
White Rot Fungi (Hymenochaetales) and Esca of Grapevine: Insights from Recent Microbiome Studies


Abstract
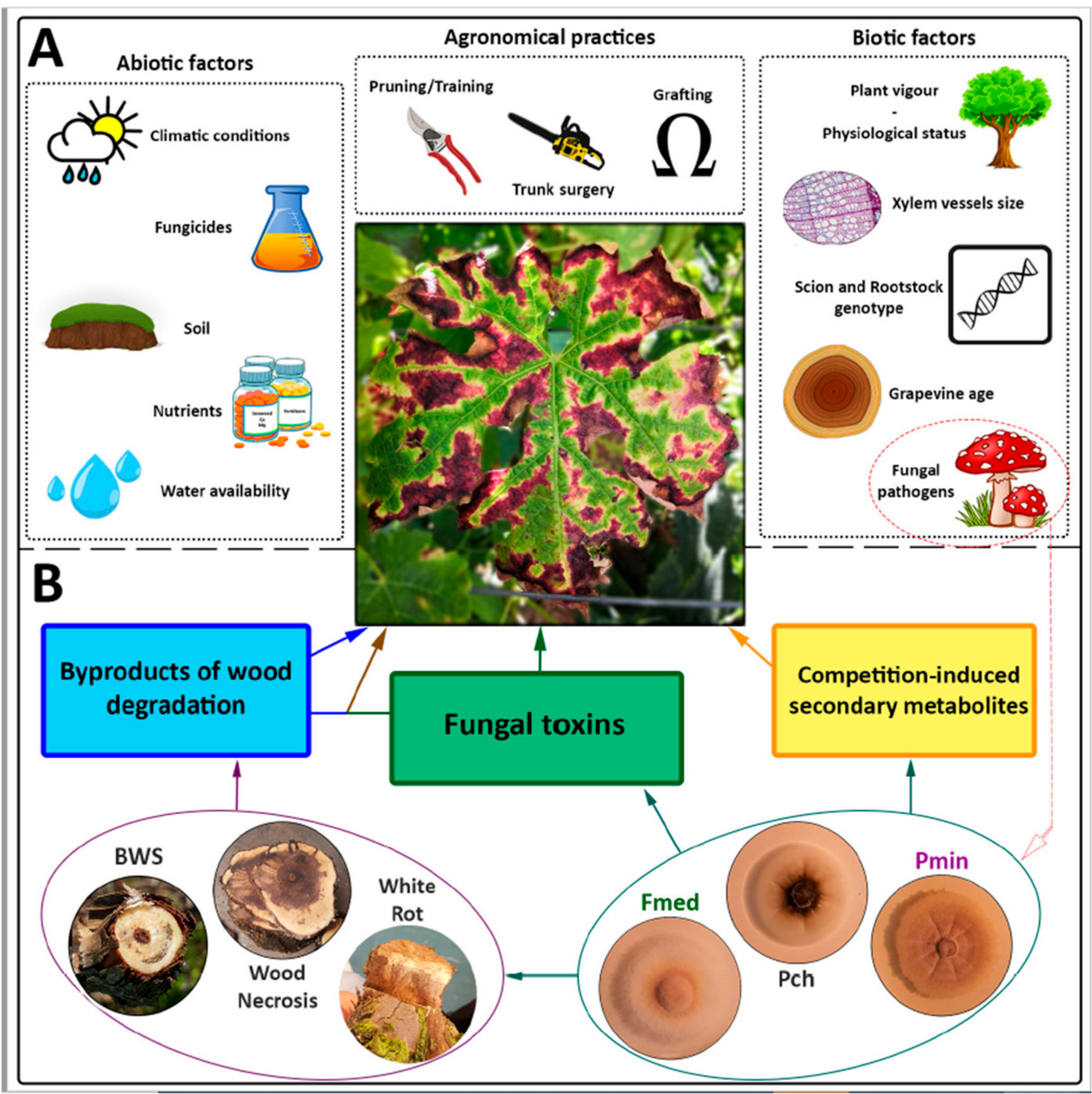
:1. Introduction
2. Natural Infections in the Field
2.1. Esca Proper
2.2. Grapevine Leaf Stripe Disease
3. Fulfilling Koch’s Postulates
3.1. Tracheomycotic Fungi
3.2. Hymenochaetales
4. Insights from Recent Microbiome Studies
5. What Did We Learn?
5.1. Toxins Hypothesis
5.2. Control of Leaf Symptoms
6. Conclusion and Perspectives
6.1. Esca Classification
6.2. Leaf Symptoms
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Valtaud, C.; Foyer, C.H.; Fleurat-Lessard, P.; Bourbouloux, A. Systemic effects on leaf glutathione metabolism and defence protein expression caused by esca infection in grapevines. Funct. Plant Biol. 2009, 36, 260–279. [Google Scholar] [CrossRef]
- Lima, M.R.M.; Felgueiras, M.L.; Graça, G.; Rodrigues, J.E.A.; Barros, A.; Gil, A.M.; Dias, A.C.P. NMR metabolomics of esca disease-affected Vitis vinifera cv. Alvarinho leaves. J. Exp. Bot. 2010, 61, 4033–4042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, M.R.M.; Felgueiras, M.L.; Cunha, A.; Chicau, G.; Ferreres, F.; Dias, A.C.P. Differential phenolic production in leaves of Vitis vinifera cv. Alvarinho affected with esca disease. Plant Physiol. Biochem. 2017, 112, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Valtaud, C.; Thibault, F.; Larignon, P.; Bertsch, C.; Fleurat, P.; Bourbouloux, A. Systemic damage in leaf metabolism caused by esca infection. Aust. J. Grape Wine Res. 2011, 17, 101–110. [Google Scholar] [CrossRef]
- Magnin-Robert, M.; Letousey, P.; Spagnolo, A.; Rabenoelina, F.; Jacquens, L.; Mercier, L.; Cĺement, C.; Fontaine, F. Leaf stripe form of esca induces alteration of photosynthesis and defence reactions in presymptomatic leaves. Funct. Plant Biol. 2011, 38, 856–866. [Google Scholar] [CrossRef]
- Lecomte, P.; Darrieutort, G.; Liminana, J.-M.; Comont, G.; Muruamendiaraz, A.; Legorburu, F.J.; Choueiri, E.; Jreijiri, F.; El Amil, R.; Fermaud, M. New insights into esca of grapevine: The development of foliar symptoms and their association with xylem discoloration. Plant Dis. 2012, 96, 924–934. [Google Scholar] [CrossRef] [Green Version]
- Calzarano, F.; D’Agostino, V.; Pepe, A.; Osti, F.; Della Pelle, F.; De Rosso, M.; Flamini, R.; Di Marco, S. Patterns of phytoalexins in the grapevine leaf stripe disease (esca complex)/grapevine pathosystem. Phytopathol. Mediterr. 2016, 55, 410–426. [Google Scholar] [CrossRef]
- Goufo, P.; Cortez, I. A lipidomic analysis of leaves of esca-affected grapevine suggests a role for galactolipids in the defense response and appearance of foliar symptoms. Biology 2020, 9, 268. [Google Scholar] [CrossRef]
- Surico, G. Towards a redefinition of the diseases within the esca complex of grapevine. Phytopathol. Mediterr. 2009, 48, 5–10. [Google Scholar] [CrossRef]
- Lecomte, P.; Darrieutort, G.; Laveau, C.; Blancard, D.; Louvet, G.; Goutouly, J.-P.; Rey, P.; Guérin-Dubrana, L. Impact of biotic and abiotic factors on the development of Esca decline disease. IOBC/WPRS Bull. 2011, 67, 171–180. [Google Scholar]
- Mary, S.; Laveau, C.; Lecomte, P.; Birebent, M.; Roby, J.-P. Impact of grafting type on Esca foliar symptoms. OENO One 2017, 51, 221–230. [Google Scholar] [CrossRef]
- Lecomte, P.; Diarra, B.; Carbonneau, A.; Rey, P.; Chevrier, C. Esca of grapevine and training practices in France: Results of a 10-year survey. Phytopathol. Mediterr. 2018, 57, 472–487. [Google Scholar] [CrossRef]
- Fischer, M.; Ashnaei, S.P. Grapevine, esca complex, and environment: The disease triangle. Phytopathol. Mediterr. 2019, 58, 17–37. [Google Scholar] [CrossRef]
- Calzarano, F.; Pagnani, G.; Pisante, M.; Bellocci, M.; Cillo, G.; Metruccio, E.G.; Di Marco, S. Factors involved on tiger-stripe foliar symptom expression of esca of grapevine. Plants 2021, 10, 1041. [Google Scholar] [CrossRef] [PubMed]
- Songy, A.; Vallet, J.; Gantet, M.; Boos, A.; Ronot, P.; Tarnus, C.; Clément, C.; Larignon, P.; Goddard, M.-L.; Fontaine, F. Sodium arsenite effect on Vitis vinifera L. physiology. J. Plant Physiol. 2019, 238, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Calzarano, F.; Di Marco, S. Further evidence that calcium, magnesium and seaweed mixtures reduce grapevine leaf stripe symptoms and increase grape yields. Phytopathol. Mediterr. 2018, 57, 459–471. [Google Scholar] [CrossRef]
- Calzarano, F.; Osti, F.; D’Agostino, V.; Pepe, A.; Di Marco, S. Mixture of calcium, magnesium and seaweed affects leaf phytoalexin contents and grape ripening on vines with grapevine leaf stripe disease. Phytopathol. Mediterr. 2017, 56, 445–457. [Google Scholar]
- Cholet, C.; Bruez, E.; Lecomte, P.; Barsacq, A.; Martignon, T.; Giudici, M. Plant resilience and physiological modifications induced by curettage of Esca-diseased grapevines. OENO One 2021, 1, 153–169. [Google Scholar] [CrossRef]
- Mugnai, L.; Graniti, A.; Surico, G. Esca (black measles) and brown wood-streaking: Two old and elusive diseases of grapevines. Plant Dis. 1999, 83, 404–418. [Google Scholar] [CrossRef] [Green Version]
- Claverie, M.; Notaro, M.; Fontaine, F.; Wery, J. Current knowledge on grapevine trunk diseases with complex etiology: A systemic approach. Phytopathol. Mediterr. 2020, 59, 29–53. [Google Scholar] [CrossRef]
- Fischer, M.; González García, V. An annotated checklist of European basidiomycetes related to white rot of grapevine (Vitis vinifera). Phytopathol. Mediterr. 2015, 54, 281–298. [Google Scholar] [CrossRef]
- Cloete, M.; Fischer, M.; Mostert, L.; Halleen, F. Hymenochaetales associated with esca-related wood rots on grapevine with a special emphasis on the status of esca in South African vineyards. Phytopathol. Mediterr. 2015, 54, 299–312. [Google Scholar] [CrossRef]
- Moretti, S.; Pacetti, A.; Pierron, R.; Kassemeyer, H.-H.; Fischer, M.; Péros, J.-P.; Perez-Gonzalez, G.; Bieler, E.; Schilling, M.; Di Marco, S.; et al. Fomitiporia mediterranea M. Fisch., the historical Esca agent: A comprehensive review on the main grapevine wood rot agent in Europe. Phytopathol. Mediterr. 2021, 60, 351–379. [Google Scholar] [CrossRef]
- Fontaine, F.; Gramaje, D.; Armengol, J.; Smart, R.; Nagy, Z.A.; Borgo, M.; Rego, C.; Corio-Costet, M.-F. Grapevine Trunk Diseases. A Review; OIV Publications: Paris, France, 2016; p. 25. Available online: http://www.oiv.int/public/medias/4650/trunk-diseases-oiv-2016.pdf (accessed on 15 August 2021).
- Edwards, J.; Pascoe, I.G. Occurrence of Phaeomoniella chlamydospora and Phaeoacremonium aleophilum associated with Petri disease and esca in Australian grapevines. Australas. Plant Pathol. 2004, 33, 273–279. [Google Scholar] [CrossRef]
- White, C.L.; Halleen, F.; Mostert, L. Symptoms and fungi associated with esca in South African vineyards. Phytopathol. Mediterr. 2011, 50, 236–246. [Google Scholar] [CrossRef]
- Gatica, M.; Dubos, B.; Larignon, P. The “hoja de malvón” grape disease in Argentina. Phytopathol. Mediterr. 2000, 39, 41–45. [Google Scholar]
- Mundy, D.C.; Manning, M.A. Ecology and management of grapevine trunk diseases in New Zealand: A review. N. Z. Plant Prot. 2010, 63, 160–166. [Google Scholar] [CrossRef]
- Abreo, E.; Martínez, S.; Bettucci, L.; Lupo, S. Phaeomoniella chlamydospora and Phaeoacremonium spp. in grapevines from Uruguay. Phytopathol. Mediterr. 2011, 50, 77–85. [Google Scholar]
- Díaz, G.A.; Latorre, B.A. Infection caused by Phaeomoniella chlamydospora associated with esca-like symptoms in grapevine in Chile. Plant Dis. 2014, 98, 351–360. [Google Scholar] [CrossRef] [Green Version]
- Pouzoulet, J.; Pivovaroff, A.L.; Santiago, S.L.; Rolshausen, P.E. Can vessel dimension explain tolerance toward fungal vascular wilt diseases in woody plants? Lessons from Dutch elm disease and esca disease in grapevine. Front. Plant Sci. 2014, 5, 253. [Google Scholar] [CrossRef]
- Bortolami, G.; Gambetta, G.A.; Delzon, S.; Lamarque, L.J.; Pouzoulet, J.; Badel, E.; Burlett, R.; Charrier, G.; Cochard, H.; Dayer, S.; et al. Exploring the hydraulic failure hypothesis of esca leaf symptom formation. Plant Physiol. 2019, 181, 1163–1174. [Google Scholar] [CrossRef] [Green Version]
- Sparapano, L.; Bruno, G.; Graniti, A. Effects on plants of metabolites produced in culture by Phaeoacremonium chlamydosporum, P. aleophilum and Fomitiporia punctata. Phytopathol. Mediterr. 2000, 39, 169–177. [Google Scholar] [CrossRef]
- Pancher, M.; Ceol, M.; Corneo, P.E.; Longa, C.M.O.; Yousaf, S.; Pertot, I.; Campisano, A. Fungal endophytic communities in grapevines (Vitis vinifera L.) respond to crop management. Appl. Environ. Microbiol. 2012, 78, 4308–4317. [Google Scholar] [CrossRef] [Green Version]
- Dissanayake, A.J.; Purahong, W.; Wubet, T.; Hyde, K.D.; Zhang, W.; Xu, H.; Zhang, G.; Fu, C.; Liu, M.; Xing, Q.; et al. Direct comparison of culture-dependent and culture-independent molecular approaches reveal the diversity of fungal endophytic communities in stems of grapevine (Vitis vinifera). Fungal Divers. 2018, 90, 85–107. [Google Scholar] [CrossRef]
- Deyett, E.; Rolshausen, P.E. Endophytic microbial assemblage in grapevine. FEMS Microbiol. Ecol. 2020, 96, fiaa053. [Google Scholar] [CrossRef] [PubMed]
- Deyett, E.; Rolshausen, P.E. Temporal dynamics of the sap microbiome of grapevine under high Pierce’s disease pressure. Front. Plant Sci. 2019, 10, 1246. [Google Scholar] [CrossRef] [PubMed]
- Calzarano, F.; Di Marco, S. Wood discoloration and decay in grapevines with esca proper and their relationship with foliar symptoms. Phytopathol. Mediterr. 2007, 46, 96–101. [Google Scholar] [CrossRef]
- Graniti, A.; Surico, G.; Mugnai, L. Esca of grapevine: A disease complex or a complex of diseases? Phytopathol. Mediterr. 2000, 39, 16–20. [Google Scholar] [CrossRef]
- Armengol, J.; Vicent, A.; Torné, L.; García-Figueres, F.; García-Jiménez, J. Fungi associated with esca and grapevine declines in Spain: A three-year survey. Phytopathol. Mediterr. 2001, 40, 325–329. [Google Scholar] [CrossRef]
- Elena, G.; Bruez, E.; Rey, P.; Luque, J. Microbiota of grapevine woody tissues with or without esca-foliar symptoms in northeast Spain. Phytopathol. Mediterr. 2018, 57, 425–438. [Google Scholar] [CrossRef]
- Serra, S.; Borgo, M.; Zanzotto, A. Investigation into the presence of fungi associated with esca of young vines. Phytopathol. Mediterr. 2000, 39, 21–25. [Google Scholar] [CrossRef]
- Pollastro, S.; Dongiovanni, C.; Abbatecola, A.; Faretra, F. Observations on the fungi associated with esca and on spatial distribution of esca-symptomatic plants in Apulian (Italy) vineyards. Phytopathol. Mediterr. 2000, 39, 206–210. [Google Scholar]
- Fischer, M.; Kassemeyer, H.-H. Fungi associated with esca disease of grapevine in Germany. Vitis 2003, 42, 109–116. [Google Scholar] [CrossRef]
- Larignon, P.; Dubos, B. Fungi associated with esca disease in grapevine. Eur. J. Plant Pathol. 1997, 103, 147–157. [Google Scholar] [CrossRef]
- Peros, J.-P.; Berger, G.; Jamaux-Despreaux, I. Symptoms, wood lesions and fungi associated with esca in organic vineyards in Languedoc-Roussillon (France). J. Phytopathol. 2008, 156, 297–303. [Google Scholar] [CrossRef]
- Kuntzmann, P.; Villaume, S.; Larignon, P.; Bertsch, C. Esca, BDA and eutypiosis: Foliar symptoms, trunk lesions and fungi observed in diseased vinestocks in two vineyards in Alsace. Vitis 2010, 49, 71–76. [Google Scholar] [CrossRef]
- Bruez, E.; Vallance, J.; Gerbore, J.; Lecomte, P.; Da Costa, J.P.; Guerin-Dubrana, L.; Rey, P. Analyses of the temporal dynamics of fungal communities colonizing the healthy wood tissues of esca leaf-symptomatic and asymptomatic vines. PLoS ONE 2014, 9, e95928. [Google Scholar] [CrossRef]
- Ouadi, L.; Bruez, E.; Bastien, S.; Yacoub, A.; Coppin, C.; Guérin-dubrana, L.; Fontaine, F.; Domec, J.-C.; Rey, P. Sap flow disruption in grapevine is the early signal predicting the structural, functional, and genetic responses to esca disease. Front. Plant Sci. 2021, 12, 1315. [Google Scholar] [CrossRef] [PubMed]
- Maher, N.; Piot, J.; Bastien, S.; Vallance, J.; Rey, P.; Guérin-dubrana, L. Wood necrosis in esca-affected vine: Types, relationships and possible links with foliar symptom expression. J. Int. des Sci. de la Vigne et du Vin. 2012, 46, 15–27. [Google Scholar] [CrossRef]
- Sofia, J.; Gonçalves, M.T.; Oliveira, H. Spatial distribution of esca symptomatic plants in Dão vineyards (Centre Portugal) and isolation of associated fungi. Phytopathol. Mediterr. 2006, 45, 87–92. [Google Scholar] [CrossRef]
- White, C.; Halleen, F.; Fischer, M.; Mostert, L. Characterisation of the fungi associated with esca diseased grapevines in South Africa. Phytopathol. Mediterr. 2011, 50, 204–223. [Google Scholar]
- Edwards, J.; Marchi, G.; Pascoe, I.G. Young esca in Australia. Phytopathol. Mediterr. 2001, 40, 303–310. [Google Scholar]
- Romanazzi, G.; Murolo, S.; Pizzichini, L.; Nardi, S. Esca in young and mature vineyards, and molecular diagnosis of the associated fungi. Eur. J. Plant Pathol. 2009, 125, 277–290. [Google Scholar] [CrossRef]
- Hofstetter, V.; Buyck, B.; Croll, D.; Viret, O.; Couloux, A.; Gindro, K. What if esca disease of grapevine were not a fungal disease? Fungal Divers 2012, 54, 51–67. [Google Scholar] [CrossRef] [Green Version]
- Raimondo, M.L.; Carlucci, A.; Ciccarone, C.; Sadallah, A.; Lops, F. Identification and pathogenicity of lignicolous fungi associated with grapevine trunk diseases in southern Italy. Phytopathol. Mediterr. 2019, 58, 639–662. [Google Scholar] [CrossRef]
- Stone, J.K.; Polishook, J.D.; White, J.F. Endophytic fungi. In Biodiversity of Fungi; Mueller, G.M., Bills, G.F., White, J.F., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 241–270. [Google Scholar]
- Mugnai, L.; Guiggiani, M.; Bazzo, F.; Marchi, G.; Surico, G. Il mal dell’esca e la carie bianca del legno: Agenti causali, sintomatologia e casi studio di esca giovane. In Il Mal Dell’esca Della Vite; Surico, G., Mugnai, L., Eds.; ARSIA Regione Toscana: Florence, Italy, 2010; pp. 65–88. [Google Scholar]
- Sparapano, L.; Bruno, G.; Graniti, A. Three-year observation of grapevines cross-inoculated with esca-associated fungi. Phytopathol. Mediterr. 2001, 40, 376–386. [Google Scholar]
- Feliciano, A.J.; Eskalen, A.; Gubler, W.D. Differential susceptibility of three grapevine cultivars to Phaeoacremonium aleophilum and Phaeomoniella chlamydospora. Phytopathol. Mediterr. 2004, 43, 66–69. [Google Scholar]
- Úrbez-Torres, J.R.; Haag, P.; Bowen, P.; O’Gorman, D.T. Grapevine trunk diseases in British Columbia: Incidence and characterization of the fungal pathogens associated with esca and Petri diseases of grapevine. Plant Dis. 2014, 98, 469–482. [Google Scholar] [CrossRef] [Green Version]
- Ye, Q.; Jia, J.; Manawasinghe, I.S.; Li, X.; Zhang, W.; Mugnai, L.; Wu, X.; Hyde, K.D.; Yan, J. Fomitiporia punicata and Phaeoacremonium minimum associated with esca complex of grapevine in China. Phytopathol. Res. 2021, 3, 11. [Google Scholar] [CrossRef]
- Halleen, F.; Mostert, L.; Crous, P.W. Pathogenicity testing of lesser-known vascular fungi of grapevines. Australas. Plant Pathol. 2007, 36, 277–285. [Google Scholar] [CrossRef]
- Laveau, C.; Letouze, A.; Louvet, G.W.; Bastien, S.; Guérin-Dubrana, L. Differential aggressiveness of fungi implicated in esca and associated diseases of grapevine in France. Phytopathol. Mediterr. 2009, 48, 32–46. [Google Scholar] [CrossRef]
- Luque, J.; Martos, S.; Aroca, A.; Raposo, R. Symptoms and fungi associated with declining mature grapevine plants in northeast Spain. J. Plant Pathol. 2009, 91, 381–390. [Google Scholar]
- Díaz, G.A.; Auger, J.; Besoain, X.; Bordeu, E.; Latorre, B.A. Prevalence and pathogenicity of fungi associated with grapevine trunk diseases in Chilean vineyards. Cienc. Investig. Agrar. 2013, 40, 327–339. [Google Scholar] [CrossRef]
- Akgül, D.S.; Güngӧr Savaş, N.; Teker, T.; Keykubat, B.; Mayorquin, J.S.; Eskalen, A. Fungal trunk pathogens of Sultana Seedless vineyards in Aegean region of Turkey. Phytopathol. Mediterr. 2015, 54, 380–393. [Google Scholar] [CrossRef]
- Brown, A.A.; Lawrence, D.P.; Baumgartner, K. Role of basidiomycete fungi in the grapevine trunk disease esca. Plant Pathol. 2020, 69, 205–220. [Google Scholar] [CrossRef]
- Amarloo, O.A.; Mohammadi, H.; Mahdian, S.A.; Tajick Ghanbary, M.A. Identification and pathogenicity of fungal species associated with grapevine trunk diseases in Khorasan-Razavi province, Iran. Mycol. Iran 2020, 7, 83–94. [Google Scholar] [CrossRef]
- Chiarappa, L. Phellinus igniarius: The cause of spongy wood decay of black measles (“esca”) disease of grapevines. Phytopathol. Mediterr. 1997, 36, 109–111. [Google Scholar]
- Sparapano, L.; Bruno, G.; Ciccarone, C.; Graniti, A. Infection of grapevines by some fungi associated with esca. I. Fomitiporia punctata as a wood-rot inducer. Phytopathol. Mediterr. 2000, 39, 46–52. [Google Scholar] [CrossRef]
- Cloete, M.; Mostert, L.; Fischer, M.; Halleen, F. Pathogenicity of South African Hymenochaetales taxa isolated from esca-infected grapevines. Phytopathol. Mediterr. 2015, 54, 368–379. [Google Scholar] [CrossRef]
- Markakis, E.A.; Ligoxigakis, E.K.; Roussos, P.A.; Sergentani, C.K.; Kavroulakis, N.; Roditakis, E.N.; Koubouris, G.C. Differential susceptibility responses of Greek olive cultivars to Fomitiporia mediterranea. Eur. J. Plant Pathol. 2019, 153, 1055–1066. [Google Scholar] [CrossRef]
- Di Marco, S.; Calzarano, F.; Osti, F.; Mazzullo, A. Pathogenicity of fungi associated with a decay of kiwifruit. Australas. Plant Pathol. 2004, 33, 337–342. [Google Scholar] [CrossRef]
- Gobbi, A.; Kyrkou, I.; Filippi, E.; Ellegaard-Jensen, L.; Hansen, L.H. Seasonal epiphytic microbial dynamics on grapevine leaves under biocontrol and copper fungicide treatments. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Del Frari, G.; Gobbi, A.; Aggerbeck, M.R.; Oliveira, H.; Hansen, L.H.; Ferreira, R.B. Characterization of the wood mycobiome of Vitis vinifera in a vineyard affected by esca. Spatial distribution of fungal communities and their putative relation with leaf symptoms. Front. Plant Sci. 2019, 10, 910. [Google Scholar] [CrossRef] [Green Version]
- Bruez, E.; Vallance, J.; Gautier, A.; Laval, V.; Compant, S.; Maurer, W.; Sessitsch, A.; Lebrun, M.-H.; Rey, P. Major changes in grapevine wood microbiota are associated with the onset of esca, a devastating trunk disease. Environ. Microbiol. 2020, 22, 5189–5206. [Google Scholar] [CrossRef]
- Bruez, E.; Larignon, P.; Bertsch, C.; Robert-Siegwald, G.; Lebrun, M.; Rey, P.; Fontaine, F. Impacts of sodium arsenite on wood microbiota of esca-diseased grapevines. J. Fungi 2021, 7, 498. [Google Scholar] [CrossRef] [PubMed]
- Pacetti, A.; Moretti, S.; Pinto, C.; Farine, S.; Bertsch, C.; Mugnai, L. Trunk surgery as a tool to reduce foliar symptoms in diseases of the esca complex and its influence on vine wood microbiota. J. Fungi 2021, 7, 521. [Google Scholar] [CrossRef]
- Gramaje, D.; Mostert, L.; Groenewald, J.Z.; Crous, P.W. Phaeoacremonium: From esca disease to phaeohyphomycosis. Fungal Biol. 2015, 119, 759–783. [Google Scholar] [CrossRef]
- Niem, J.M.; Billones-baaijens, R.; Stodart, B.; Savocchia, S. Diversity profiling of grapevine microbial endosphere and antagonistic potential of endophytic Pseudomonas against grapevine trunk diseases. Front. Microbiol. 2020, 11, 477. [Google Scholar] [CrossRef] [PubMed]
- Haidar, R.; Yacoub, A.; Vallance, J.; Compant, S.; Antonielli, L.; Saad, A.; Habenstein, B.; Kauffmann, B.; Grélard, A.; Loquet, A.; et al. Bacteria associated with wood tissues of Esca-diseased grapevines: Functional diversity and synergy with Fomitiporia mediterranea to degrade wood components. Environ. Microbiol. 2021. [Google Scholar] [CrossRef]
- Marchi, G.; Peduto, F.; Mugnai, L.; Di Marco, S.; Calzarano, F.; Surico, G. Some observations on the relationship of manifest and hidden esca to rainfall. Phytopathol. Mediterr. 2006, 45, S117–S126. [Google Scholar]
- Bruez, E.; Lecomte, P.; Grosman, J.; Doublet, B.; Bertsch, C.; Fontaine, F.; Ugaglia, A.; Teissedre, P.L.; Da Costa, J.P.; Guerin-Dubrana, L.; et al. Overview of grapevine trunk diseases in France in the 2000s. Phytopathol. Mediterr. 2013, 52, 262–275. [Google Scholar] [CrossRef]
- Andreini, L.; Cardelli, R.; Bartolini, S.; Scalabrelli, G.; Viti, R. Esca symptoms appearance in Vitis vinifera L.: Influence of climate, pedo-climatic conditions and rootstock/cultivar combination. Vitis 2014, 53, 33–38. [Google Scholar]
- Calzarano, F.; Osti, F.; Baránek, M.; Di Marco, S. Rainfall and temperature influence expression of foliar symptoms of grapevine leaf stripe disease (esca complex) in vineyards. Phytopathol. Mediterr. 2018, 57, 488–505. [Google Scholar] [CrossRef]
- Serra, S.; Ligios, V.; Schianchi, N.; Prota, V.A.; Scanu, B. Expression of grapevine leaf stripe disease foliar symptoms in four cultivars in relation to grapevine phenology and climatic conditions. Phytopathol. Mediterr. 2018, 57, 557–568. [Google Scholar] [CrossRef]
- Schmidt, R.; Cordovez, V.; de Boer, W.; Raaijmakers, J.; Garbeva, P. Volatile affairs in microbial interactions. ISME J. 2015, 9, 2329–2335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazazzara, V.; Avesani, S.; Robatscher, P.; Oberhuber, M.; Pertot, I.; Schuhmacher, R.; Perazzolli, M. Biogenic volatile organic compounds in the grapevine response to pathogens, beneficial microorganisms, resistance inducers and abiotic factors. J. Exp. Bot. 2021, erab367. [Google Scholar] [CrossRef] [PubMed]
- Vaz, A.T.; Del Frari, G.; Chagas, R.; Ferreira, A.; Oliveira, H.; Ferreira, R.B. Precise nondestructive location of defective woody tissue in grapevines affected by wood diseases. Phytopathol. Mediterr. 2020, 59, 441–451. [Google Scholar] [CrossRef]
- Reis, P.; Magnin-Robert, M.; Nascimento, T.; Spagnolo, A.; Abou-Mansour, E.; Fioretti, C.; Clement, C.; Rego, C.; Fontaine, F. Reproducing Botryosphaeria Dieback foliar symptoms in a simple model system. Plant Dis. 2016, 100, 1071–1079. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Fungal Isolates | Symptoms | Essay Length (Months) | Grapevine Cultivar | Potted Plants (P)/Field Study (F) Plant Age (Years Old) | Country/Reference | ||
|---|---|---|---|---|---|---|---|
| White Rot | Tiger Stripes (%) | Other Wood Symptoms # | |||||
| Fomitiporia mediterranea | n | n | y | 6 | Cabernet Sauvignon | p (1) | France [45] |
| 18 | |||||||
| F. mediterranea (10 isolates) | n * | n | y | 15 | Cabernet Sauvignon | p (1) | France [64] |
| F. mediterranea (3 Isolates) | n | n | y | 9 | Macabeo | p (1) | Spain [65] |
| n | Tempranillo | ||||||
| Inocutis sp. | n | n | y | 15 | Carménère | p (2) | Chile [66] |
| 2 | Cabernet Sauvignon | F (1) | |||||
| F. mediterranea | n | n | y | 4 | Sultana Seedless | p (1) | Turkey [67] |
| F. polymorpha, F. langloisii and Tropicoporus texanus | n | y + | y | 12 | Crimson Seedless | p (1) | United States [68] |
| F. mediterranea | n | n | y | 10 | Kolahdari | p (2) | Iran [69] |
| F. punicata | n | 25% | y | 3 | Cabernet Franc | p (2) | China [62] |
| F. mediterranea | y | n | y | 96 | Emperor | F (7) | United States [70] |
| F. mediterranea (3 isolates) | y | n | n | 24 | Sangiovese | F (13) | Italy [59] |
| y ** | y | 8 | Italia | F (6) | |||
| Matilde | F (9) | ||||||
| F. mediterranea | y | 11.1–22.2% ++ | y | 36 | Italia | F (5) | Italy [71] |
| n | Matilde | F (9) | |||||
| Inonotus setuloso-croceus, Fomitiporella sp., F. capensis, Phellinus sp., others | y | n | n | 24 | Shiraz | F (10) | South Africa [68] |
| Y º | Mourvédre | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Del Frari, G.; Oliveira, H.; Boavida Ferreira, R. White Rot Fungi (Hymenochaetales) and Esca of Grapevine: Insights from Recent Microbiome Studies. J. Fungi 2021, 7, 770. https://doi.org/10.3390/jof7090770
Del Frari G, Oliveira H, Boavida Ferreira R. White Rot Fungi (Hymenochaetales) and Esca of Grapevine: Insights from Recent Microbiome Studies. Journal of Fungi. 2021; 7(9):770. https://doi.org/10.3390/jof7090770
Chicago/Turabian StyleDel Frari, Giovanni, Helena Oliveira, and Ricardo Boavida Ferreira. 2021. "White Rot Fungi (Hymenochaetales) and Esca of Grapevine: Insights from Recent Microbiome Studies" Journal of Fungi 7, no. 9: 770. https://doi.org/10.3390/jof7090770