
Extracellular Enzymatic Activities of Oceanic Pelagic Fungal Strains and the Influence of Temperature


Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture of Fungi Species
2.2. Determining Extracellular Enzymatic Activity and Fungal Biomass
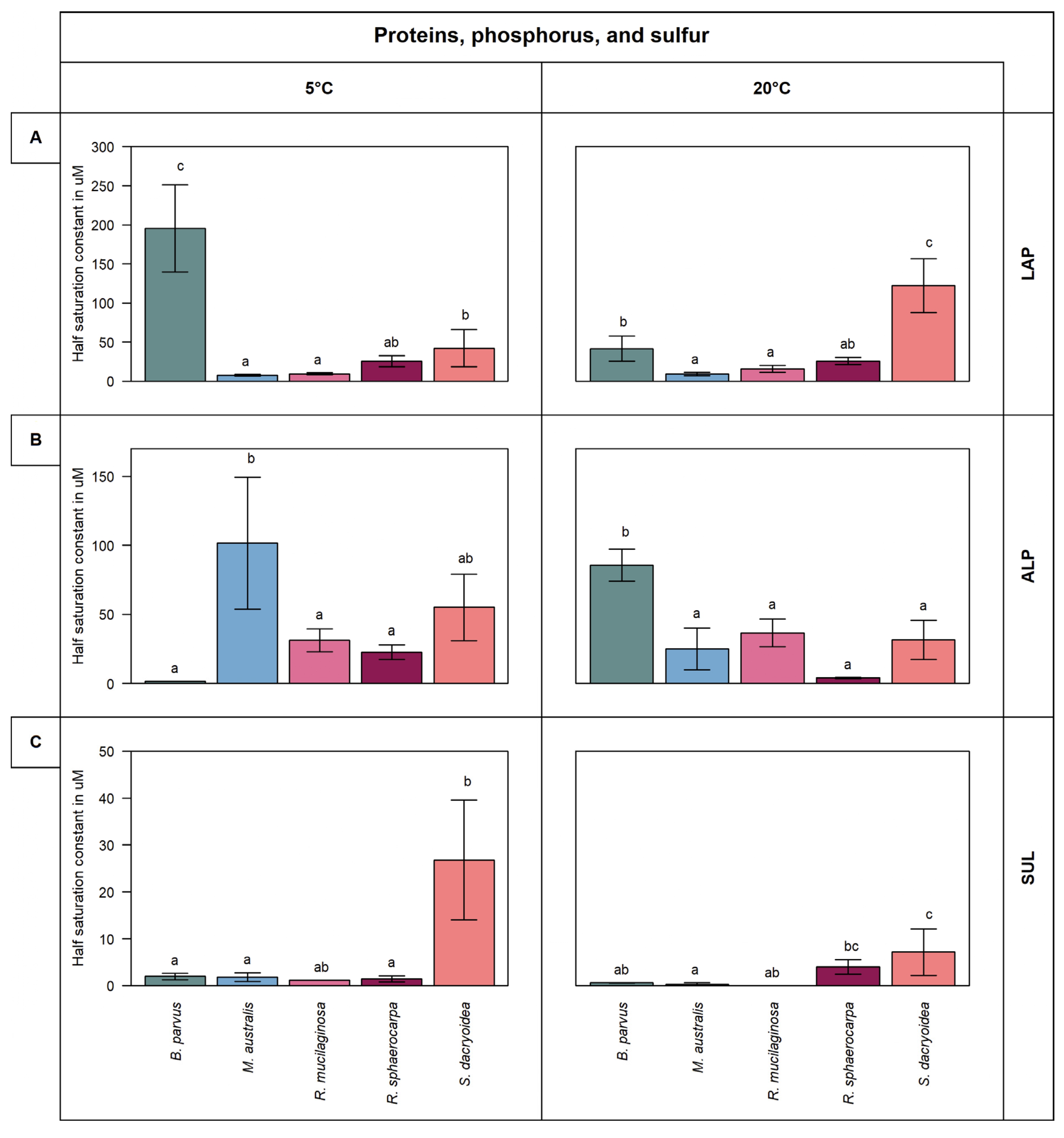
2.3. Determination of the Kinetic Parameters of the Extracellular Enzymatic Activity (EEA)
2.4. Statistical Analyses
3. Results
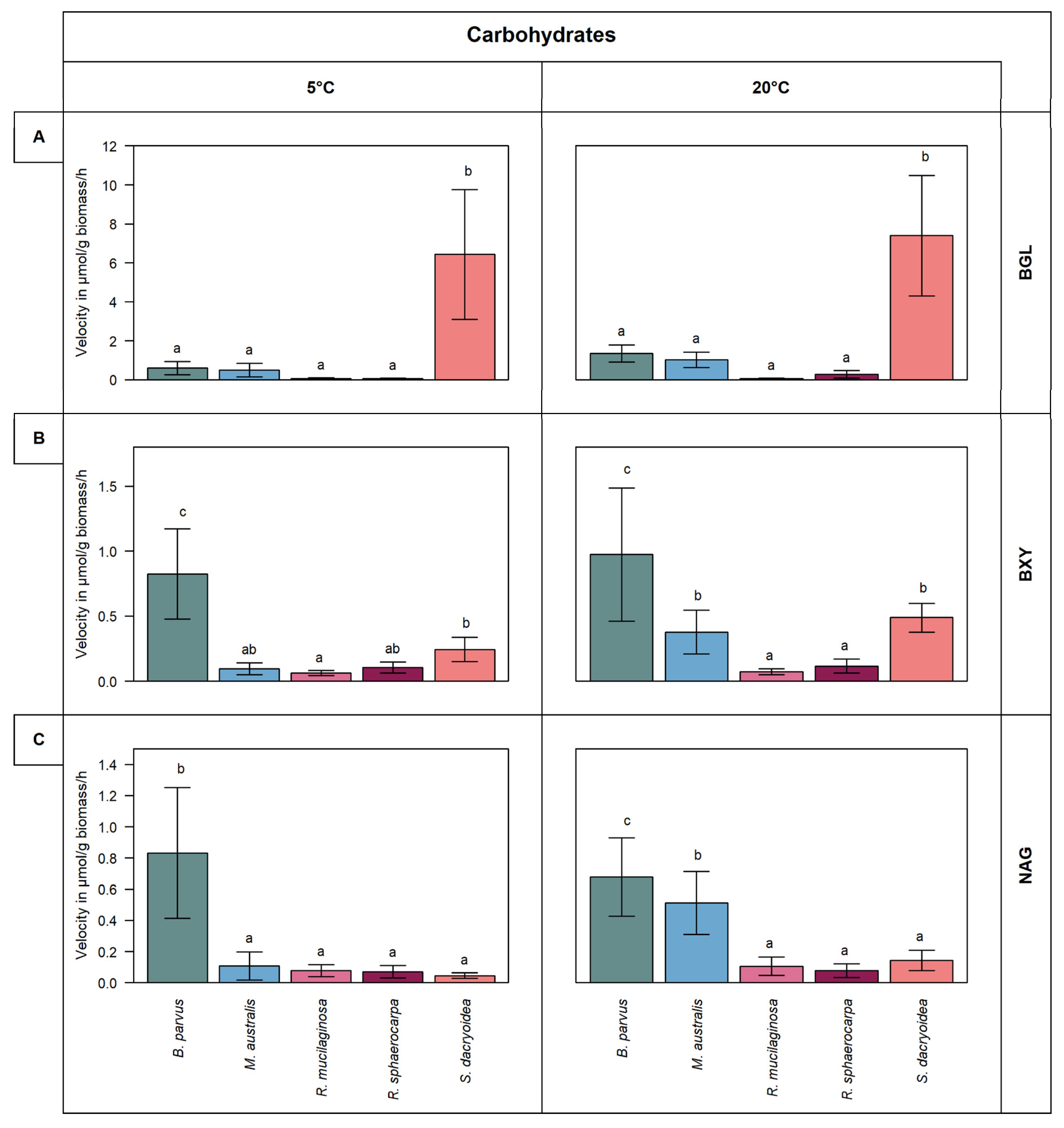
3.1. Carbohydrate Active Enzymes
3.1.1. β-Glucosidase (BGL)
3.1.2. β-Xylosidase (BXY)
3.1.3. N-acetyl-β-D-glucosaminidase (NAG)
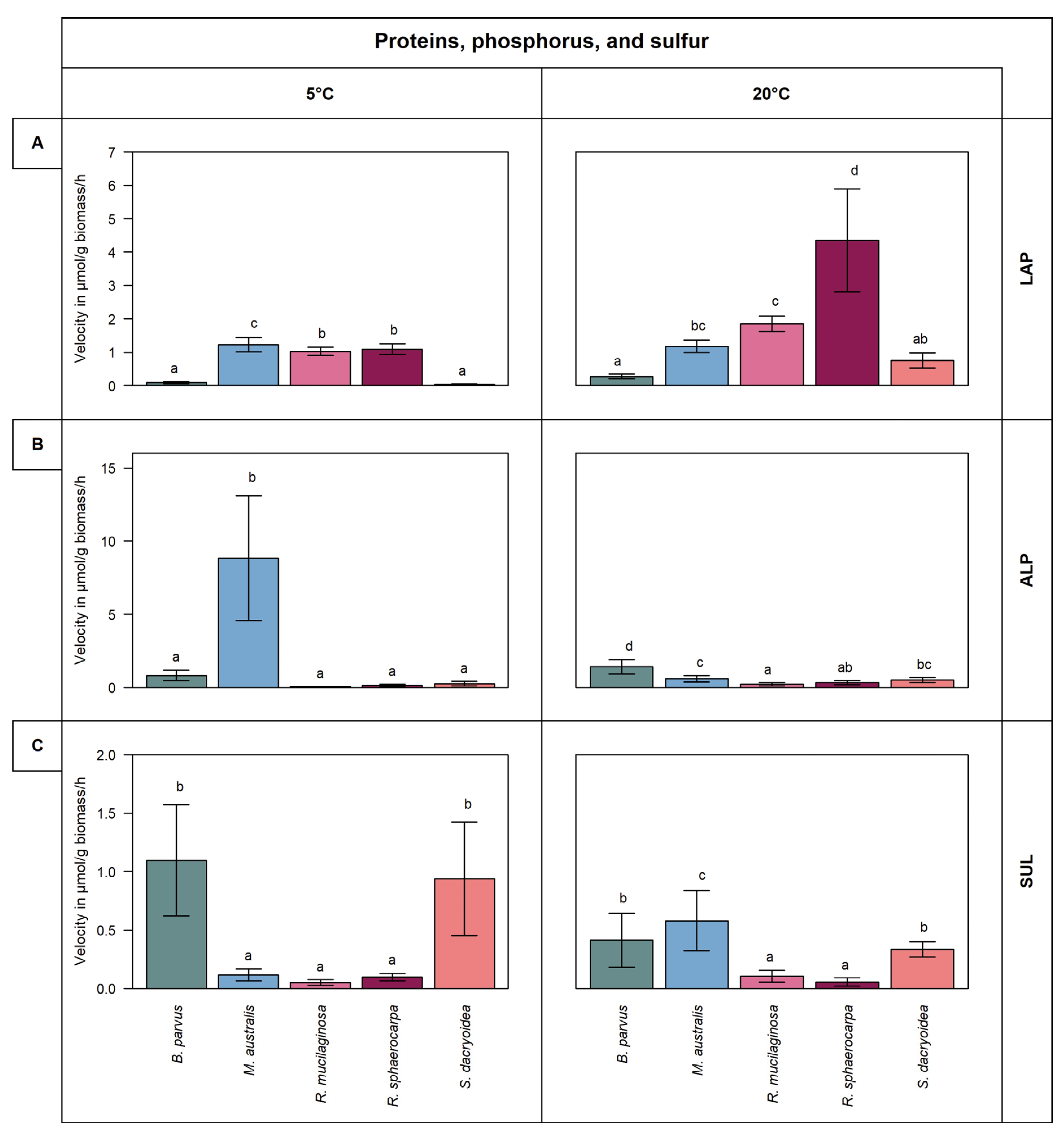
3.2. Extracellular Enzymes Targeting Proteins, Phosphorus, and Sulfur Compounds
3.2.1. Leucine Aminopeptidase (LAP)
3.2.2. Alkaline Phosphatase (ALP)
3.2.3. Sulfatase (SUL)
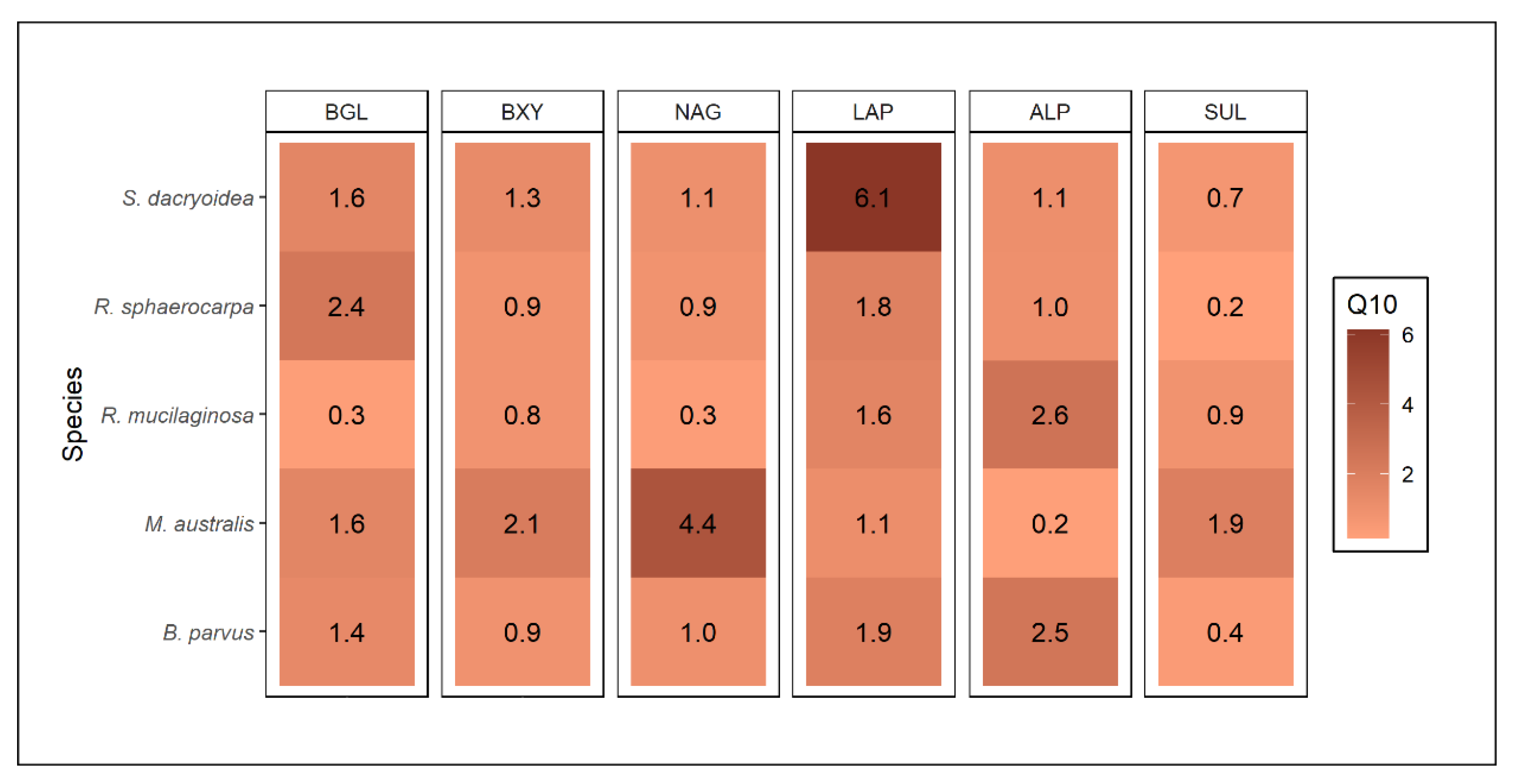
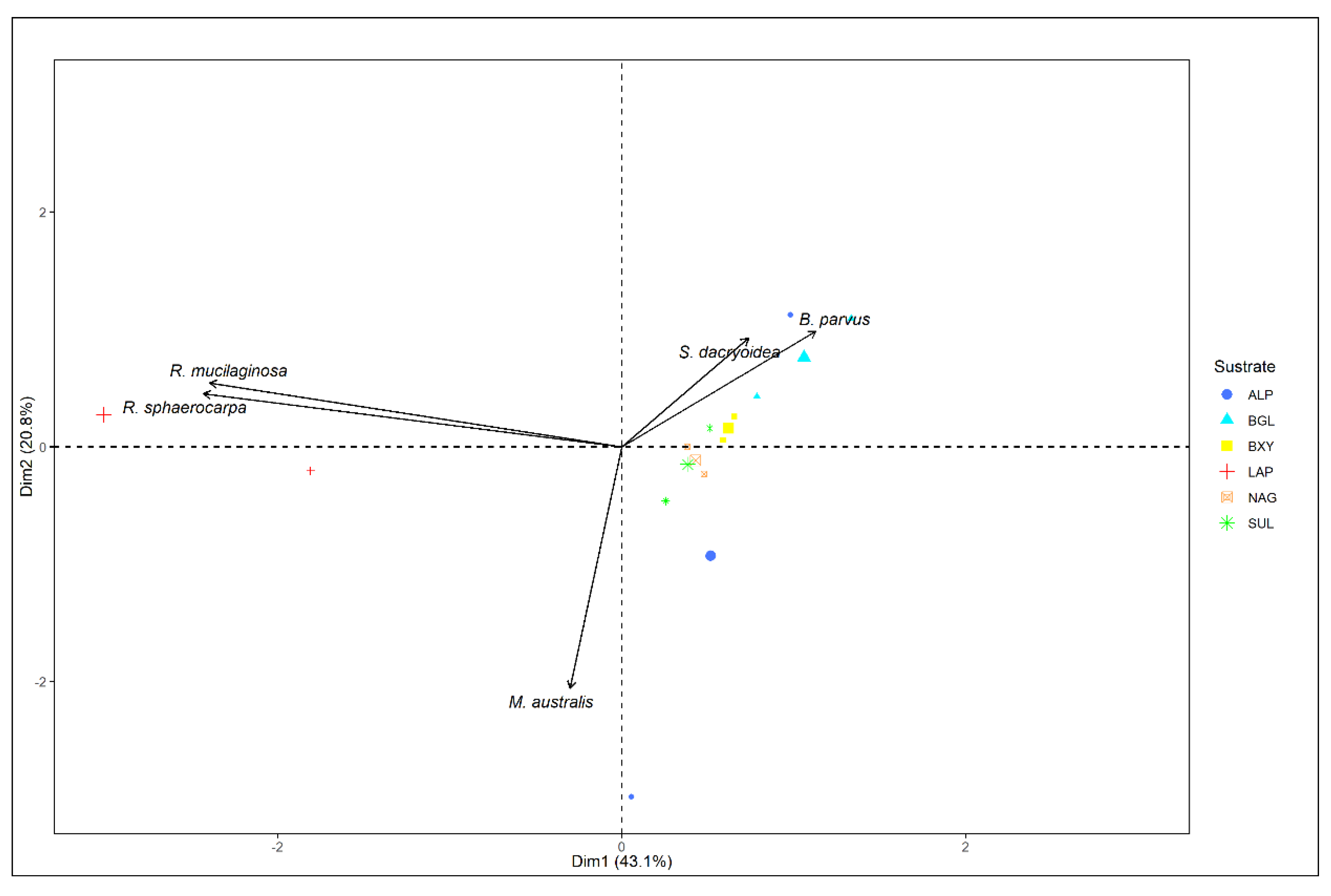
3.3. Relation between Enzyme Kinetic Parameters, Enzyme Types, and Phylogeny
4. Discussion
4.1. Extracellular Enzymatic Activities of Pelagic Fungal Isolates
4.1.1. Influence of Taxonomy/Diversity on the Different EEAs
4.1.2. Temperature Influence
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nevalainen, H.; Kautto, L.; Te’o, J. Methods for isolation and cultivation of filamentous fungi. Methods Mol. Biol. 2014, 1096, 3–16. [Google Scholar]
- Richards, T.A.; Jones, M.D.; Leonard, G.; Bass, D. Marine fungi: Their ecology and molecular diversity. Annu. Rev. Mar. Sci. 2012, 4, 495–522. [Google Scholar] [CrossRef] [PubMed]
- Arrieta, J.M.; Herndl, G.J. Changes in bacterial β-glucosidase diversity during a coastal phytoplankton bloom. Limnol. Oceanogr. 2002, 47, 594–599. [Google Scholar] [CrossRef]
- Baltar, F.; Arístegui, J.; Sintes, E.; Van Aken, H.M.; Gasol, J.M.; Herndl, G.J. Prokaryotic extracellular enzymatic activity in relation to biomass production and respiration in the meso-and bathypelagic waters of the (sub) tropical Atlantic. Environ. Microbiol. 2009, 11, 1998–2014. [Google Scholar] [CrossRef]
- Gutiérrez, M.H.; Pantoja, S.; Tejos, E.; Quiñones, R.A. The role of fungi in processing marine organic matter in the upwelling ecosystem off Chile. Mar. Biol. 2011, 158, 205–219. [Google Scholar] [CrossRef]
- Cunliffe, M.; Hollingsworth, A.; Bain, C.; Sharma, V.; Taylor, J.D. Algal polysaccharide utilisation by saprotrophic planktonic marine fungi. Fungal Ecol. 2017, 30, 135–138. [Google Scholar] [CrossRef]
- Gao, Z.; Johnson, Z.I.; Wang, G. Molecular characterization of the spatial diversity and novel lineages of mycoplankton in Hawaiian coastal waters. ISME J. 2010, 4, 111–120. [Google Scholar] [CrossRef]
- Bochdansky, A.B.; Clouse, M.A.; Herndl, G.J. Eukaryotic microbes, principally fungi and labyrinthulomycetes, dominate biomass on bathypelagic marine snow. ISME J. 2017, 11, 362–373. [Google Scholar] [CrossRef]
- Morales, S.E.; Biswas, A.; Herndl, G.J.; Baltar, F. Global Structuring of Phylogenetic and Functional Diversity of Pelagic Fungi by Depth and Temperature. Front. Mar. Sci. 2019, 6, 131. [Google Scholar] [CrossRef] [Green Version]
- Baltar, F.; Zhao, Z.; Herndl, G.J. Potential and expression of carbohydrate utilization by marine fungi in the global ocean. Microbiome 2021, 9, 106. [Google Scholar] [CrossRef]
- D’Amico, S.; Collins, T.; Marx, J.C.; Feller, G.; Gerday, C.; Gerday, C. Psychrophilic microorganisms: Challenges for life. EMBO Rep. 2006, 7, 385–389. [Google Scholar] [CrossRef] [PubMed]
- Yusof, N.A.; Hashim, N.H.F.; Bharudin, I. Cold adaptation strategies and the potential of psychrophilic enzymes from the antarctic yeast, Glaciozyma antarctica PI12. J. Fungi 2021, 7, 528. [Google Scholar] [CrossRef] [PubMed]
- Morita, R.Y. Psychrophilic bacteria. Bacteriol. Rev. 1975, 39, 144–167. [Google Scholar] [CrossRef] [PubMed]
- Feller, G.; Gerday, C. Psychrophilic enzymes: Hot topics in cold adaptation. Nat. Rev. Microbiol. 2003, 1, 200–208. [Google Scholar] [CrossRef]
- Vonnahme, T.R.; Dietrich, U.; Hassett, B.T. Progress in microbial ecology in ice-covered seas. In YOUMARES 9—The Oceans: Our Research, Our Future; Springer: Cham, Switzerland, 2020; p. 261. [Google Scholar]
- Amend, A.; Burgaud, G.; Cunliffe, M.; Edgcomb, V.P.; Ettinger, C.L.; Gutiérrez, M.H.; Heitman, J.; Hom, E.F.Y.; Ianiri, G.; Jones, A.C.; et al. Fungi in the Marine Environment: Open Questions and Unsolved Problems. mBio 2019, 10, e01189-18. [Google Scholar] [CrossRef] [Green Version]
- Taylor, J.D.; Cunliffe, M. Multi-year assessment of coastal planktonic fungi reveals environmental drivers of diversity and abundance. ISME J. 2016, 10, 2118–2128. [Google Scholar] [CrossRef] [Green Version]
- Fell, J.; Statzell, A.C. Sympodiomyces gen. n., a yeast-like organism from southern marine waters. Antonie Van Leeuwenhoek 1971, 37, 359–367. [Google Scholar] [CrossRef]
- Fell, J.W.; Hunter, I.L. Isolation of heterothallic yeast strains of Metschnikowia Kamienski and their mating reactions with Chlamydozyma wickerham spp. Antonie Van Leeuwenhoek 1968, 34, 365–376. [Google Scholar] [CrossRef]
- Fell, J.W.; Hunter, I.L.; Tallman, A.S. Marine basidiomycetous yeasts (Rhodosporidium spp. n.) with tetrapolar and multiple allelic bipolar mating systems. Can. J. Microbiol. 1973, 19, 643–657. [Google Scholar] [CrossRef]
- Newell, S.Y.; Fell, J. The perfect form of a marine-occurring yeast of the genus Rhodotorula. Mycologia 1970, 62, 272–281. [Google Scholar] [CrossRef]
- Sampaio, J.P. Rhodotorula Harrison (1928). In The Yeasts; Elsevier: Burlington, VT, USA, 2011; pp. 1873–1927. [Google Scholar]
- Wickerham, L.J. A taxonomic study of Monilia albicans with special emphasis on morphology and morphological variation. J. Tropical Med. Hyg. 1939, 42, 174–179. [Google Scholar]
- Wickerham, L.J. Taxonomy of Yeasts; US Department of Agriculture: Washington, DC, USA, 1951.
- Salazar Alekseyeva, K.; Mähnert, B.; Berthiller, F.; Breyer, E.; Herndl, G.J.; Baltar, F. Adapting an Ergosterol Extraction Method with Marine Yeasts for the Quantification of Oceanic Fungal Biomass. J. Fungi 2021, 7, 690. [Google Scholar] [CrossRef]
- Norkrans, B. Degradation of cellulose. Annu. Rev. Phytopathol. 1963, 1, 325–350. [Google Scholar] [CrossRef]
- Pointing, S.B.; Vrijmoed, L.L.P.; Jones, E.B.G. A Qualitative Assessment of Lignocellulose Degrading Enzyme Activity in Marine Fungi. Bot. Mar. 1998, 41, 293–298. [Google Scholar] [CrossRef]
- Collins, T.; Gerday, C.; Feller, G. Xylanases, xylanase families and extremophilic xylanases. FEMS Microbiol. Rev. 2005, 29, 3–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- dos Santos, J.A.; Vieira, J.M.F.; Videira, A.; Meirelles, L.A.; Rodrigues, A.; Taniwaki, M.H.; Sette, L.D. Marine-derived fungus Aspergillus cf. tubingensis LAMAI 31: A new genetic resource for xylanase production. AMB Express 2016, 6, 25. [Google Scholar]
- Hoppe, H.-G. Significance of exoenzymatic activities in the ecology of brackish water: Measurements by means of methylumbelliferyl-substrates. Mar. Ecol. Prog. Ser. 1983, 11, 299–308. [Google Scholar] [CrossRef]
- Huitema, C.; Horsman, G. Analyzing Enzyme Kinetic Data Using the Powerful Statistical Capabilities of R. bioRxiv 2018, 1, 316588. [Google Scholar]
- Sterratt, D.C. Q10: The Effect of Temperature on Ion Channel Kinetics, in Encyclopedia of Computational Neuroscience; Jaeger, D., Jung, R., Eds.; Springer: New York, NY, USA, 2015; pp. 2551–2552. [Google Scholar]
- Arnosti, C.; Bell, C.; Moorhead, D.L.; Sinsabaugh, R.L.; Steen, A.D.; Stromberger, M.; Wallenstein, M.; Weintraub, M.N. Extracellular enzymes in terrestrial, freshwater, and marine environments: Perspectives on system variability and common research needs. Biogeochemistry 2014, 117, 5–21. [Google Scholar] [CrossRef]
- Cullings, K.; Courty, P.-E. Saprotrophic capabilities as functional traits to study functional diversity and resilience of ectomycorrhizal community. Oecologia 2009, 161, 661–664. [Google Scholar] [CrossRef]
- Helbert, W. Marine Polysaccharide Sulfatases. Front. Mar. Sci. 2017, 4, 6. [Google Scholar] [CrossRef] [Green Version]
- Metreveli, E.; Khardziani, T.; Elisashvili, V. The Carbon Source Controls the Secretion and Yield of Polysaccharide-Hydrolyzing Enzymes of Basidiomycetes. Biomolecules 2021, 11, 1341. [Google Scholar] [CrossRef]
- Houtman, C.J.; Atalla, R.H. Cellulose-Lignin Interactions (A Computational Study). Plant Physiol. 1995, 107, 977–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domozych, D.; Ciancia, M.; Fangel, J.; Mikkelsen, M.; Ulvskov, P.; Willats, W. The Cell Walls of Green Algae: A Journey through Evolution and Diversity. Front. Plant Sci. 2012, 3, 82. [Google Scholar] [CrossRef] [Green Version]
- Arora, D.K. Handbook of Applied Mycology: Volume 1: Soil and Plants; CRC Press: Boca Raton, FL, USA, 1991. [Google Scholar]
- Hong, J.-H.; Jang, S.; Heo, Y.M.; Min, M.; Lee, H.; Lee, Y.M.; Lee, H.; Kim, J.-J. Investigation of Marine-Derived Fungal Diversity and Their Exploitable Biological Activities. Mar. Drugs 2015, 13, 4137–4155. [Google Scholar] [CrossRef] [PubMed]
- Meyers, S.; Scott, E. Cellulose degradation by Lulworthia floridana and other lignicolous marine fungi. Mar. Biol. 1968, 2, 41–46. [Google Scholar] [CrossRef]
- Baraldo Junior, A.; Borges, D.G.; Tardioli, P.W.; Farinas, C.S. Characterization of β-glucosidase produced by Aspergillus niger under solid-state fermentation and partially purified using MANAE-Agarose. Biotechnol. Res. Int. 2014, 2014, 317092. [Google Scholar] [CrossRef] [Green Version]
- Ab Wahab, A.F.F.; Karim, N.A.A.; Ling, J.G.; Hasan, N.S.; Yong, H.Y.; Bharudin, I.; Kamaruddin, S.; Bakar, F.D.A.; Murad, A.M. Functional characterisation of cellobiohydrolase I (Cbh1) from Trichoderma virens UKM1 expressed in Aspergillus niger. Protein Expr. Purif. 2019, 154, 52–61. [Google Scholar] [CrossRef]
- Vaz, A.B.M.; Rosa, L.H.; Vieira, M.L.A.; de Garcia, V.; Brandão, L.R.; Teixeira, L.C.R.S.; Moliné, M.; Libkind, D.; van Broock, M.; Rosa, C. The diversity, extracellular enzymatic activities and photoprotective compounds of yeasts isolated in Antarctica. Braz. J. Microbiol. 2011, 42, 937–947. [Google Scholar] [CrossRef]
- Hudson, H.J. Fungal Saprophytism; Edward Arnold: London, UK, 1972. [Google Scholar]
- Roussel, M.R. A Life Scientist’s Guide to Physical Chemistry; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Balabanova, L.; Slepchenko, L.; Son, O.; Tekutyeva, L. Biotechnology Potential of Marine Fungi Degrading Plant and Algae Polymeric Substrates. Front. Microbiol. 2018, 9, 1527. [Google Scholar] [CrossRef] [Green Version]
- Goddard-Borger, E.D.; Sakaguchi, K.; Reitinger, S.; Watanabe, N.; Ito, M.; Withers, S.G. Mechanistic insights into the 1,3-xylanases: Useful enzymes for manipulation of algal biomass. J. Am. Chem. Soc. 2012, 134, 3895–3902. [Google Scholar] [CrossRef] [PubMed]
- Synytsya, A.; Čopíková, J.; Kim, W.J.; Park, Y.I. Cell Wall Polysaccharides of Marine Algae. In Springer Handbook of Marine Biotechnology; Kim, S.-K., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 543–590. [Google Scholar]
- Domingues, R.; Bondar, M.; Palolo, I.; Queirós, O.; de Almeida, C.D.; Cesário, M.T. Xylose Metabolism in Bacteria—Opportunities and Challenges towards Efficient Lignocellulosic Biomass-Based Biorefineries. Appl. Sci. 2021, 11, 8112. [Google Scholar] [CrossRef]
- Raghukumar, C.; Raghukumar, S.; Chinnaraj, A.; Chandramohan, D.; D’souza, T.; Reddy, C. Laccase and Other Lignocellulose Modifying Enzymes of Marine Fungi Isolated from the Coast of India. Botanica Marina. 1994, 37, 515–523. [Google Scholar] [CrossRef]
- Duarte, A.W.F.; Dayo-Owoyemi, I.; Nobre, F.; Pagnocca, F.C.; Chaud, L.C.S.; Pessoa, A.; Felipe, M.G.A.; Sette, L.D. Taxonomic assessment and enzymes production by yeasts isolated from marine and terrestrial Antarctic samples. Extremophiles 2013, 17, 1023–1035. [Google Scholar] [CrossRef] [PubMed]
- Numata, K.; Kaplan, D.L. 20—Biologically derived scaffolds. In Advanced Wound Repair Therapies; Farrar, D., Ed.; Woodhead Publishing: Sawston, UK, 2011; pp. 524–551. [Google Scholar]
- Ruiz-Herrera, J. Fungal Cell Wall: Structure, Synthesis, and Assembly; CRC Press: Boca Raton, FL, USA, 1991. [Google Scholar]
- Fenice, M. The psychrotolerant Antarctic fungus Lecanicillium muscarium CCFEE 5003: A powerful producer of cold-tolerant chitinolytic enzymes. Molecules 2016, 21, 447. [Google Scholar] [CrossRef] [Green Version]
- Fenice, M.; Selbmann, L.; Di Giambattista, R.; Federici, F. Chitinolytic activity at low temperature of an Antarctic strain (A3) of Verticillium lecanii. Res. Microbiol. 1998, 149, 289–300. [Google Scholar] [CrossRef]
- Hartl, L.; Zach, S.; Seidl-Seiboth, V. Fungal chitinases: Diversity, mechanistic properties and biotechnological potential. Appl. Microbiol. Biotechnol. 2012, 93, 533–543. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Rubio, R.; de Oliveira, H.C.; Rivera, J.; Trevijano-Contador, N. The Fungal Cell Wall: Candida, Cryptococcus, and Aspergillus Species. Front. Microbiol. 2020, 10, 2993. [Google Scholar] [CrossRef]
- Burley, S.K.; David, P.R.; Taylor, A.; Lipscomb, W.N. Molecular structure of leucine aminopeptidase at 2.7—A resolution. Proc. Natl. Acad. Sci. USA 1990, 87, 6878–6882. [Google Scholar] [CrossRef] [Green Version]
- Nampoothiri, K.M.; Nagy, V.; Kovacs, K.; Szakacs, G.; Pandey, A. L-leucine aminopeptidase production by filamentous Aspergillus fungi. Lett. Appl. Microbiol. 2005, 41, 498–504. [Google Scholar] [CrossRef]
- Karl, D.M. Phosphorus, the staff of life. Nature 2000, 406, 31–33. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.; Saavedra, D.E.M.; Thomson, B.; García, J.A.L.; Zhao, Z.; Patrick, W.M.; Herndl, G.J.; Baltar, F. Enzyme promiscuity in natural environments: Alkaline phosphatase in the ocean. ISME J. 2021, 15, 3375–3383. [Google Scholar] [CrossRef] [PubMed]
- Baltar, F.; Lundin, D.; Palovaara, J.; Lekunberri, I.; Reinthaler, T.; Herndl, G.J.; Pinhassi, J. Prokaryotic Responses to Ammonium and Organic Carbon Reveal Alternative CO2 Fixation Pathways and Importance of Alkaline Phosphatase in the Mesopelagic North Atlantic. Front. Microbiol. 2016, 7, 1670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shvetsova, S.V.; Zhurishkina, E.V.; Bobrov, K.S.; Ronzhina, N.L.; Lapina, I.M.; Ivanen, D.R.; Gagkaeva, T.Y.; Kulminskaya, A.A. The novel strain Fusarium proliferatum LE1 (RCAM02409) produces α-l-fucosidase and arylsulfatase during the growth on fucoidan. J. Basic Microbiol. 2015, 55, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Berlemont, R. Distribution and diversity of enzymes for polysaccharide degradation in fungi. Sci. Rep. 2017, 7, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loque, C.P.; Medeiros, A.O.; Pellizzari, F.M.; Oliveira, E.C.; Rosa, C.A.; Rosa, L.H. Fungal community associated with marine macroalgae from Antarctica. Polar Biol. 2010, 33, 641–648. [Google Scholar] [CrossRef]
- Georlette, D.; Blaise, V.; Collins, T.; D’Amico, S.; Gratia, E.; Hoyoux, A.; Marx, J.-C.; Sonan, G.; Feller, G.; Gerday, C. Some like it cold: Biocatalysis at low temperatures. FEMS Microbiol. Rev. 2004, 28, 25–42. [Google Scholar] [CrossRef] [Green Version]
- John, A.B.; Diana, E.V.; Paul, J.H. Effects of temperature on growth rate, cell composition and nitrogen metabolism in the marine diatom Thalassiosira pseudonana (Bacillariophyceae). Mar. Ecol. Prog. Ser. 2002, 225, 139–146. [Google Scholar]
- Collins, T.; D’Amico, S.; Marx, J.C.; Feller, G.; Gerday, C. Cold-Adapted Enzymes. Physiol. Biochem. Extrem. 2007, 1, 165–179. [Google Scholar]
- Kawahara, H.; Iwanaka, Y.; Higa, S.; Muryoi, N.; Sato, M.; Honda, M.; Omura, H.; Obata, H. A novel, intracellular antifreeze protein in an antarctic bacterium, Flavobacterium xanthum. Cryoletters 2007, 28, 39–49. [Google Scholar]
- Krembs, C.; Eicken, H.; Junge, K.; Deming, J. High concentrations of exopolymeric substances in Arctic winter sea ice: Implications for the polar ocean carbon cycle and cryoprotection of diatoms. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2002, 49, 2163–2181. [Google Scholar] [CrossRef]
- Los, D.A.; Murata, N. Membrane fluidity and its roles in the perception of environmental signals. Biochim. Et Biophys. Acta 2004, 1666, 142–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerday, C.; Aittaleb, M.; Bentahir, M.; Chessa, J.-P.; Claverie, P.; Collins, T.; D’Amico, S.; Dumont, J.; Garsoux, G.; Georlette, D. Cold-adapted enzymes: From fundamentals to biotechnology. Trends Biotechnol. 2000, 18, 103–107. [Google Scholar] [CrossRef]
- Feller, G.; Gerday, C. Psychrophilic enzymes: Molecular basis of cold adaptation. Cell. Mol. Life Sci. CMLS 1997, 53, 830–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aghajari, N.; Haser, R.; Feller, G.; Gerday, C. Crystallization and preliminary X-ray diffraction studies of α-amylase from the antarctic psychrophile Alteromonas haloplanctis A23. Protein Sci. 1996, 5, 2128–2129. [Google Scholar] [CrossRef] [Green Version]
- Truongvan, N.; Jang, S.-H.; Lee, C. Flexibility and stability trade-off in active site of cold-adapted Pseudomonas mandelii esterase EstK. Biochemistry 2016, 55, 3542–3549. [Google Scholar] [CrossRef]
- Ramli, A.N.M.; Mahadi, N.M.; Shamsir, M.S.; Rabu, A.; Joyce-Tan, K.H.; Murad, A.M.A.; Illias, R. Structural prediction of a novel chitinase from the psychrophilic Glaciozyma antarctica PI12 and an analysis of its structural properties and function. J. Comput. Aided Mol. Des. 2012, 26, 947–961. [Google Scholar] [CrossRef]
- Baeza, M.; Zúñiga, S.; Peragallo, V.; Barahona, S.; Alcaino, J.; Cifuentes, V. Identification of stress-related genes and a comparative analysis of the amino acid compositions of translated coding sequences based on draft genome sequences of Antarctic yeasts. Front. Microbiol. 2021, 12, 133. [Google Scholar] [CrossRef]
- DasSarma, S.; Capes, M.D.; Karan, R.; DasSarma, P. Amino acid substitutions in cold-adapted proteins from Halorubrum lacusprofundi, an extremely halophilic microbe from Antarctica. PLoS ONE 2013, 8, e58587. [Google Scholar] [CrossRef] [Green Version]
- Michetti, D.; Brandsdal, B.O.; Bon, D.; Isaksen, G.V.; Tiberti, M.; Papaleo, E. A comparative study of cold-and warm-adapted Endonucleases A using sequence analyses and molecular dynamics simulations. PLoS ONE 2017, 12, e0169586. [Google Scholar] [CrossRef]
- Duarte, A.W.F.; Barato, M.B.; Nobre, F.S.; Polezel, D.A.; de Oliveira, T.B.; dos Santos, J.A.; Rodrigues, A.; Sette, L.D. Production of cold-adapted enzymes by filamentous fungi from King George Island, Antarctica. Polar Biol. 2018, 41, 2511–2521. [Google Scholar] [CrossRef] [Green Version]
- Baharudin, I.; Zaki, N.; Bakar, F.A.; Mahadi, N.; NAJMUDIN, N.; Illias, R.; Murad, A. Comparison of RNA extraction methods for transcript analysis from the psychrophilic yeast, Glaciozyma antarctica. Malays. Appl. Biol. 2014, 43, 71–79. [Google Scholar]
- Fell, J.W.; Pitt, J.I. Taxonomy of the yeast genus Metschnikowia: A correction and a new variety. J. Bacteriol. 1969, 98, 853–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Uden, N.; Fell, J. Marine yeasts. Adv. Microbiol. Sea. 1968, 1, 167–201. [Google Scholar]
- Hoondee, P.; Wattanagonniyom, T.; Weeraphan, T.; Tanasupawat, S.; Savarajara, A. Occurrence of oleaginous yeast from mangrove forest in Thailand. World J. Microbiol. Biotechnol. 2019, 35, 1–17. [Google Scholar] [CrossRef]
- Gerday, C.; Aittaleb, M.; Arpigny, J.L.; Baise, E.; Chessa, J.-P.; Garsoux, G.; Petrescu, I.; Feller, G. Psychrophilic enzymes: A thermodynamic challenge. Biochim. Et Biophys. Acta 1997, 1342, 119–131. [Google Scholar] [CrossRef]
- Dugdale, R.C.; Wilkerson, F.P. Low specific nitrate uptake rate: A common feature of high-nutrient, low-chlorophyll marine ecosystems. Limnol. Oceanogr. 1991, 36, 1678–1688. [Google Scholar] [CrossRef]
- Plíhal, O.; Sklenář, J.; Hofbauerová, K.; Novák, P.; Man, P.; Pompach, P.; Kavan, D.; Ryšlavá, H.; Weignerová, L.; Charvátová-Pišvejcová, A. Large propeptides of fungal β-N-acetylhexosaminidases are novel enzyme regulators that must be intracellularly processed to control activity, dimerization, and secretion into the extracellular environment. Biochemistry 2007, 46, 2719–2734. [Google Scholar] [CrossRef]
- Reyes, F.; Calatayud, J.; Vazquez, C.; Martínez, M.J. β-N-Acetylglucosaminidase from Aspergillus nidulans which degrades chitin oligomers during autolysis. FEMS Microbiol. Lett. 1989, 65, 83–87. [Google Scholar] [CrossRef]
- Santiago, M.; Ramírez-Sarmiento, C.A.; Zamora, R.A.; Parra, L.P. Discovery, molecular mechanisms, and industrial applications of cold-active enzymes. Front. Microbiol. 2016, 7, 1408. [Google Scholar] [CrossRef]
- Ito, E.; Ikemoto, Y.; Yoshioka, T. Thermodynamic implications of high Q 10 of thermo-TRP channels in living cells. Biophysics 2015, 11, 33–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Code | Name | Standard |
|---|---|---|---|
| Carbohydrates | BGL | β-glucosidase | MUF |
| BXY | β-xylosidase | MUF | |
| NAG | N-acetyl-β-D-glucosaminidase | MUF | |
| Proteins, peptides | LAP | Leucine aminopeptidase | MCA |
| Phosphorus | ALP | Alkaline phosphatase | MUF |
| Sulfur | SUL | Sulfatase | MUF |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salazar Alekseyeva, K.; Herndl, G.J.; Baltar, F. Extracellular Enzymatic Activities of Oceanic Pelagic Fungal Strains and the Influence of Temperature. J. Fungi 2022, 8, 571. https://doi.org/10.3390/jof8060571
Salazar Alekseyeva K, Herndl GJ, Baltar F. Extracellular Enzymatic Activities of Oceanic Pelagic Fungal Strains and the Influence of Temperature. Journal of Fungi. 2022; 8(6):571. https://doi.org/10.3390/jof8060571
Chicago/Turabian StyleSalazar Alekseyeva, Katherine, Gerhard J. Herndl, and Federico Baltar. 2022. "Extracellular Enzymatic Activities of Oceanic Pelagic Fungal Strains and the Influence of Temperature" Journal of Fungi 8, no. 6: 571. https://doi.org/10.3390/jof8060571