fungi across 130 european environmental samples. Proc. R. Soc. B Biol. Sci. 2015, 282, 20152243. [Google Scholar] [CrossRef] [PubMed]
- Couturier, M.; Bennati-Granier, C.; Urio, M.B.; Ramos, L.P.; Berrin, J.G. Fungal enzymatic degradation of cellulose. In Green Fuels Technology; Green Energy and Technology; Soccol, C., Brar, S., Faulds, C., Ramos, L., Eds.; Springer: Cham, Switzerland, 2016; pp. 133–146. [Google Scholar]
- Fabiano, M.; Danovaro, R. Meiofauna distribution and mesoscale variability in two sites of the Ross Sea (Antarctica) with contrasting food supply. Polar Biol. 1999, 22, 115–123. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Station | Depth | Latitude | Longitude | Temperature | Salinity | Dissolved Oxygen |
|---|---|---|---|---|---|---|---|
| (m) | EPSG: 4326 | EPSG: 4326 | (°C) | (mg L−1) | |||
| B | B1 | 585 | −74.03951 | 175.07945 | −1.88 ± 0.0002 | 34.752 ± 0.0002 | 10.96 ± 0.05 |
| B | B2 | 587 | −74.01603 | 175.04279 | −1.878 ± 0.0002 | 34.756 ± 0.0001 | 11 ± 0.05 |
| C | C1 | 433 | −72.49527 | 174.94336 | −0.5 ± 0.001 | 34.65 ± 0.0002 | 6.25 ± 0.003 |
| C | C2 | 434 | −72.49967 | 174.99696 | −0.046 ± 0.0024 | 34.667 ± 0.0001 | 9.92 ± 0.005 |
| Biopolymeric Carbon | Total phytopigments | P:C | |||||
| (mg g−1) | (µg g−1) | ||||||
| B | B1 | 3.23 ± 0.23 | 3.28 ± 1.27 | 0.39 ± 0.1 | |||
| B | B2 | 2.32 ± 0.61 | 3.44 ± 0.84 | 0.19 ± 0.1 | |||
| C | C1 | 0.28 ± 0.08 | 0.33 ± 0.08 | 0.7 ± 0.19 | |||
| C | C2 | 0.23 ± 0.08 | 0.24 ± 0.11 | 0.46 ± 0.06 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barone, G.; Corinaldesi, C.; Rastelli, E.; Tangherlini, M.; Varrella, S.; Danovaro, R.; Dell’Anno, A. Local Environmental Conditions Promote High Turnover Diversity of Benthic Deep-Sea Fungi in the Ross Sea (Antarctica). J. Fungi 2022, 8, 65. https://doi.org/10.3390/jof8010065
Barone G, Corinaldesi C, Rastelli E, Tangherlini M, Varrella S, Danovaro R, Dell’Anno A. Local Environmental Conditions Promote High Turnover Diversity of Benthic Deep-Sea Fungi in the Ross Sea (Antarctica). Journal of Fungi. 2022; 8(1):65. https://doi.org/10.3390/jof8010065
Chicago/Turabian StyleBarone, Giulio, Cinzia Corinaldesi, Eugenio Rastelli, Michael Tangherlini, Stefano Varrella, Roberto Danovaro, and Antonio Dell’Anno. 2022. "Local Environmental Conditions Promote High Turnover Diversity of Benthic Deep-Sea Fungi in the Ross Sea (Antarctica)" Journal of Fungi 8, no. 1: 65. https://doi.org/10.3390/jof8010065