
Basidiobolus omanensis sp. nov. Causing Angioinvasive Abdominal Basidiobolomycosis

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Phenotypic Studies
2.2. DNA Amplification and Sequencing
2.3. Phylogenetic Inference
2.4. Antifungal Susceptibility
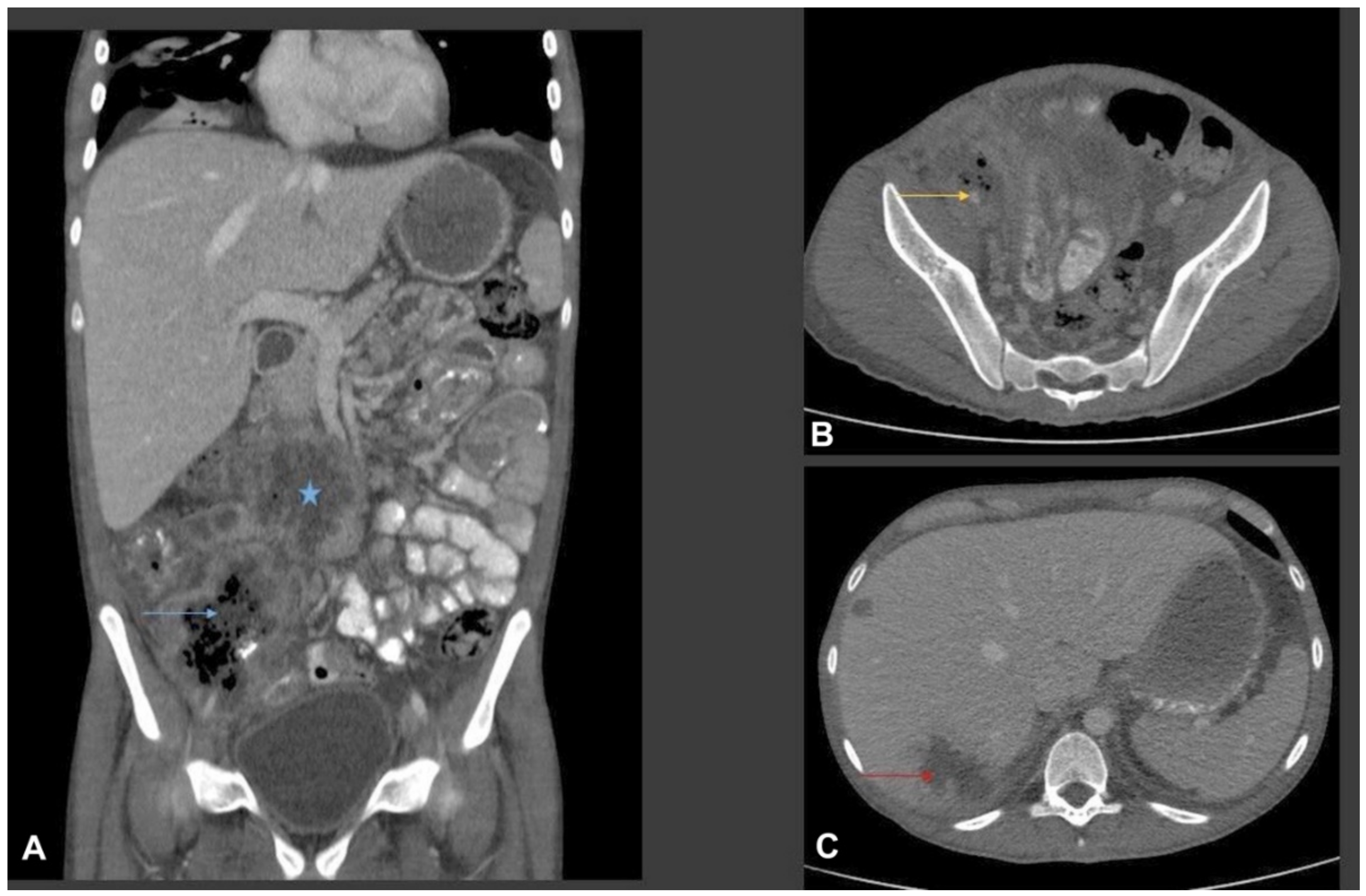
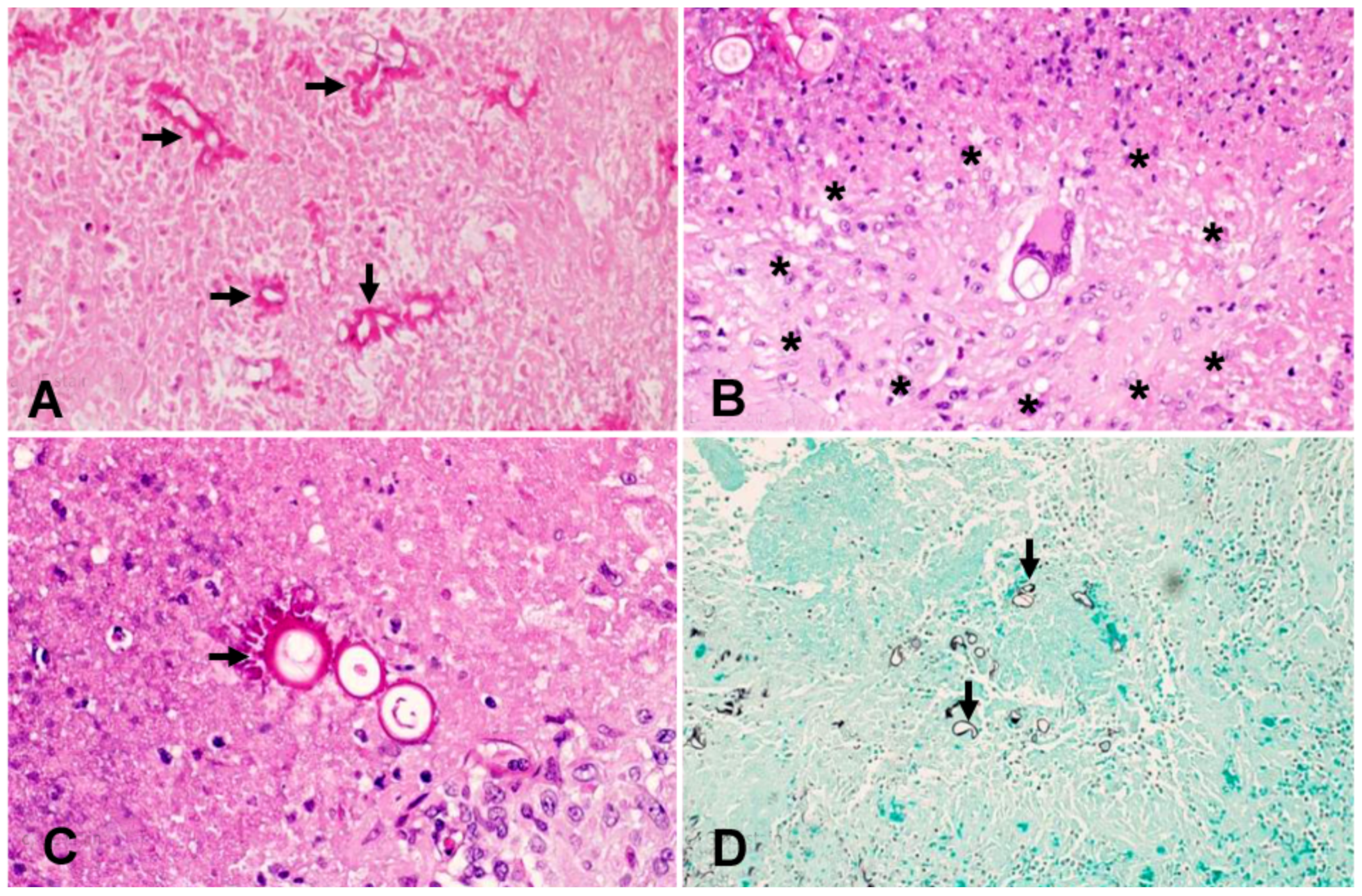
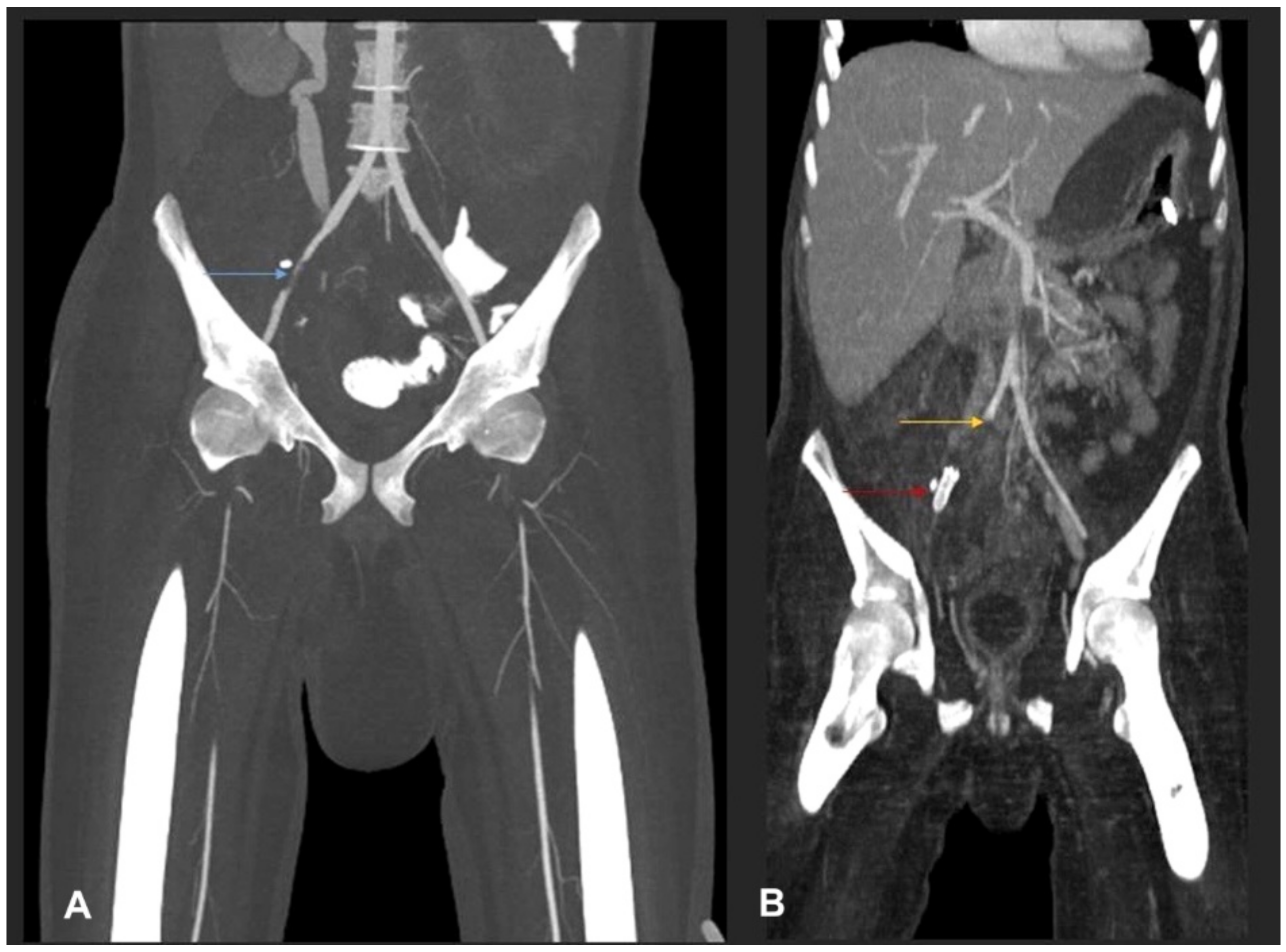
3. Case Report and Results
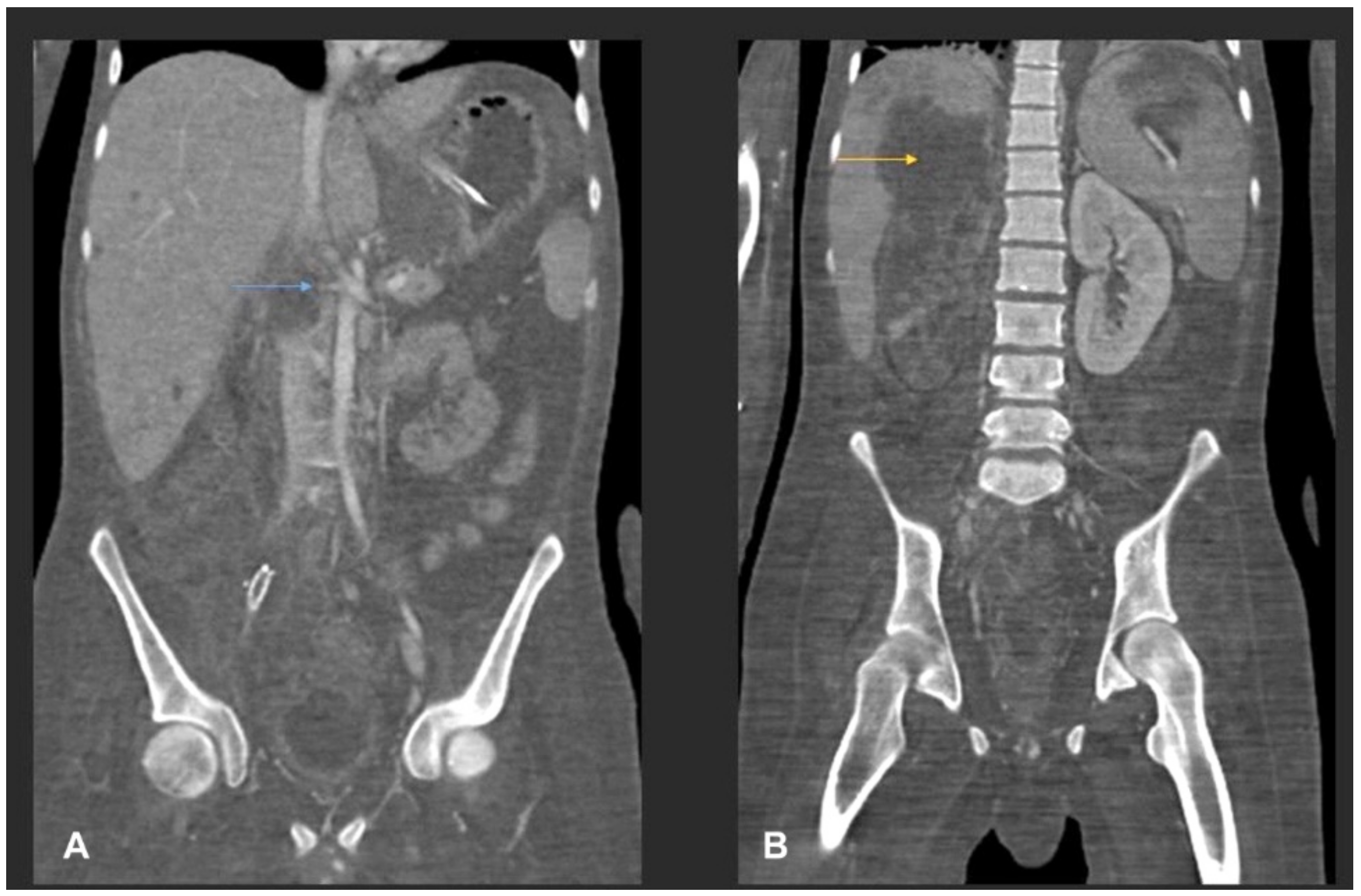
3.1. Case Presentation
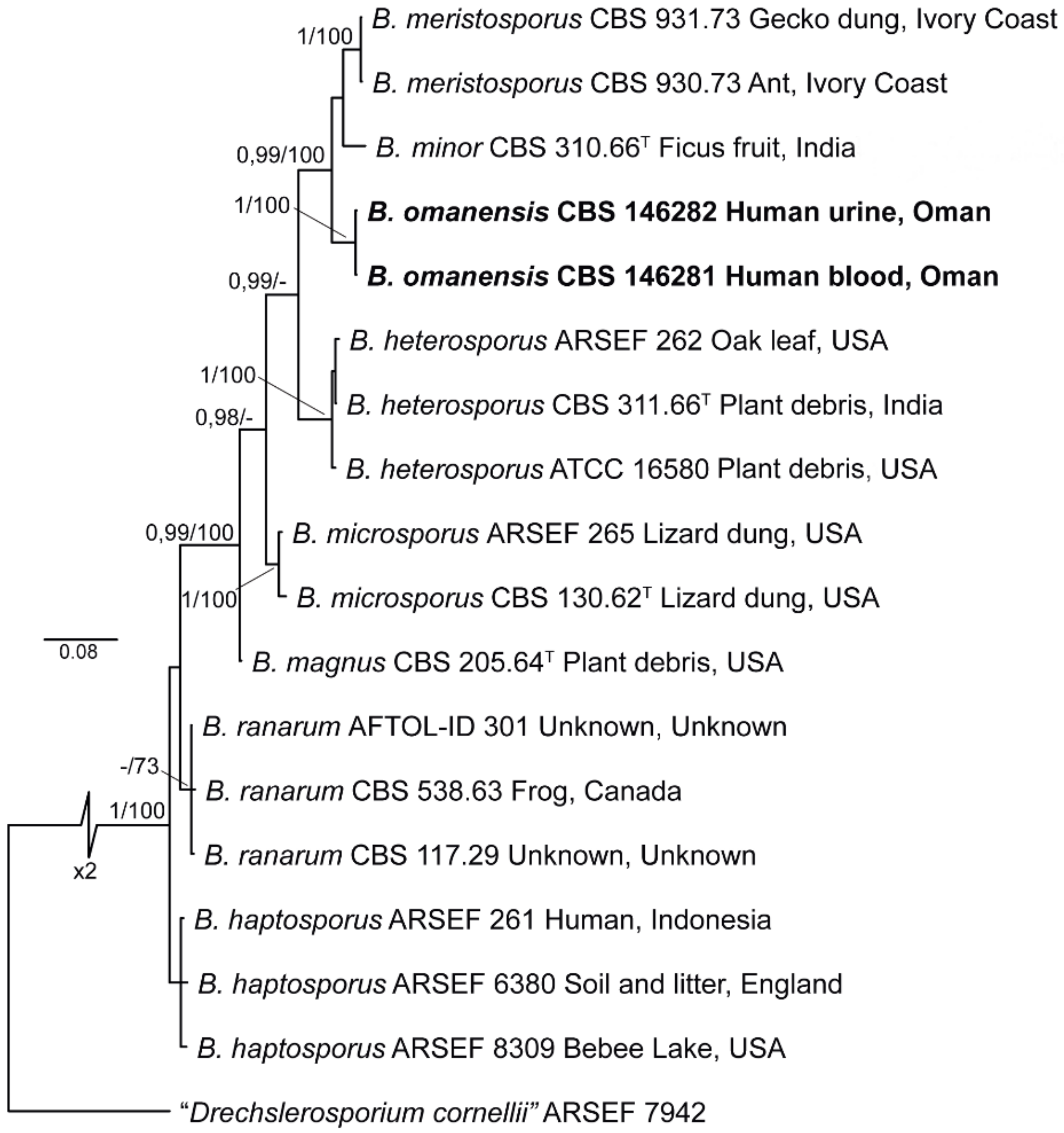
3.2. Phylogenetic Analysis
3.3. Antifungal Susceptibility
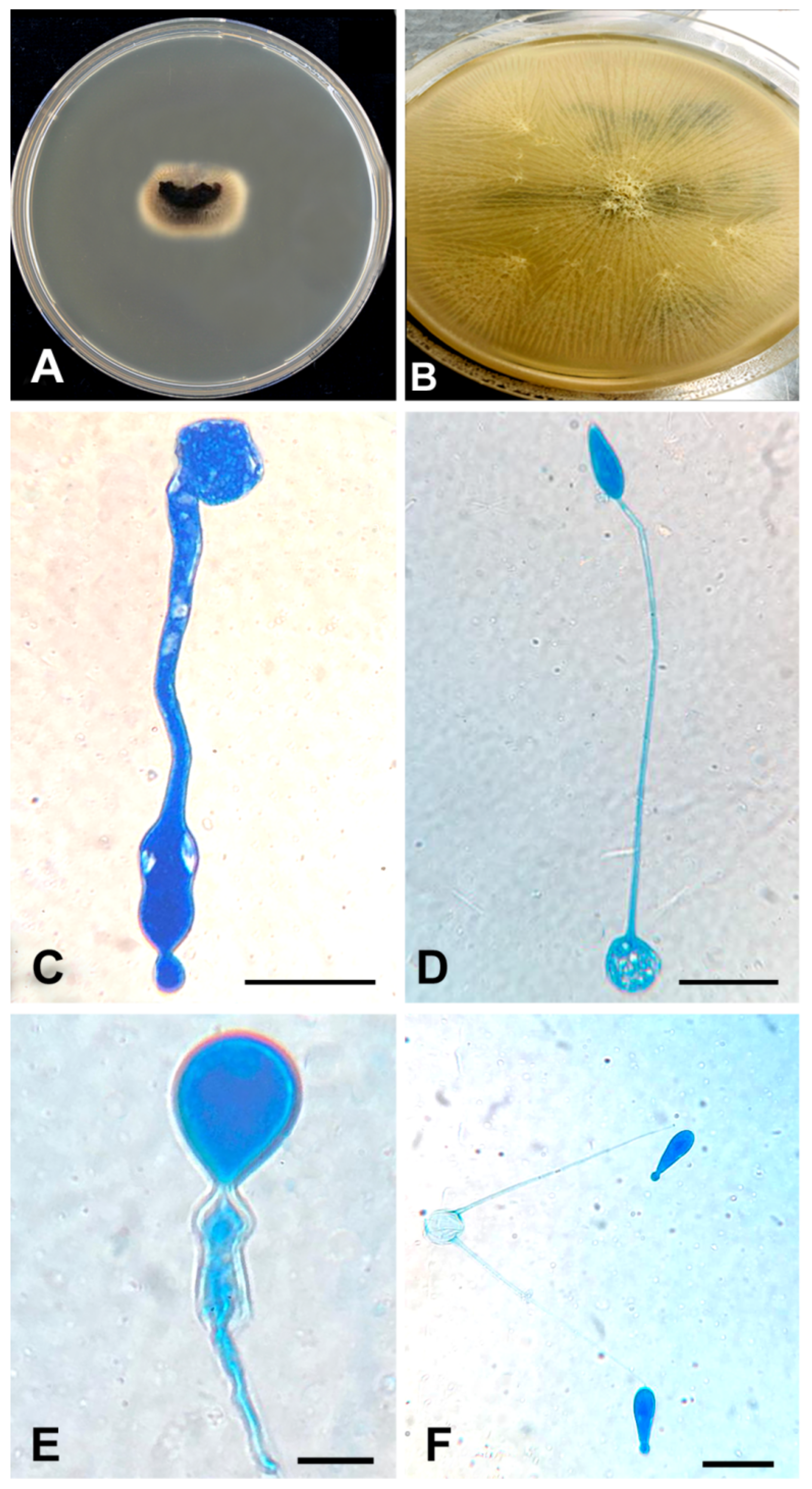
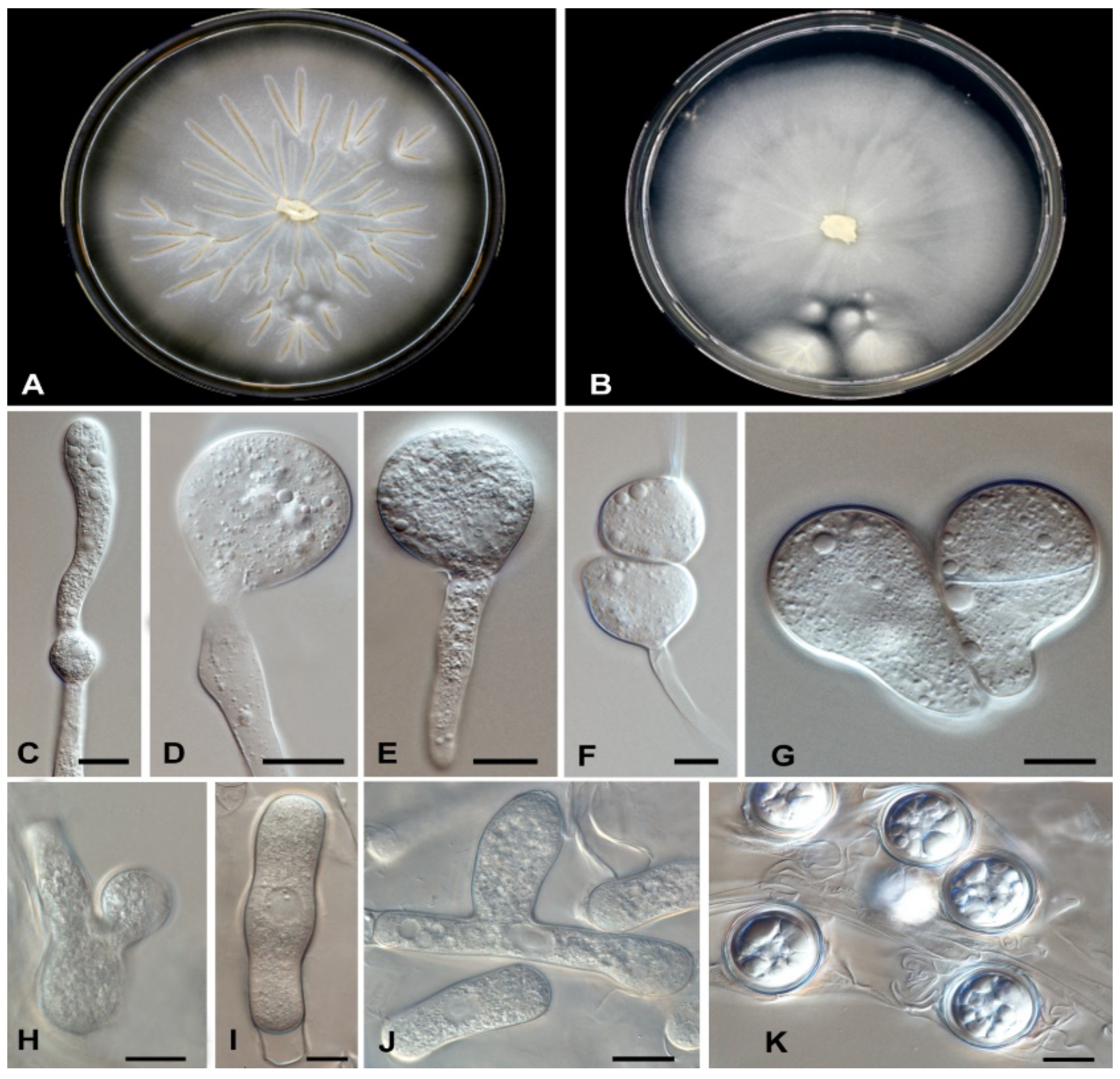
3.4. Taxonomy and Description
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Joe, L.K.; Njo-Injo, T.E.; Pohan, A.; Van Der Meulen, H. Basidiobolus ranarum as a cause of subcutaneous phycomycosis in Indonesia. Arch. Dermatol. Syphol. 1956, 74, 378–383. [Google Scholar] [CrossRef]
- Van Overeem, C. Uber ein merkwurdiges Vorkommen von Basidiobolus ranarum Eidam. Bull. Jardin Bot. 1925, 7, 423–431. [Google Scholar]
- Greene, C.E.; Brockus, C.W.; Currin, M.P.; Jones, C.J. Infection with Basidiobolus ranarum in two dogs. J. Am. Vet. Med. Assoc. 2002, 221, 528–532. [Google Scholar] [CrossRef]
- Al-Maani, A.S.; Paul, G.; Jardani, A.; Nayar, M.; Al-Lawati, F.; Al-Baluishi, S.; Hussain, I.B. Gastrointestinal basidiobolomycosis: First case report from Oman and literature review. Sultan Qaboos Univ. Med. J. 2014, 14, e241. [Google Scholar]
- Khan, Z.U.; Khoursheed, M.; Makar, R.; Al-Waheeb, S.; Al-Bader, I.; Al-Muzaini, A.; Chandy, R.; Mustafa, A.S. Basidiobolus ranarum as an etiologic agent of gastrointestinal zygomycosis. J. Clin. Microbiol. 2001, 39, 2360–2363. [Google Scholar] [CrossRef] [Green Version]
- AL-Naemi, A.Q.; Khan, L.A.; AL-Naemi, I.; Amin, K.; Athlawy, Y.A.; Awad, A.; Sun, Z. A Case report of gastrointestinal basidiobolomycosis treated with voriconazole. Medicine 2015, 94, e1430. [Google Scholar] [CrossRef]
- Al Bshabshe, A.; Joseph, M.R.P.; Al Hakami, A.M.; Al Azraqi, T.; Al Humayed, S.; Hamid, M.E. Basidiobolus haptosporus-like fungus as a causal agent of gastrointestinal basidiobolomycosis. Med. Mycol. 2020, 58, 264–267. [Google Scholar] [CrossRef] [PubMed]
- Schüβler, A.; Schwarzott, D.; Walker, C. A new fungal phylum, the Glomeromycota: Phylogeny and evolution. Mycol. Res. 2001, 105, 1413–1421. [Google Scholar] [CrossRef] [Green Version]
- Hibbett, D.S.; Binder, M.; Bischoff, J.F.; Blackwell, M.; Cannon, P.F.; Eriksson, O.E.; Huhndorf, S.; James, T.; Kirk, P.M.; Lücking, R.; et al. A higher-level phylogenetic classification of the Fungi. Mycol. Res. 2007, 111, 509–547. [Google Scholar] [CrossRef]
- Kwon-Chung, K.J. Taxonomy of fungi causing mucormycosis and entomophthoramycosis (zygomycosis) and nomenclature of the disease: Molecular mycologic perspectives. Clin. Infect. Dis. 2012, 54 (Suppl. S1), S8–S15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humber, R.A. Entomophthoromycota: A new phylum and reclassification for entomophthoroid fungi. Mycotaxon 2012, 120, 477–492. [Google Scholar] [CrossRef]
- Gryganskyi, A.P.; Humber, R.A.; Miadlikovska, J.; Smith, M.E.; Wu, S.; Voigt, K.; Walther, G.; Anishchenko, I.M.; Vilgalys, R. Molecular phylogeny of the Entomophthoromycota. Mol. Phylogenet. Evol. 2012, 65, 682–694. [Google Scholar] [CrossRef]
- Gryganskyi, A.P.; Humber, R.A.; Smith, M.E.; Hodge, K.; Huang, B.; Voigt, K.; Vilgalys, R. Phylogenetic lineages in Entomophthoromycota. Persoonia 2013, 30, 94–105. [Google Scholar] [CrossRef] [Green Version]
- Spatafora, J.W.; Chang, Y.; Benny, G.L.; Lazarus, K.; Smith, M.E.; Berbee, M.L.; Bonito, G.; Corradi, N.; Grigoriev, I.; Gryganskyi, A.; et al. A phylum-level phylogenetic classification of zygomycete fungi based on genome-scale data. Mycologia 2016, 108, 1028–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilela, R.; Mendoza, L. Human pathogenic Entomophthorales. Clin. Microbiol. Rev. 2018, 31, e00014-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eidam, E. Basidiobolus eine neue gattung der Entomophthoracean. Beitr. Biol. Pflanz. 1886, 4, 181–251. [Google Scholar]
- Claussen, M.; Schmidt, S. Differentiation of Basidiobolus spp. isolates: RFLP of a diagnostic PCR amplicon matches sequence-based classification and growth temperature preferences. J. Fungi 2021, 7, 110. [Google Scholar] [CrossRef] [PubMed]
- El-Shabrawi, M.H.F.; Arnaout, H.; Madkour, L.; Kamal, N.M. Entomophthoromycosis: A challenging emerging disease. Mycoses 2014, 57, 132–137. [Google Scholar] [CrossRef]
- Chetambath, R.; Deepa Sarma, M.S.; Suraj, K.P.; Jyothi, E.; Mohammed, S.; Philomina, B.J.; Ramadevi, S. Basidiobolus: An unusual cause of lung abscess. Lung India 2010, 27, 89–92. [Google Scholar] [CrossRef]
- Shaikh, N.; Hussain, K.A.; Petraitiene, R.; Schuetz, A.N.; Walsh, T.J. Entomophthoramycosis: A neglected tropical mycosis. Clin. Microbiol. Infect. 2016, 22, 688–694. [Google Scholar] [CrossRef] [Green Version]
- James, T.Y.; Kauff, F.; Schoch, C.L.; Matheny, P.B.; Hofstetter, V.; Cox, C.J.; Celio, G.; Gueidan, C.; Fraker, E.; Miadlikowska, J.; et al. Reconstructing the early evolution of fungi using a six-gene phylogeny. Nature 2006, 443, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Moller, E.M.; Bahnweg, G.; Sandermann, H.; Geiger, H.H. A simple and efficient protocol for isolation of high molecular weight DNA from filamentous fungi, fruit bodies, and infected plant tissues. Nucleic Acids Res. 1992, 20, 6115–6116. [Google Scholar] [CrossRef] [Green Version]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gargas, A.; Taylor, J.W. Polymerase chain reaction (PCR) primers for amplifying and sequencing 18S rDNA from lichenized fungi. Mycologia 1992, 84, 589–592. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNAgenes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software v. 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.A.; Pfeier, W.; Schwartz, T. The Cipres Science Gateway: Enabling high-impact science for phylogenetics researchers with limited resources. In Proceedings of the 1st Conference of the Extreme Science and Engineering Discovery Environment: Bridging from the Extreme to the Campus and Beyond, Chicago, IL, USA, 16–20 July 2012; pp. 1–8. [Google Scholar]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MrBayes: Bayesian inference of phylogeny. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Mason-Gamer, R.; Kellogg, E. Testing for phylogenetic conflict among molecular data sets in the tribe Triticeae (Gramineae). Syst. Biol. 1996, 45, 524–545. [Google Scholar] [CrossRef]
- Wiens, J.J. Testing phylogenetic methods with tree congruence: Phylogenetic analysis of polymorphic morphological characters in phrynosomatid lizards. Syst. Biol. 1998, 47, 427–444. [Google Scholar] [CrossRef] [Green Version]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi, 3rd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Rabie, M.E.; Al Qahtani, A.S.; Jamil, S.; Mikhail, N.T.; El Hakeem, I.; Hummadi, A.; Elshaar, K.E.; Abdelraheem, I.; Saudi, D. Gastrointestinal basidiobolomycosis: An emerging potentially lethal fungal infection. Saudi. Surg. J. 2019, 7, 1–9. [Google Scholar]
- Geramizadeh, B.; Heidari, M.; Shekarkhar, G. Gastrointestinal basidiobolomycosis, a rare and under-diagnosed fungal infection in immunocompetent hosts: A review article. Iran. J. Med. Sci. 2015, 40, 90–97. [Google Scholar] [PubMed]
- Vikram, H.R.; Smilack, J.D.; Leighton, J.A.; Crowell, M.D.; De Petris, G. Emergence of gastrointestinal basidiobolomycosis in the United States, with a review of worldwide cases. Clin. Infect. Dis. 2012, 54, 1685–1691. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, R.; Ansari Chaharsoghi, M.; Khorvash, F.; Kaleidari, B.; Sanei, M.H.; Ahangarkani, F.; Abtahian, Z.; Meis, J.F.; Badali, H. An unusual case of gastrointestinal basidiobolomycosis mimicking colon cancer; Literature and review. J. Mycol. Med. 2019, 29, 75–79. [Google Scholar] [CrossRef]
- El-Shabrawi, M.H.; Kamal, N.M.; Jouini, R.; Al-Harbi, A.; Voigt, K.; Al-Malki, T. Gastrointestinal basidiobolomycosis: An emerging fungal infection causing bowel perforation in a child. J. Med. Microbiol. 2011, 60, 1395–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gouba, N.; Drancourt, M. Digestive tract mycobiota: A source of infection. Med. Mal. Infect. 2015, 45, 9–16. [Google Scholar] [CrossRef]
- Flicek, K.T.; Vikram, H.R.; De Petris, G.D.; Johnson, C.D. Abdominal imaging findings in gastrointestinal basidiobolomycosis. Abdom. Imaging 2015, 40, 246–250. [Google Scholar] [CrossRef]
- Al Jarie, A.; Al Azraki, T.; Al Mohsen, I.; Al Jumaah, S.; Almutawa, A.; Mohd Fahim, Y.; Al Shehri, M.; Abu Dayah, A.; Ibrahim, A.; Maw Shabana, M. Basidiobolomycosis: Case series. J. Mycol. Med. 2011, 21, 37–45. [Google Scholar] [CrossRef]
- Al-Masqari, M.; Al-Maani, A.; Ramadhan, F. Gastrointestinal Basidiobolomycosis: An emerging fungal infection of the gastrointestinal tract, the Royal Hospital (Sultanate of Oman) experience. Pediatr. Inf. Dis. 2021, 3, 46–49. [Google Scholar]
- Balkhair, A.; Al Wahaibi, A.; Al-Qadhi, H.; Al-Harthy, A.; Lakhtakia, R.; Rasool, W.; Ibrahim, S. Gastrointestinal basidiobolomycosis: Beware of the great masquerade a case report. IDCases 2019, 18, e00614. [Google Scholar] [CrossRef]
- De Hoog, G.S.; Guarro, J.; Gené, J.; Ahmed, S.; Al-Hatmi, A.M.S.; Figueras, M.J.; Vitale, R.G. Atlas of Clinical Fungi, 4th ed.; Foundation Atlas of Clinical Fungi: Hilversum, The Netherlands, 2020. [Google Scholar]
- Drechsler, C. A southern Basidiobolus forming many sporangia from globose and from elongated adhesive conidia. J. Wash. Acad. Sci. 1955, 45, 49–56. [Google Scholar]
- Benjamin, R.K. A new Basidiobolus that forms microspores. Aliso 1962, 5, 223–233. [Google Scholar] [CrossRef] [Green Version]
- Drechsler, C. An odorous Basidiobolus often producing conidia plurally and forming some diclinous sexual apparatus. Am. J. Bot. 1964, 51, 770–777. [Google Scholar] [CrossRef]
- Hutchison, J.A.; King, D.S.; Nickerson, M.A. Studies on temperature requirements, odor production and zygospore wall undulation of the genus Basidiobolus. Mycologia 1972, 64, 467–474. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Epithet | Strain 1 | Host/Substrate | Country | Sequence Accession Number 2 | |
|---|---|---|---|---|---|
| ITS | LSU | ||||
| B. haptosporus | ARSEF 261 = NRRL 1232 = RSA 228 | Human | Indonesia | EF392520 | MH869969 |
| ARSEF 6380 | Soil and litter | UK | EF392528 | MH869969 | |
| ARSEF 8309 | Unknown | USA | EF392531 | EF392420 | |
| B. heterosporus | ARSEF 262 = RSA 964 | Oak leaf | USA | EF392521 | EF392411 |
| CBS 311.66 = ATCC 16580 = IMI 118288 = NRRL 3687 (ex-type) | Plant debris | India | MH858801 | JX242587 | |
| B. magnus | CBS 205.64 = ATCC 15379 = NRRL 3734 (ex-type) | Plant debris | USA | NR_077175 | EF392423 |
| B. meristosporus | CBS 931.73 | Gecko dung | Ivory Coast | MT830914 | MT831974 |
| CBS 930.73 | Ant cemetery of Paltothyreus tarsatus | Ivory Coast | MT830915 | MT831975 | |
| B. microsporus | ARSEF 265 = RSA 962 | Lizard dung | USA | EF392523 | DQ364202 |
| CBS 130.62 = ATCC 14708 = DSM 3120 = IMI 093345 = RSA 977 (ex-type) | Lizard dung | USA | NR_159603 | MH869698 | |
| B. minor | CBS 310.66 = ATCC 16579 = IMI 118287 = NRRL 3677 = NBRC 109014 (ex-type) | Ficus fruit | India | EF392535 | EF392424 |
| B. omanensis | CBS 146281 (ex-type) | human intravascular thrombus | Oman | MT830913 | MT831973 |
| CBS 146282 | Human urine | Oman | MT830912 | MT831972 | |
| B. ranarum | AFTOL-ID 301 | Unknown | Unknown | AY997030 | DQ273807 |
| CBS 538.63 | Frog | Canada | MH858348 | MH869969 | |
| CBS 117.29 = ATCC 11230 = IFO 9117 | Unknown | Unknown | IF00911701 a | IF00911701 a | |
| Conidiobolus sp. | ARSEF 7942 | Leaf litter | USA | EF392537 | KC788411 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Hatmi, A.M.S.; Balkhair, A.; Al-Busaidi, I.; Sandoval-Denis, M.; Al-Housni, S.; Ba Taher, H.; Al Shehhi, A.H.; Raniga, S.; Al Shaibi, M.; Al Siyabi, T.; et al. Basidiobolus omanensis sp. nov. Causing Angioinvasive Abdominal Basidiobolomycosis. J. Fungi 2021, 7, 653. https://doi.org/10.3390/jof7080653
Al-Hatmi AMS, Balkhair A, Al-Busaidi I, Sandoval-Denis M, Al-Housni S, Ba Taher H, Al Shehhi AH, Raniga S, Al Shaibi M, Al Siyabi T, et al. Basidiobolus omanensis sp. nov. Causing Angioinvasive Abdominal Basidiobolomycosis. Journal of Fungi. 2021; 7(8):653. https://doi.org/10.3390/jof7080653
Chicago/Turabian StyleAl-Hatmi, Abdullah M. S., Abdullah Balkhair, Ibrahim Al-Busaidi, Marcelo Sandoval-Denis, Saif Al-Housni, Hashim Ba Taher, Asmaa Hamdan Al Shehhi, Sameer Raniga, Maha Al Shaibi, Turkiya Al Siyabi, and et al. 2021. "Basidiobolus omanensis sp. nov. Causing Angioinvasive Abdominal Basidiobolomycosis" Journal of Fungi 7, no. 8: 653. https://doi.org/10.3390/jof7080653