
Inactivation of Dermatophytes Causing Onychomycosis Using Non-Thermal Plasma as a Prerequisite for Therapy

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
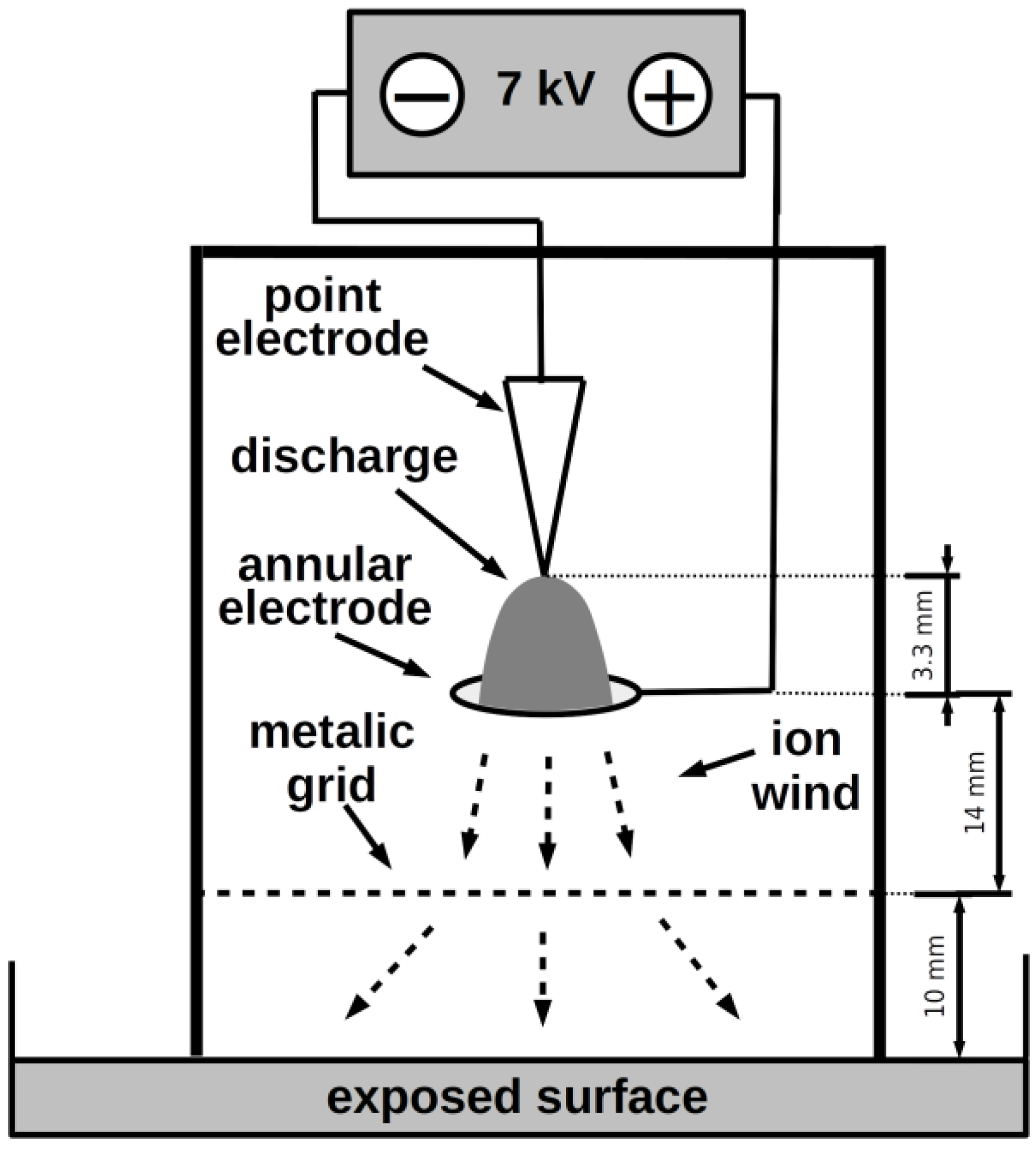
2.1. Plasma Source
2.2. Micromycete Strains
2.3. Exposure Arrangement
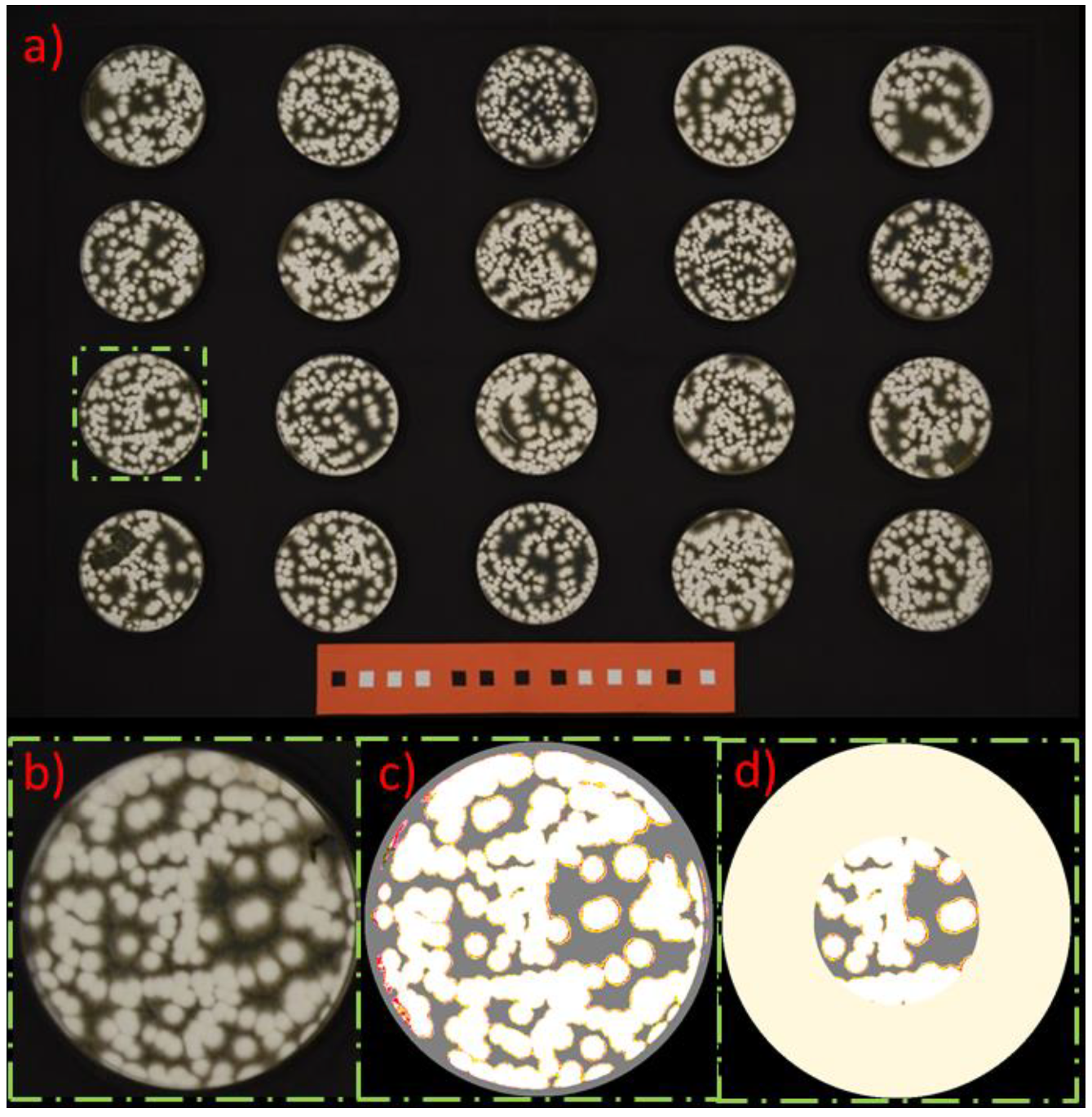
2.4. Evaluation
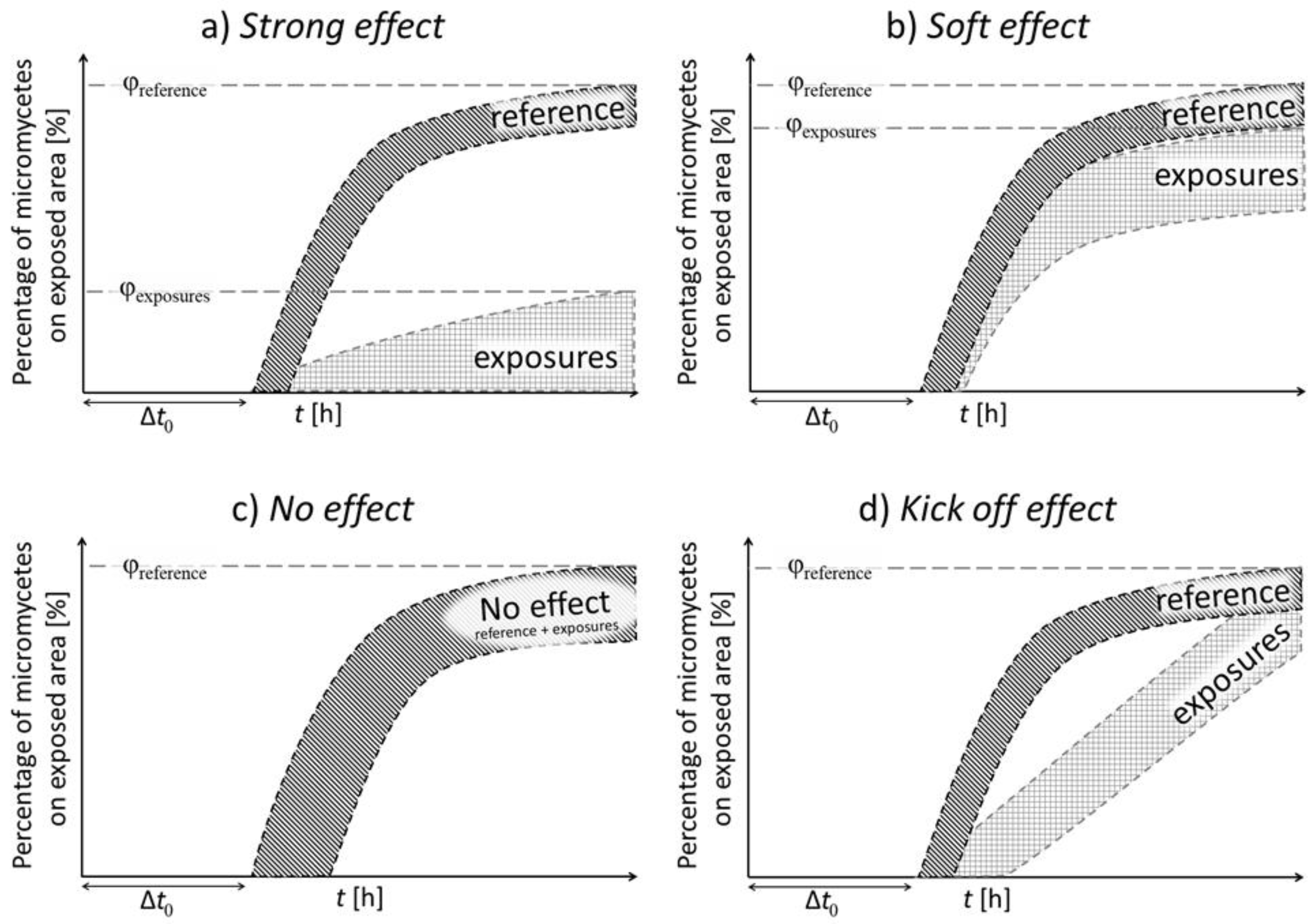
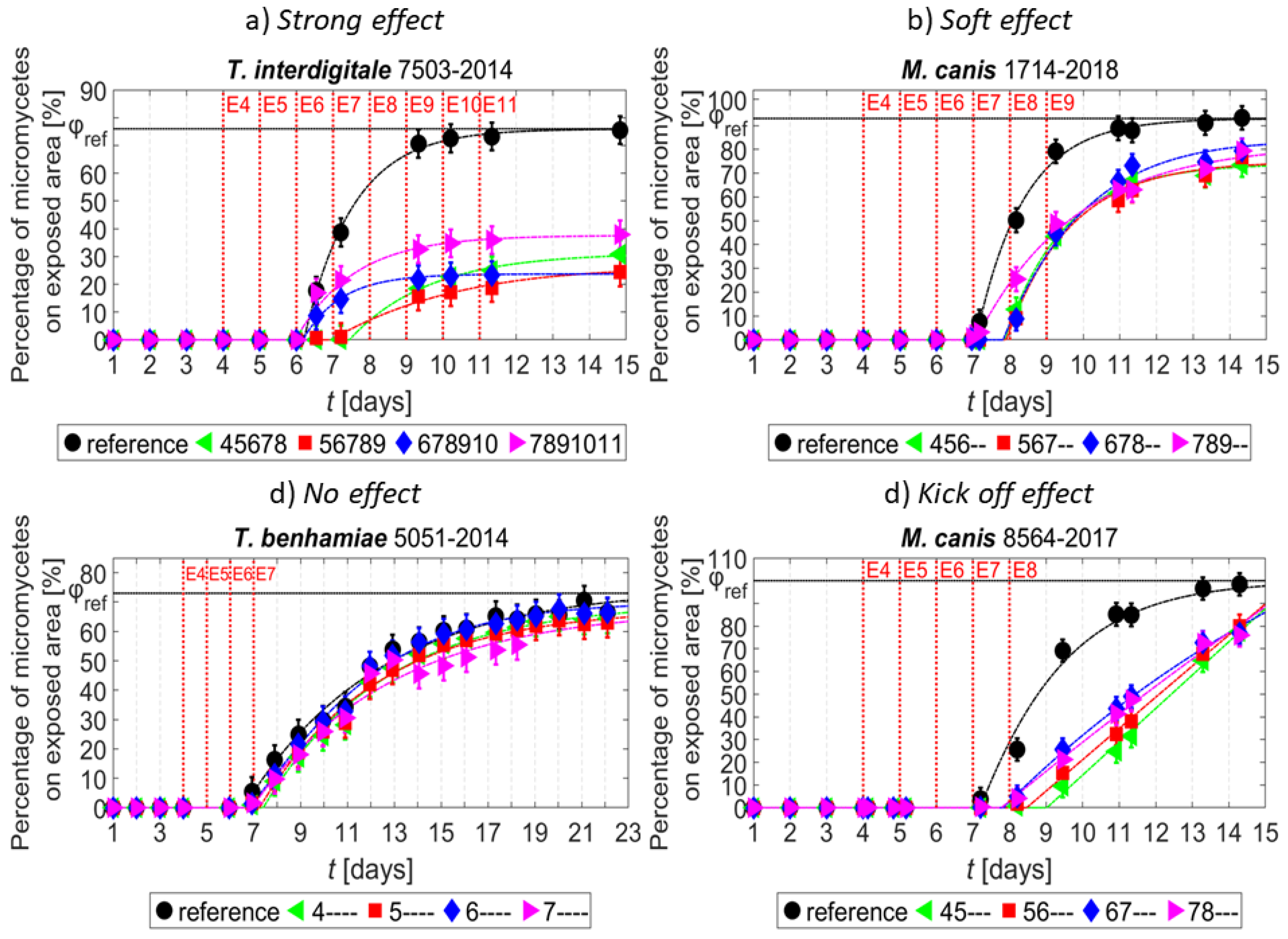
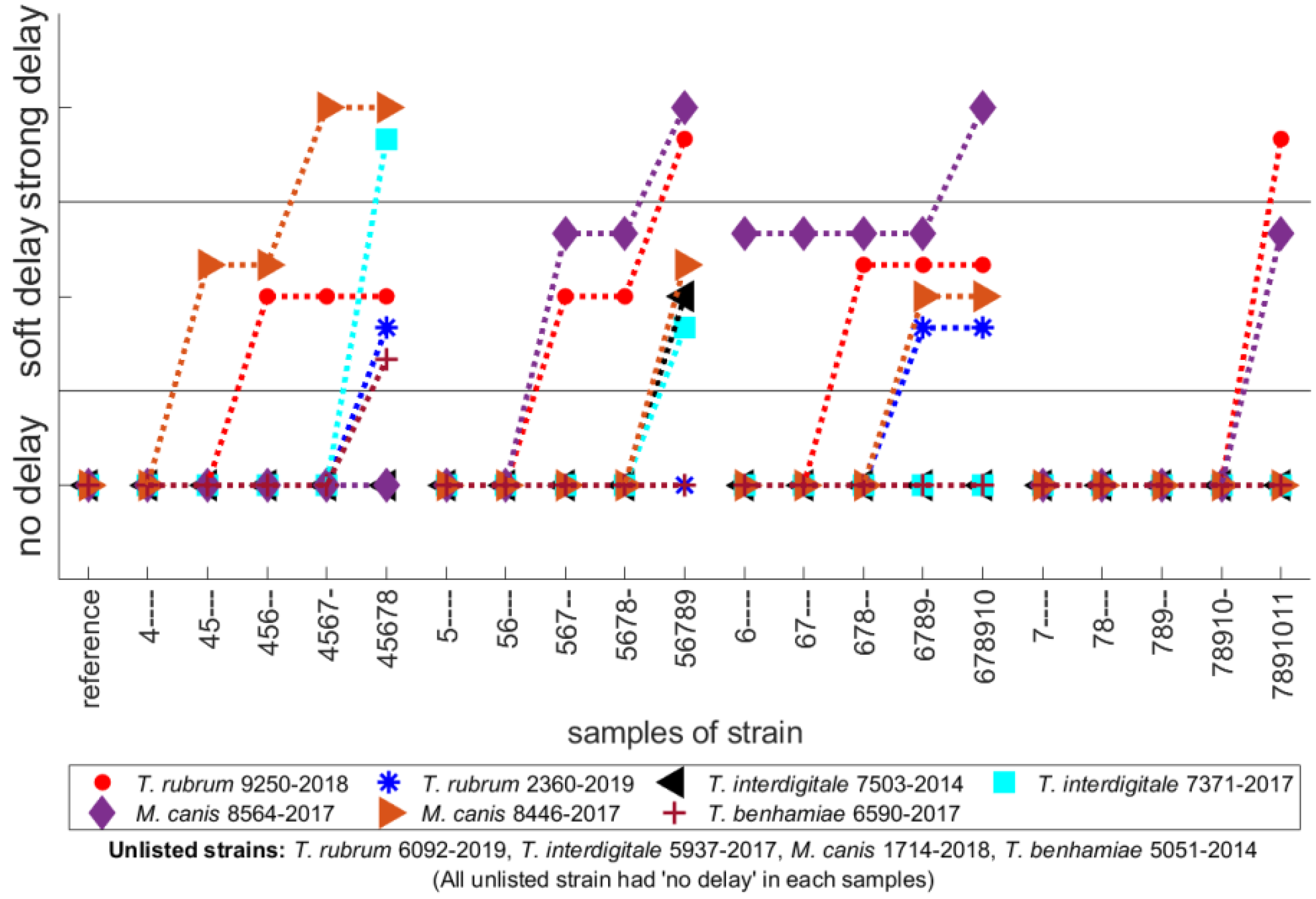
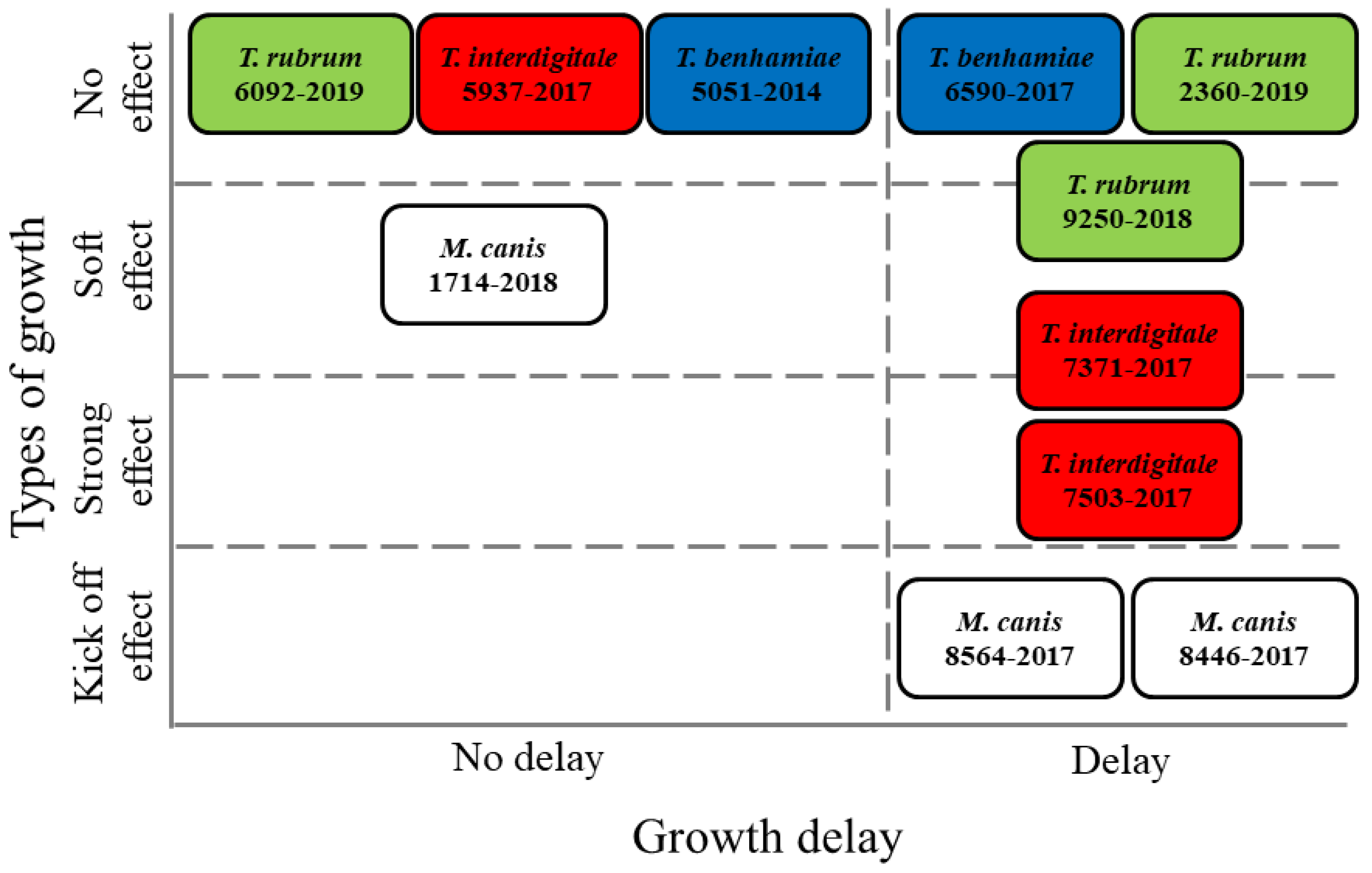
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yousfi, M.; Merbahi, N.; Sarrette, J.P.; Eichwald, O.; Ricard, A.; Gardou, J.-P.; Ducasse, O.; Benhenni, M. Non thermal plasma sources of production of active species for biomedical uses: Analyses, optimization and prospect. In Biomedical Engineering—Frontiers and Challenges; InTech: London, UK, 2011. [Google Scholar] [CrossRef] [Green Version]
- Khun, J.; Scholtz, V.; Hozák, P.; Fitl, P.E.; Julák, J. Various DC-driven point-to-plain discharges as non-thermal plasma sources and their bactericidal effects. Plasma Sources Sci. Technol. 2018, 27, 065002. [Google Scholar] [CrossRef]
- Setsuhara, Y. Low-temperature atmospheric-pressure plasma sources for plasma medicine. Arch. Biochem. Biophys. 2016, 605, 3–10. [Google Scholar] [CrossRef]
- Scholtz, V.; Julák, J. The “cometary” discharge, a possible new type of DC electric discharge in air at atmospheric pressure, and its bactericidal properties. J. Phys. Conf. Ser. 2010, 223, 012005. [Google Scholar] [CrossRef]
- Scholtz, V.; Julák, J. Plasma Jetlike point-to-point electrical discharge in air and its bactericidal properties. Plasma Sci. IEEE Trans. 2010, 38, 1978–1980. [Google Scholar] [CrossRef]
- Wong, K.S.; Lim, W.T.H.; Ooi, C.W.; Yeo, L.Y.; Tan, M.K. In situ generation of plasma-activated aerosols via surface acoustic wave nebulization for portable spray-based surface bacterial inactivation. Lab A Chip 2020, 20, 1856–1868. [Google Scholar] [CrossRef]
- Lux, J.; Dobiáš, R.; Kuklová, I.; Litvik, R.; Scholtz, V.; Soušková, H.; Khun, J.; Mrázek, J.; Kantorová, M.; Jaworská, P.; et al. Inactivation of dermatophytes causing onychomycosis and Its therapy using non-thermal plasma. J. Fungi 2020, 6, 214. [Google Scholar] [CrossRef]
- Moisan, M.; Barbeau, J.; Moreau, S.; Pelletier, J.; Tabrizian, M.; Yahia, L.H. Low-temperature sterilization using gas plasmas: A review of the experiments and an analysis of the inactivation mechanisms. Int. J. Pharm. 2001, 226, 1–21. [Google Scholar] [CrossRef]
- Graves, D.B. The emerging role of reactive oxygen and nitrogen species in redox biology and some implications for plasma applications to medicine and biology. J. Phys. D Appl. Phys. 2012, 45, 263001. [Google Scholar] [CrossRef]
- Kelly, S.; Turner, M.M. Atomic oxygen patterning from a biomedical needle-plasma source. J. Appl. Phys. 2013, 114, 123301. [Google Scholar] [CrossRef] [Green Version]
- Sysolyatina, E.; Mukhachev, A.; Yurova, M.; Grushin, M.; Karalnik, V.; Petryakov, A.; Trushkin, N.; Ermolaeva, S.; Akishev, Y. Role of the charged particles in bacteria inactivation by plasma of a positive and negative corona in ambient air. Plasma Process. Polym. 2014, 11, 315–334. [Google Scholar] [CrossRef]
- Liu, D.X.; Liu, Z.C.; Chen, C.; Yang, A.J.; Li, D.; Rong, M.Z.; Chen, H.L.; Kong, M.G. Aqueous reactive species induced by a surface air discharge: Heterogeneous mass transfer and liquid chemistry pathways. Sci. Rep. 2016, 6, 23737. [Google Scholar] [CrossRef]
- Cheng, J.-H.; Lv, X.; Pan, Y.; Sun, D.-W. Foodborne bacterial stress responses to exogenous reactive oxygen species (ROS) induced by cold plasma treatments. Trends Food Sci. Technol. 2020, 103, 239–247. [Google Scholar] [CrossRef]
- Lunov, O.; Zablotskii, V.; Churpita, O.; Jäger, A.; Polívka, L.; Syková, E.; Dejneka, A.; Kubinová, Š. The interplay between biological and physical scenarios of bacterial death induced by non-thermal plasma. Biomaterials 2016, 82, 71–83. [Google Scholar] [CrossRef]
- Paldrychová, M.; Vaňková, E.; Scholtz, V.; Julák, J.; Sembolová, E.; Mat’átková, O.; Masák, J. Effect of non-thermal plasma on AHL-dependent QS systems and biofilm formation in Pseudomonas aeruginosa: Difference between non-hospital and clinical isolates. AIP Adv. 2019, 9, 055117. [Google Scholar] [CrossRef] [Green Version]
- Čtvrtečková, L.; Pichová, A.; Scholtz, V.; Khun, J.; Julák, J. Non-thermal plasma-induced apoptosis in yeast Saccharomyces cerevisiae. Contrib. Plasma Phys. 2019, 59, e201800064. [Google Scholar] [CrossRef]
- Ito, M.; Hashizume, H.; Oh, J.-S.; Ishikawa, K.; Ohta, T.; Hori, M. Inactivation mechanism of fungal spores through oxygen radicals in atmospheric-pressure plasma. Jpn. J. Appl. Phys. 2020, 60, 010503. [Google Scholar] [CrossRef]
- Ali, A.; Hong, Y.J.; Park, J.; Lee, S.; Choi, E.H.; Kwon, G.C.; Park, B.J. A novel approach to inactivate the clinical isolates of Trichophyton mentagrophytes and Trichophyton rubrum by using non-thermal plasma. J. Microb. Biochem. Technol. 2014, 6, 314–319. [Google Scholar] [CrossRef] [Green Version]
- Borges, A.C.; Nishime, T.M.C.; de Moura Rovetta, S.; Lima, G.d.M.G.; Kostov, K.G.; Thim, G.P.; de Menezes, B.R.C.; Machado, J.P.B.; Koga-Ito, C.Y. Cold atmospheric pressure plasma jet reduces Trichophyton rubrum adherence and infection capacity. Mycopathologia 2019, 184, 585–595. [Google Scholar] [CrossRef]
- Bulson, J.M.; Liveris, D.; Derkatch, I.; Friedman, G.; Geliebter, J.; Park, S.; Singh, S.; Zemel, M.; Tiwari, R.K. Non-thermal atmospheric plasma treatment of onychomycosis in an in vitro human nail model. Mycoses 2020, 63, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Švarcová, M.; Julák, J.; Hubka, V.; Soušková, H.; Scholtz, V. Treatment of a superficial mycosis by low-temperature plasma: A case report. Prague Med Rep. 2014, 115, 73–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Julák, J.; Scholtz, V.; Vaňková, E. Medically important biofilms and non-thermal plasma. World J. Microbiol. Biotechnol. 2018, 34, 178. [Google Scholar] [CrossRef]
- Julák, J.; Soušková, H.; Scholtz, V.; Kvasničková, E.; Savická, D.; Kříha, V. Comparison of fungicidal properties of non-thermal plasma produced by corona discharge and dielectric barrier discharge. Folia Microbiol 2018, 63, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Soušková, H.; Scholtz, V.; Julák, J.; Kommová, L.; Savická, D.; Pazlarová, J. The survival of micromycetes and yeasts under the low-temperature plasma generated in electrical discharge. Folia Microbiol. 2011, 56, 77–79. [Google Scholar] [CrossRef]
- Scholtz, V.; Soušková, H.; Hubka, V.; Švarcová, M.; Julák, J. Inactivation of human pathogenic dermatophytes by non-thermal plasma. J. Microbiol. Methods 2015, 119, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Julák, J.; Scholtz, V.; Kvasničková, E.; Kříha, V.; Jíra, J. Bactericidal properties of cometary discharge with inserted grid. In Proceedings of the 4th International Conference on Plasma Medicine, Orléans, France, 17–21 June 2012; Mikikian, M., Rabat, H., Robert, E., Pouvesle, J.-M., Eds.; Book of Abstracts;. p. 141. [Google Scholar]
- Scholtz, V.; Kvasničková, E.; Julák, J. Microbial inactivation by electric discharge with metallic grid. Acta Phys. Pol. A 2013, 124, 62–65. [Google Scholar] [CrossRef]
- Hubka, V.; Vetrovsky, T.; Dobiášová, S.; Skořepová, M.; Lyskova, P.; Mencl, K.; Mallátová, N.; Janouškovcová, H.; Hanzlíčková, J.; Dobiáš, R.; et al. Molecular epidemiology of dermatophytoses in the Czech republic—Two-year-study results. Ceskoslovenská Dermatol. 2014, 89, 167–174. [Google Scholar]
- Hubka, V.; Čmoková, A.; Peano, A.; Větrovský, T.; Dobiáš, R.; Mallátová, N.; Lysková, P.; Mencl, K.; Janouškovcová, H.; Stará, J.; et al. Zoonotické dermatofytózy: Klinický obraz, diagnostika, etiologie, léčba, epidemiologická situace u nás. [Zoonose dermatophytoses: Clinics, diagnostic, etiology, treatment and epidemiologic situation in our country]. Cesk. Dermatol. 2018, 93, 205–292. [Google Scholar]
- Nise, N.S. Control Systems Engineering; John Wiley & Sons Inc.: Jefferson City, MO, USA, 2010; Volume 6, p. 688. [Google Scholar]
- Franklin, G.F.; Powell, J.D.; Emami-Naeini, A. Feedback Control of Dynamic Systems, 7th ed.; Pearson: New York, NY, USA, 2015; p. 885. [Google Scholar]
- Martins, M.; Silva, L.; Rossi, A.; Sanches, P.; Souza, L.; Martinez-Rossi, N. Global Analysis of Cell Wall Genes Revealed Putative Virulence Factors in the Dermatophyte Trichophyton rubrum. Front. Microbiol. 2019, 10, 2168. [Google Scholar] [CrossRef] [PubMed]
- Laurita, R.; Barbieri, D.; Gherardi, M.; Colombo, V.; Lukes, P. Chemical analysis of reactive species and antimicrobial activity of water treated by nanosecond pulsed DBD air plasma. Clin. Plasma Med. 2015, 3, 53–61. [Google Scholar] [CrossRef]
- Hozák, P.; Scholtz, V.; Khun, J.; Mertová, D.; Vaňková, E.; Julák, J. Further contribution to the chemistry of plasma-activated water: Influence on bacteria in planktonic and biofilm forms. Plasma Phys. Rep. 2018, 44, 799–804. [Google Scholar] [CrossRef]
- Thirumdas, R.; Kothakota, A.; Annapure, U.; Siliveru, K.; Blundell, R.; Gatt, R.; Valdramidis, V.P. Plasma activated water (PAW): Chemistry, physico-chemical properties, applications in food and agriculture. Trends Food Sci. Technol. 2018, 77, 21–31. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lokajová, E.; Julák, J.; Khun, J.; Soušková, H.; Dobiáš, R.; Lux, J.; Scholtz, V. Inactivation of Dermatophytes Causing Onychomycosis Using Non-Thermal Plasma as a Prerequisite for Therapy. J. Fungi 2021, 7, 715. https://doi.org/10.3390/jof7090715
Lokajová E, Julák J, Khun J, Soušková H, Dobiáš R, Lux J, Scholtz V. Inactivation of Dermatophytes Causing Onychomycosis Using Non-Thermal Plasma as a Prerequisite for Therapy. Journal of Fungi. 2021; 7(9):715. https://doi.org/10.3390/jof7090715
Chicago/Turabian StyleLokajová, Eliška, Jaroslav Julák, Josef Khun, Hana Soušková, Radim Dobiáš, Jaroslav Lux, and Vladimír Scholtz. 2021. "Inactivation of Dermatophytes Causing Onychomycosis Using Non-Thermal Plasma as a Prerequisite for Therapy" Journal of Fungi 7, no. 9: 715. https://doi.org/10.3390/jof7090715