
Timing of Resource Addition Affects the Migration Behavior of Wood Decomposer Fungal Mycelia


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Fungal Culture and Wood Block Preparation
2.2. Microcosm Preparation
2.3. Microcosm Incubation
2.4. Image Analysis
2.5. Statistical Analysis
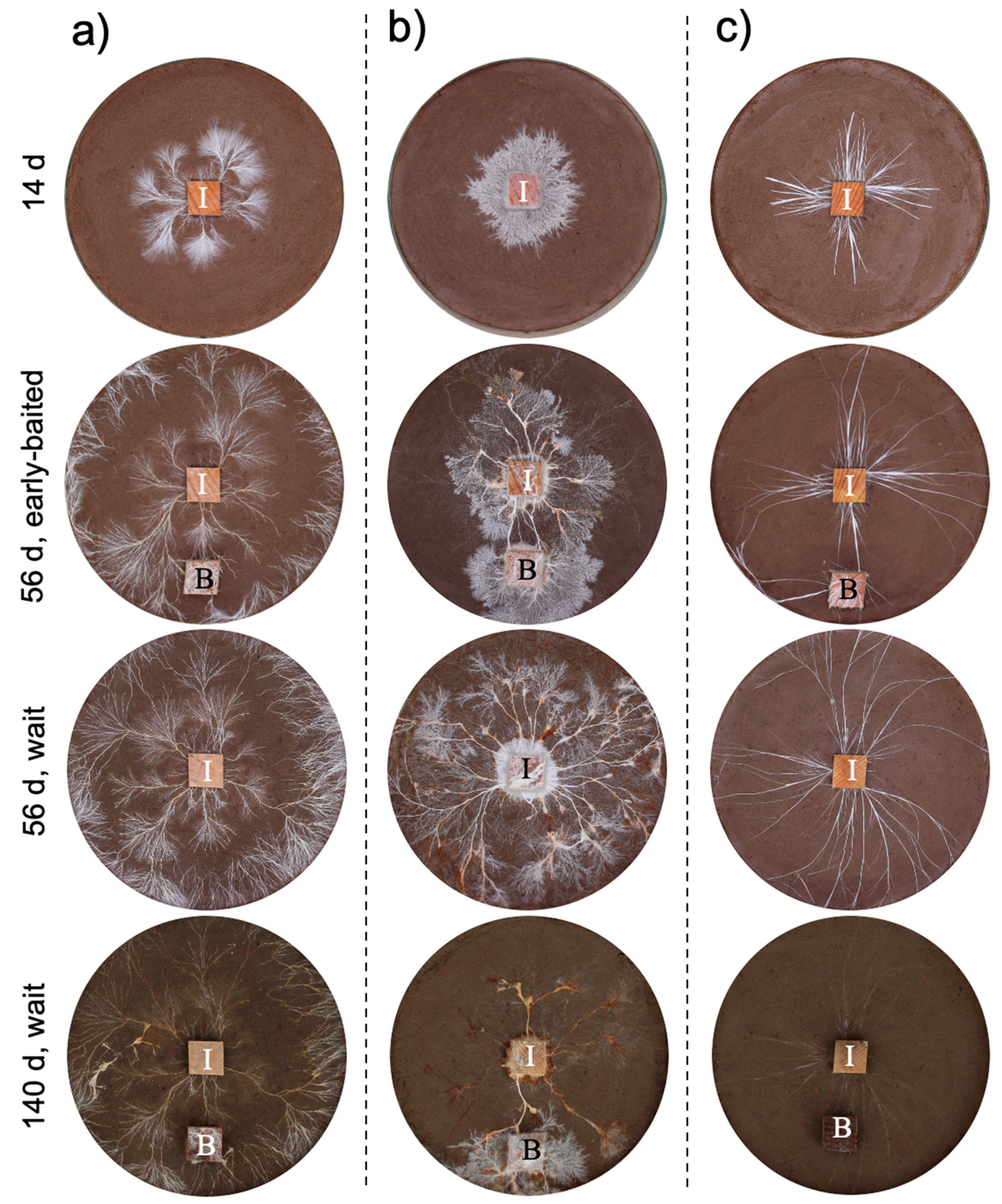
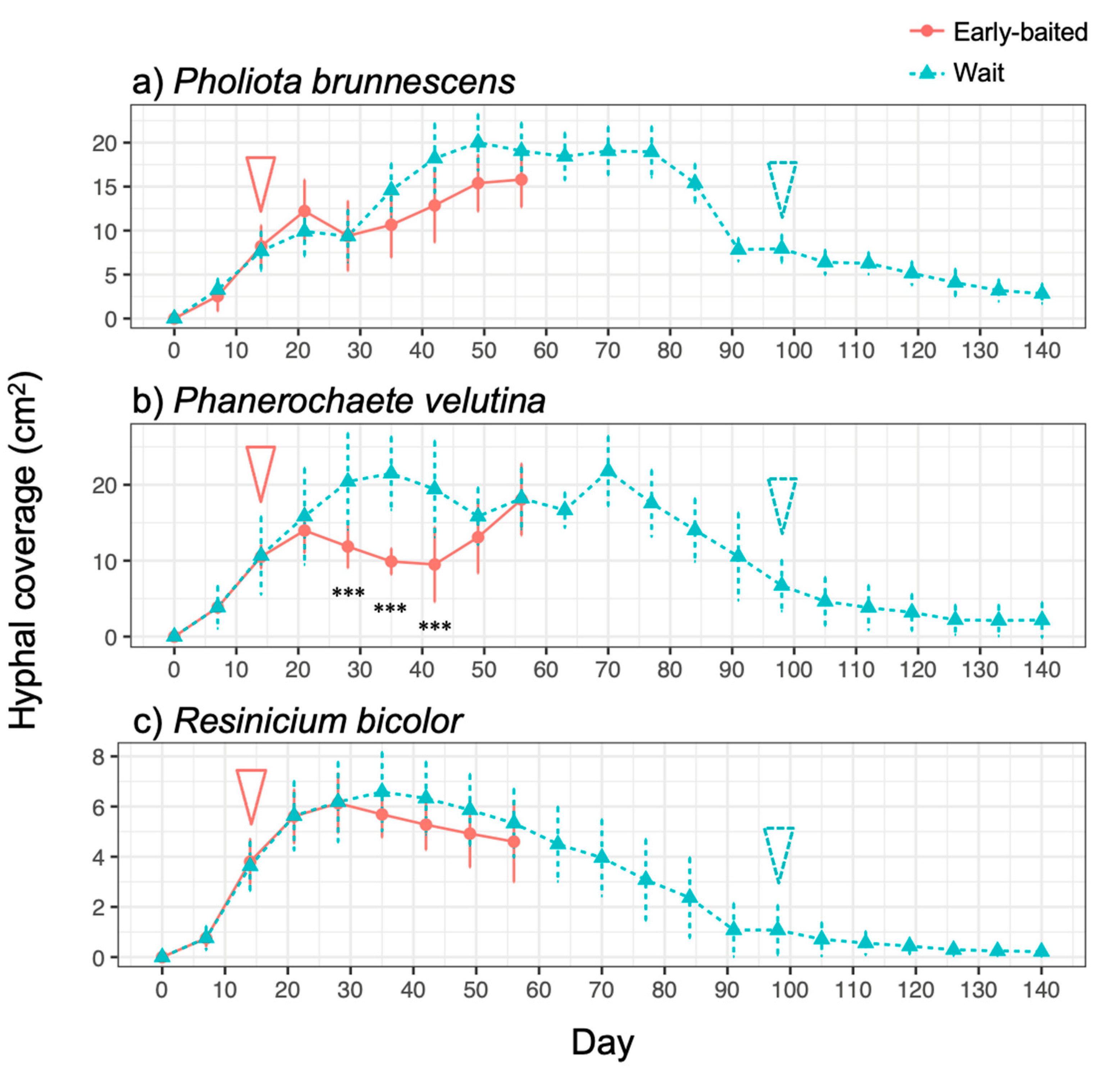
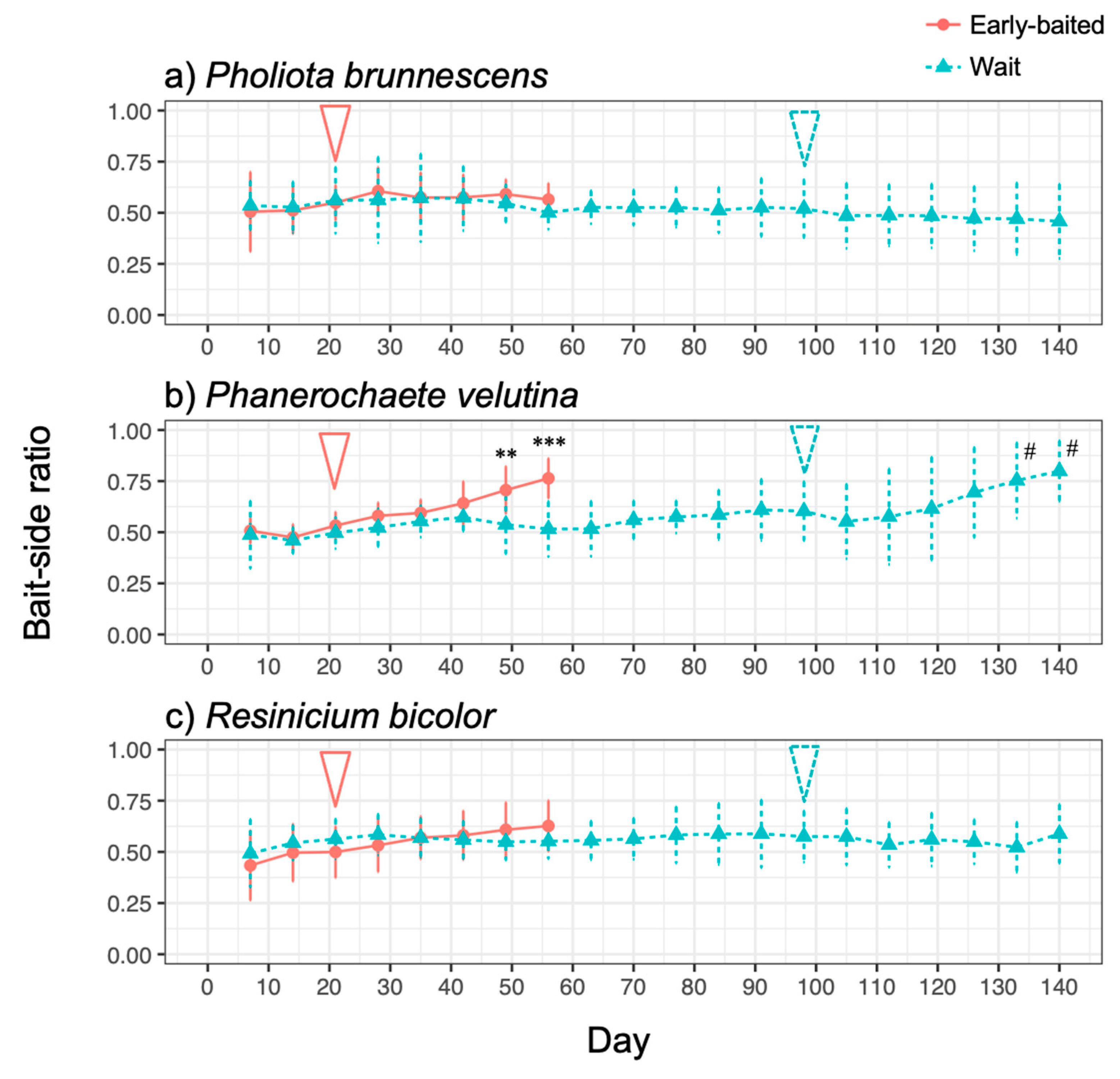
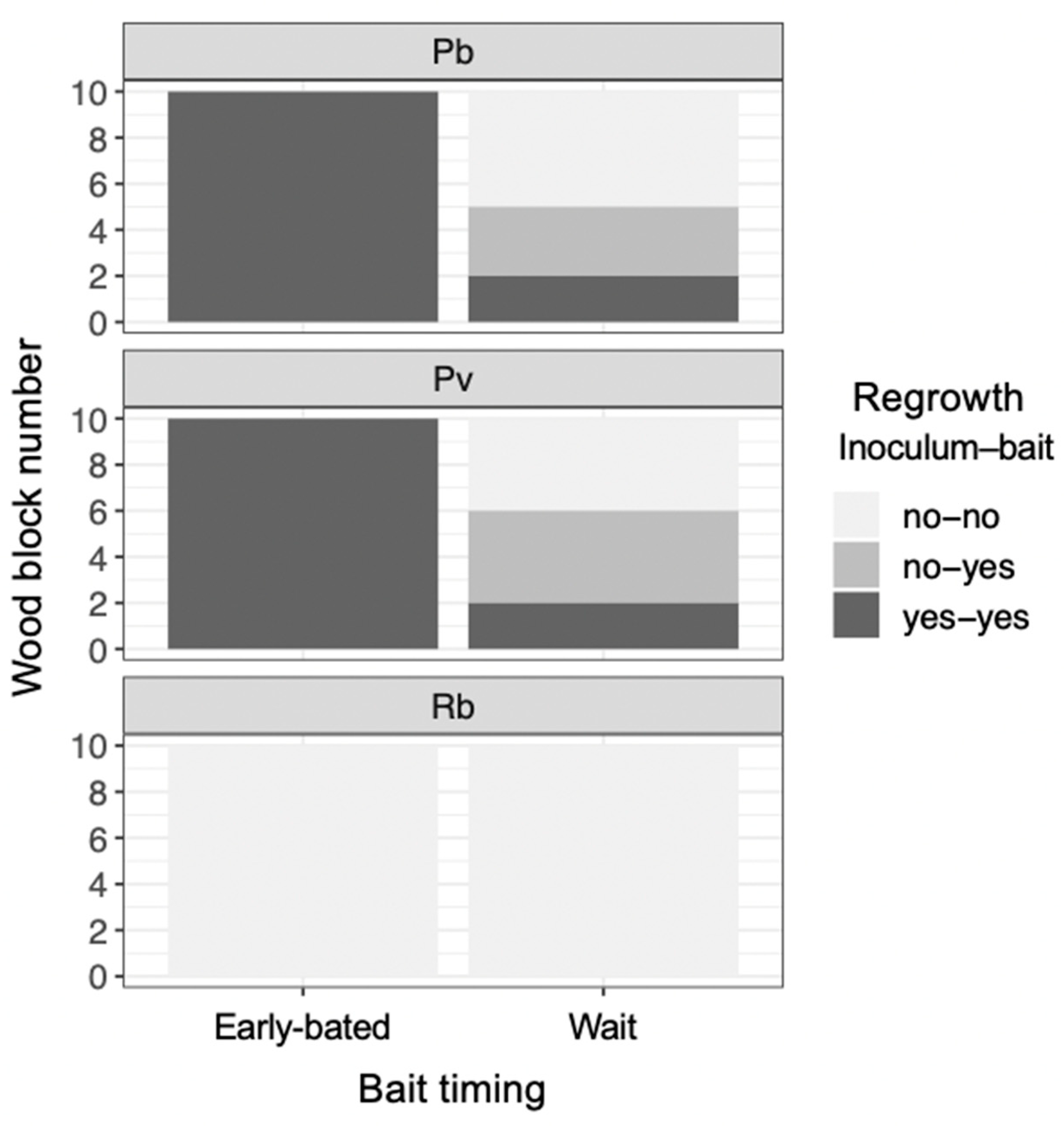
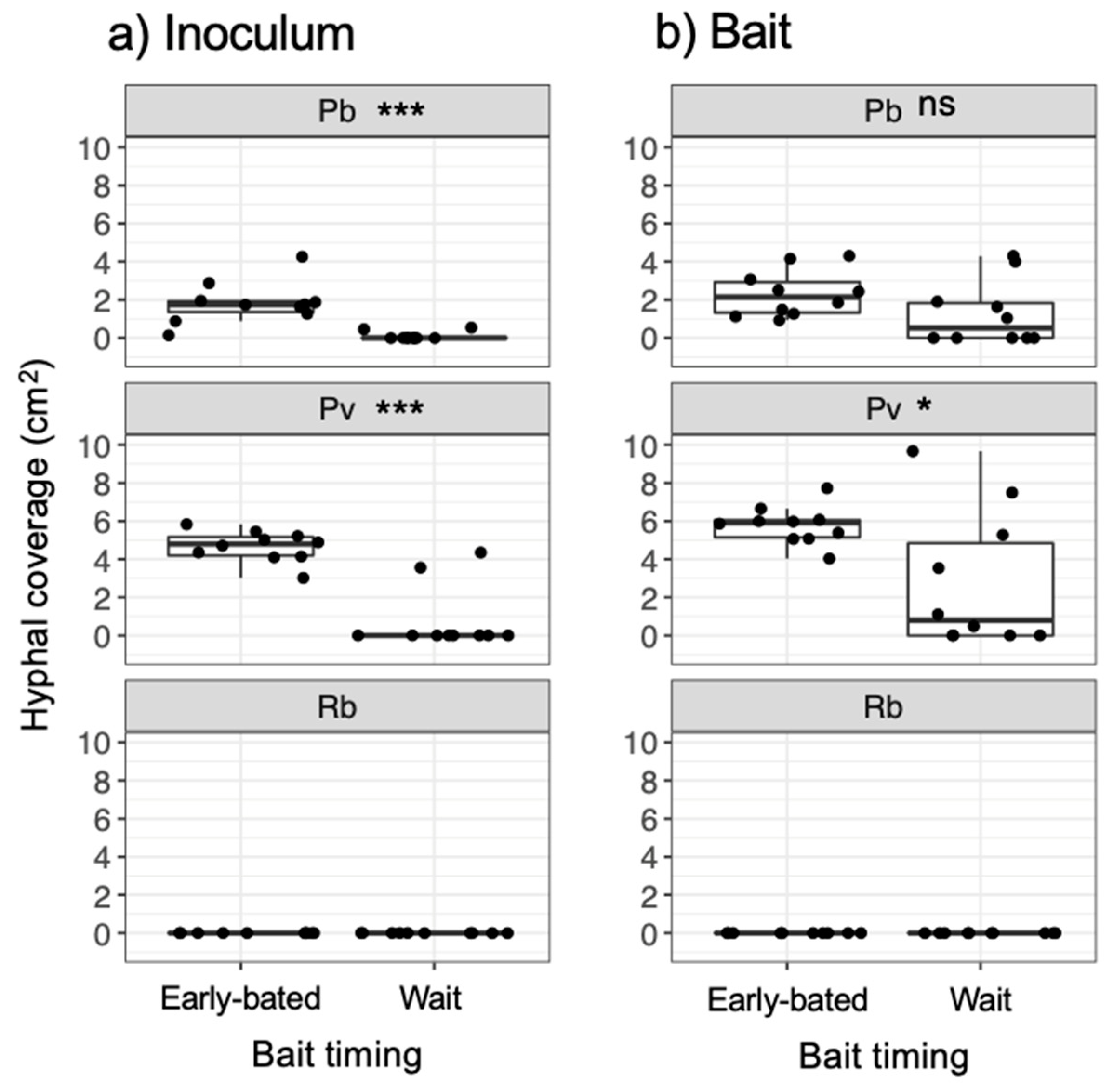
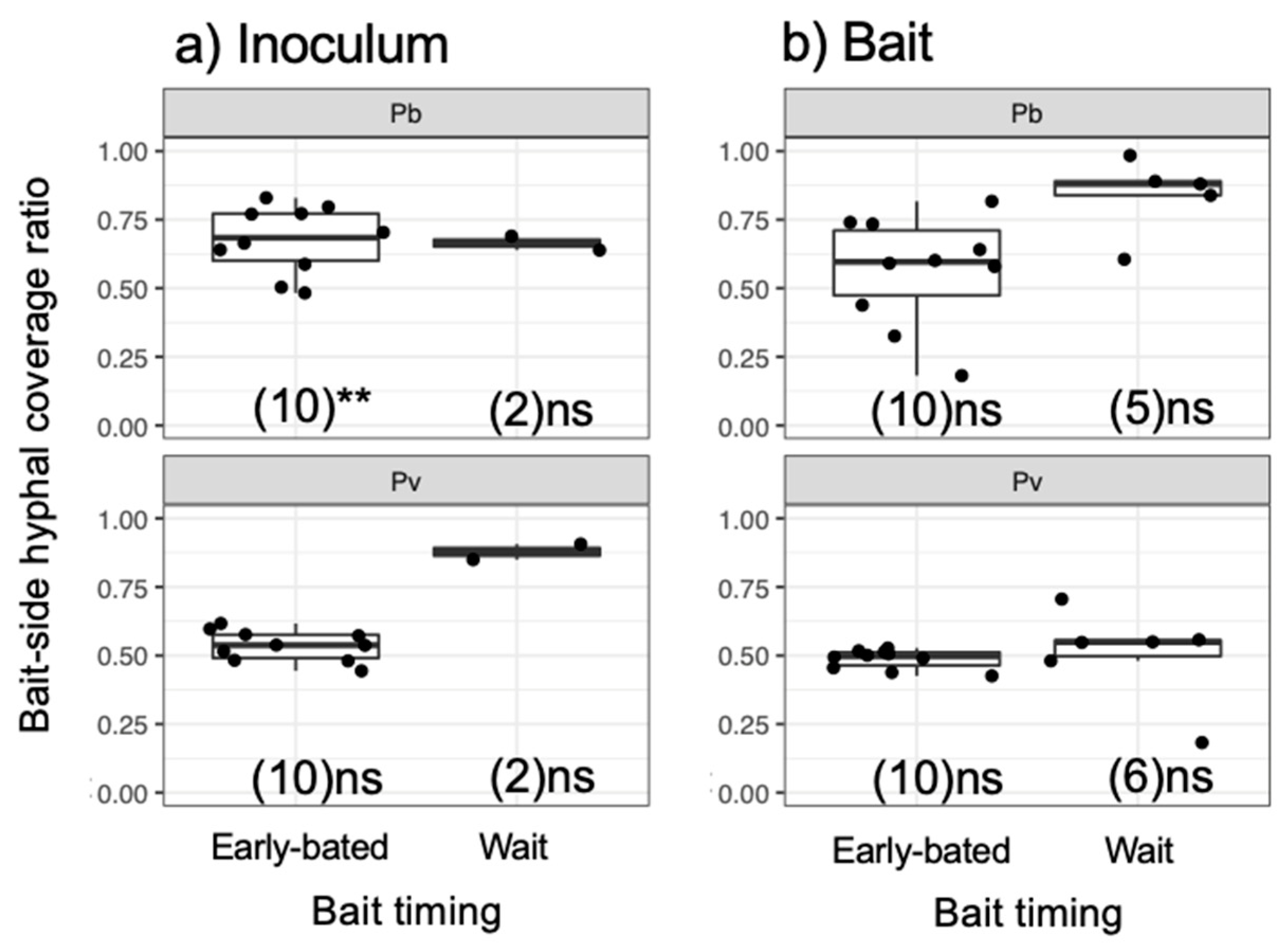
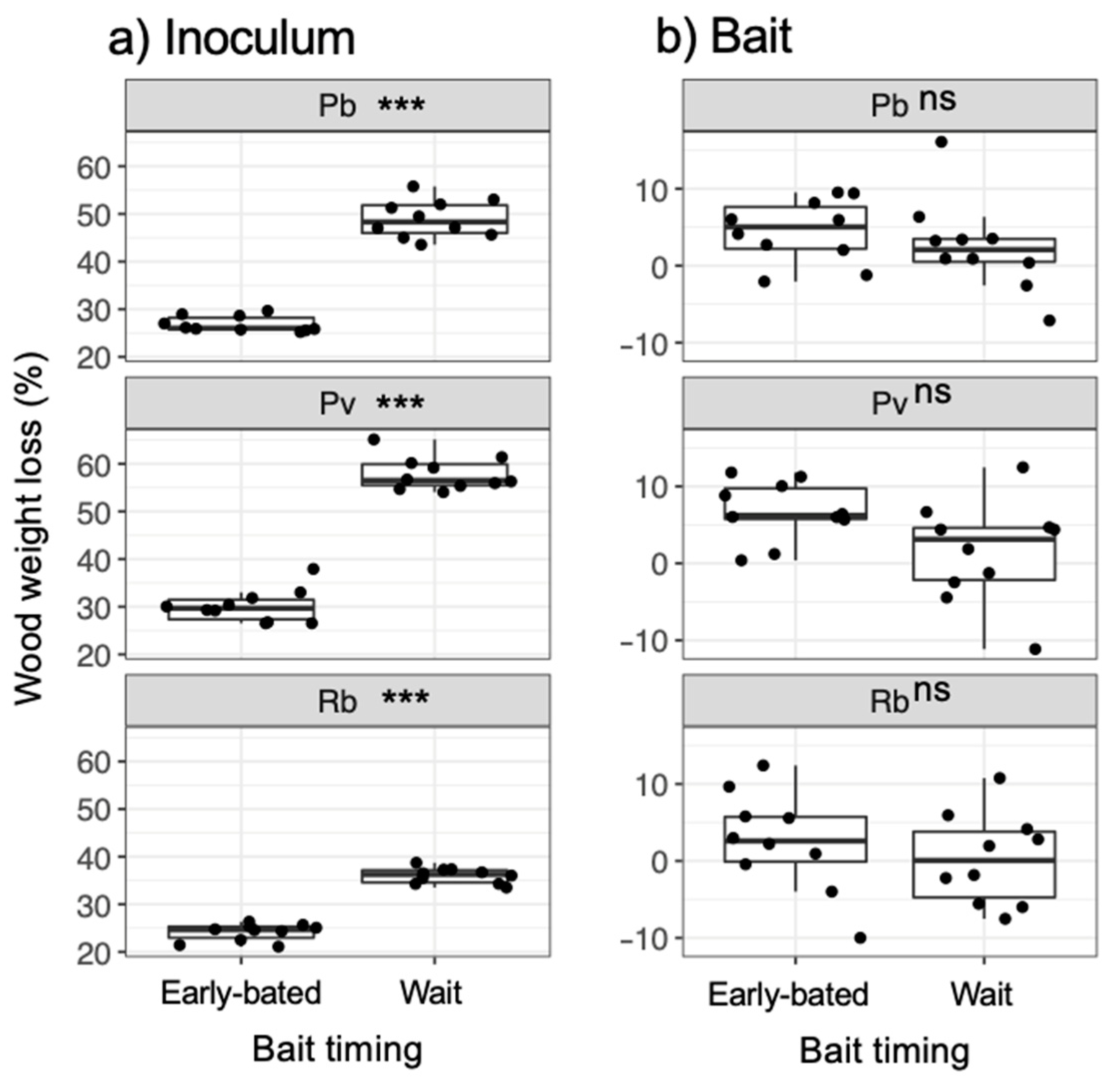
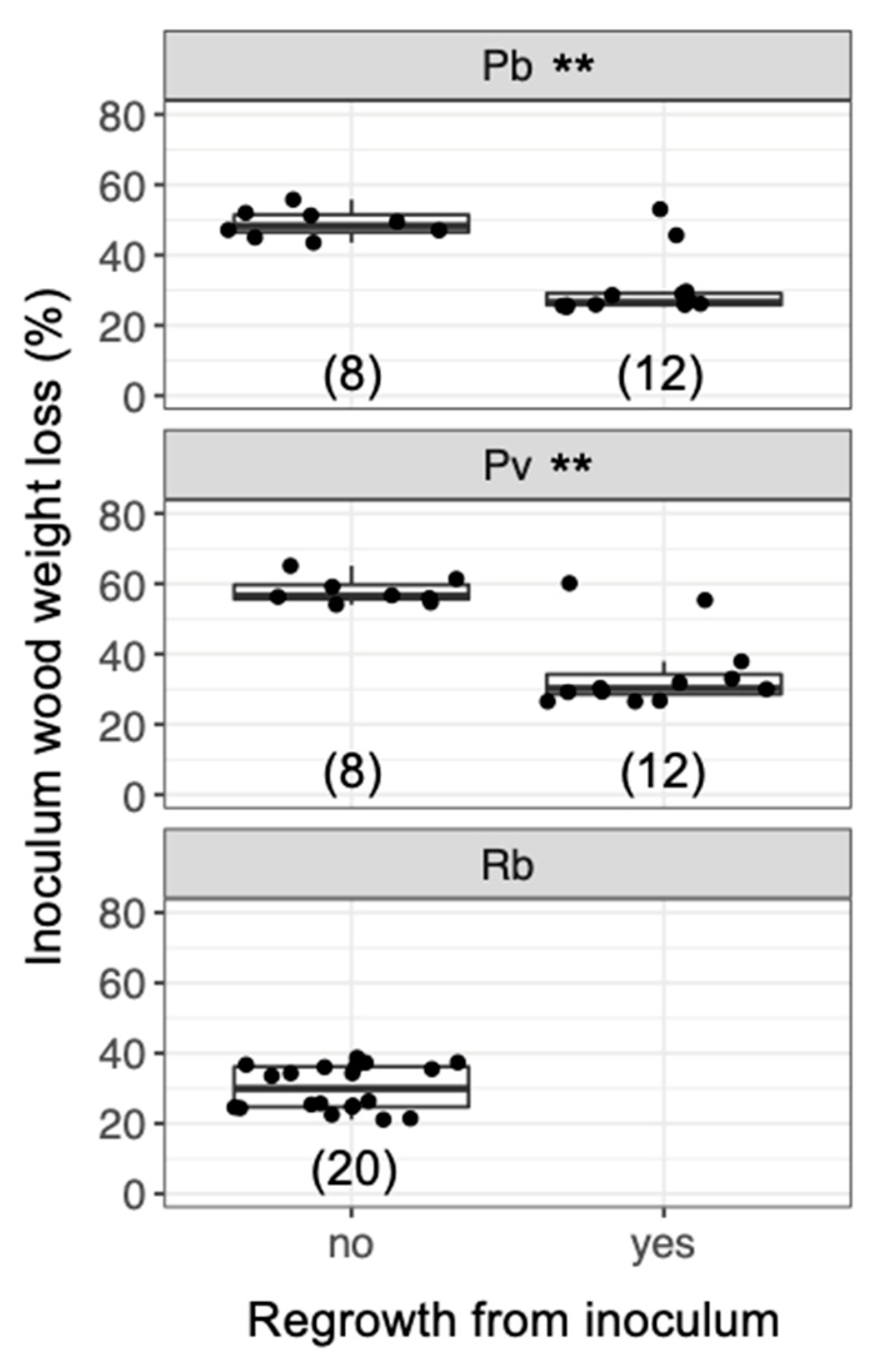
3. Results
4. Discussion
4.1. Effects of Bait Addition Timing on the Mycelial Decision to Migrate
4.2. Mycelial Memory of the Direction of Growth
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rayner, A.D.M.; Boddy, L. Fungal Decomposition of Wood: Its Biology and Ecology; John Wiley & Sons: Chichester, UK, 1988. [Google Scholar]
- Wilding, N.; Collins, N.M.; Hammond, P.M.; Webber, J.E. Insect-Fungus Interactions; Academic Press: Tokyo, Japan, 1989. [Google Scholar]
- Burslem, D.; Pinard, M.; Hartley, S. Biotic Interactions in the Tropics; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: Tokyo, Japan, 2008. [Google Scholar]
- Pirttilä, A.M.; Frank, A.C. Endophytes of Forest Trees; Springer: London, UK, 2011. [Google Scholar]
- Cairney, J.W.G. Translocation of solutes in ectomycorrhizal and saprotrophic rhizomorphs. Mycol. Res. 1992, 96, 135–141. [Google Scholar] [CrossRef]
- Lindahl, B.; Taylor, A.F.S.; Finlay, R.D. Defining nutritional constraints on carbon cycling in boreal forests—Towards a less ‘phytocentric’ perspective. Plant Soil. 2002, 242, 123–135. [Google Scholar] [CrossRef]
- Watkinson, S.; Bebber, D.; Darrah, P.; Fricker, M.; Tlalka, M.; Boddy, L. The role of wood decay fungi in the carbon and nitrogen dynamics of the forest floor. In Fungi in Biogeochemical Cycles; Gadd, G.M., Ed.; Cambridge University Press: Cambridge, UK, 2006; pp. 151–181. [Google Scholar]
- Boddy, L. Saprotrophic cord-forming fungi: Warfare strategies and other ecological aspects. Mycol. Res. 1993, 97, 641–655. [Google Scholar] [CrossRef]
- Bailer, K.J.; Durall, D.M.; Simard, S.W.; Maxell, S.A.; Kretzer, A.M. Architecture of the wood-wide web: Rhizopogon spp. genets link multiple Douglas-fir cohorts. New Phytol. 2010, 185, 543–553. [Google Scholar] [CrossRef]
- Crowther, T.W.; Maynard, D.S.; Crowther, T.R.; Peccia, J.; Smith, J.R.; Bradford, M.A. Untangling the fungal niche: The trait-based approach. Front. Microbiol. 2014, 5, 579. [Google Scholar] [CrossRef] [Green Version]
- Crowther, T.W.; Van den Hoogen, J.; Wan, J.; Mayes, M.A.; Keiser, A.D.; Mo, L.; Averill, C.; Maynard, D.S. The global soil community and its influence on biogeochemistry. Science 2019, 365, eaav0550. [Google Scholar] [CrossRef]
- Maynard, D.S.; Bradford, M.A.; Covey, K.R.; Lindner, D.; Glaeser, J.; Talbert, D.A.; Tinker, P.J.; Walker, D.M.; Crowther, T.W. Consistent trade-offs in fungal trait expression across broad spatial scales. Nat. Microbiol. 2019, 4, 846–853. [Google Scholar] [CrossRef]
- Crowther, T.W. The emergence of trait-based approaches in fungal ecology. Fung. Ecol. 2020, 46, 100946. [Google Scholar] [CrossRef]
- Lustenhouwer, N.; Maynard, D.S.; Bradford, M.A.; Lindner, D.L.; Oberle, B.; Zanne, A.E.; Crowther, T.W. A trait-based understanding of wood decomposition by fungi. Proc. Nat. Acad. Sci. USA 2020, 117, 11551–11558. [Google Scholar] [CrossRef] [PubMed]
- Dawson, S.K.; Boddy, L.; Halbwachs, H.; Bässler, C.; Andrew, C.; Crowther, T.W.; Heilmann-Clausen, J.; Nordén Ovaskainen, O.; Jönsson, M. Handbook for the measurement of macrofungal functional traits: A start with basidiomycete wood fungi. Func. Ecol. 2019, 33, 372–387. [Google Scholar] [CrossRef] [Green Version]
- Boddy, L. Saprotrophic cord-forming fungi: Meeting the challenge of heterogeneous environments. Mycologia 1999, 91, 13–32. [Google Scholar] [CrossRef]
- Boddy, L. Saprotrophic cord systems: Dispersal mechanisms in space and time. Mycoscience 2009, 50, 9–19. [Google Scholar] [CrossRef]
- Aleklett, K.; Boddy, L. Fungal behaviour: A new frontier in behavioural ecology. Trends Ecol. Evol. 2021, 36, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Money, N.P. Hyphal and mycelial consciousness: The concept of the fungal mind. Fungal Biol. 2021, 125, 257–259. [Google Scholar] [CrossRef] [PubMed]
- Fukasawa, Y.; Savoury, M.; Boddy, L. Ecological memory and relocation decisions in fungal mycelial networks: Responses to quantity and location of new resources. ISME J. 2020, 14, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Fukasawa, Y.; Kaga, K. Effects of wood resource size and decomposition on hyphal outgrowth of a cord-forming basidiomycete, Phanerochaete velutina. Sci. Rep. 2020, 10, 21936. [Google Scholar] [CrossRef] [PubMed]
- Held, M.; Kaspar, O.; Edwards, C.; Nicolau, D.V. Intracellular mechanisms of fungal space searching in microenvironments. Proc. Nat. Acad. Sci. USA 2019, 116, 13543–13552. [Google Scholar] [CrossRef] [Green Version]
- Aaronson, L.R.; Johnston, A.M.; Martin, C.E. The effects of temperature acclimation on membrane sterols and phospholipids of Neurospora crassa. Biochem. Biophysica. Acta 1982, 713, 456–462. [Google Scholar] [CrossRef]
- Addy, H.D.; Boswell, E.P.; Koide, R.T. Low temperature acclimation and freezing resistance of extraradical VA mycorrhizal hyphae. Mycol. Res. 1998, 102, 582–586. [Google Scholar] [CrossRef]
- Robinson, C.H. Cold adaptation in Arctic and Antarctic fungi. New Phytol. 2001, 151, 341–353. [Google Scholar] [CrossRef]
- Lenge, O.L.; Green, T.G.A. Lichen show that fungi can acclimate their respiration to seasonal change in temperature. Oecologia 2005, 142, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Malcolm, G.M.; López-Gutiérrez, J.C.; Koide, R.T.; Eissenstat, D.M. Acclimation to temperature and temperature sensitivity of metabolism by ectomycorrhizal fungi. Glob. Chan. Biol. 2008, 14, 1169–1180. [Google Scholar] [CrossRef]
- Crowther, T.W.; Bradford, M.A. Thermal acclimation in widespread heterotrophic soil microbes. Ecol. Lett. 2013, 16, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Andrade-Linares, D.R.; Veresoglou, S.D.; Rillig, M.C. Temperature priming and memory in soil filamentous fungi. Fungal Ecol. 2016, 21, 10–15. [Google Scholar] [CrossRef]
- Moncalvo, J.M.; Lutzoni, F.M.; Rehner, S.A.; Johnson, J.; Vilgalys, R. Phylogenetic relationships of agaric fungi based on nuclear large subunit ribosomal DNA sequences. Syst. Biol. 2000, 49, 278–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, E.; Matheny, P.B. A phylogenetic assessment of Pholiota and the new genus Pyrrhulomyces. Mycologia 2021, 113, 146–167. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; The R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Fricker, M.D.; Heaton, L.L.M.; Jones, N.S.; Boddy, L. The mycelium as a network. Microbiol. Spectr. 2017, 5, FUNK-0033-2017. [Google Scholar] [CrossRef]
- Wells, J.M.; Boddy, L. Wood decay, and phosphorus and fungal biomass allocation, in mycelial cord systems. New Phytol. 1990, 116, 285–295. [Google Scholar] [CrossRef]
- Wells, J.M.; Boddy, L. Translocation of soil-derived phosphorus in mycelial cord systems in relation to inoculum resource size. FEMS Microbiol. Ecol. 1995, 17, 67–75. [Google Scholar] [CrossRef]
- Wells, J.M.; Hughes, C.; Boddy, L. The fate of soil-derived phosphorus in mycelial cord systems of Phanerochaete velutina and Phallus impudicus. New Phytol. 1990, 114, 595–606. [Google Scholar] [CrossRef]
- Wells, J.M.; Boddy, L.; Evans, R. Carbon translocation in mycelial cord systems of Phanerochaete velutina (DC.: Pers.) Parmasto. New Phytol. 1995, 129, 467–476. [Google Scholar] [CrossRef]
- Hughes, C.L.; Boddy, L. Sequential encounter of wood resources by mycelial cord of Phanerochete velutina: Effect on growth patterns and phosphorus allocation. New Phytol. 1996, 133, 713–726. [Google Scholar] [CrossRef]
- Fricker, M.D.; Tlalka, M.; Bebber, D.; Tagaki, S.; Watkinson, S.C.; Darrah, P.R. Fourier-based spatial mapping of oscillatory phenomena in fungi. Fungal Gen. Biol. 2007, 44, 1077–1084. [Google Scholar] [CrossRef]
- Fricker, M.D.; Lee, J.A.; Bebber, D.P.; Tlalka, M.; Hynes, J.; Darrah, P.R.; Watkinson, S.C.; Boddy, L. Imaging complex nutrient dynamics in mycelial networks. J. Microsc. 2008, 231, 317–331. [Google Scholar] [CrossRef]
- Fricker, M.D.; Boddy, L.; Nakagaki, T.; Bebber, D.P. Adaptive biological networks. In Adaptive Networks: Theory, Models and Applications (Understanding Complex Systems); Gross, T., Sayama, H., Eds.; Springer: Heidelberg, Germany, 2009; pp. 51–70. [Google Scholar]
- Tlalka, M.; Bebber, D.P.; Darrah, P.R.; Watkinson, S.C.; Fricker, M.D. Emergence of self-organised oscillatory domains in fungal mycelia. Fungal Gen. Biol. 2007, 44, 1085–1095. [Google Scholar] [CrossRef]
- Herman, K.C.; Wosten, H.A.B.; Fricker, M.D.; Bleichrodt, R.J. Growth induced translocation effectively directs an amino acid analogue to developing zones in Agaricus bisporus. Fungal Biol. 2020, 124, 1013–1023. [Google Scholar] [CrossRef]
- Dussutour, A.; Latty, T.; Beekman, M.; Simpson, S.J. Amoeboid organism solves complex nutritional challenges. Proc. Nat. Acad. Sci. USA 2010, 107, 4607–4611. [Google Scholar] [CrossRef] [Green Version]
- Boddy, L.; Hiscox, J. Fungal ecology: Principles and mechanisms of colonization and competition by saprotrophic fungi. Microbiol. Spectr. 2016, 4, FUNK–0019–2016. [Google Scholar] [CrossRef] [Green Version]
- El Ariebi, N.; Hiscox, J.; Scriven, S.A.; Müller, C.T.; Boddy, L. Production and effects of volatile organic compounds during interspecific interactions. Fung. Ecol. 2016, 20, 144–154. [Google Scholar] [CrossRef]
- Hiscox, J.; Boddy, L. Armed and dangerous—Chemical warfare in wood decay communities. Fungal Biol. Rev. 2017, 31, 169–184. [Google Scholar] [CrossRef]
- Donnelly, D.P.; Boddy, L. Repeated damage results in polarised development of foraging mycelial systems of Phanerochaete velutina. FEMS Microbiol. Ecol. 1998, 26, 101–108. [Google Scholar] [CrossRef]
- Takamatsu, A.; Takaba, E.; Takizawa, G. Environment-dependent morphology in plasmodium of true slime mold Physarum polycepharum and a network growth model. J. Theor. Biol. 2009, 256, 29–44. [Google Scholar] [CrossRef] [PubMed]
- Rajala, T.; Peltoniemi, M.; Pennanen, T.; Mäkipää, R. Fungal community dynamics in relation to substrate quality of decaying Norway spruce (Picea abies [L.] Karst.) logs in boreal forests. FEMS Microbiol. Ecol. 2012, 81, 494–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hynes, J.; Müller, C.T.; Jones, T.H.; Boddy, L. Changes in volatile production during the course of fungal mycelial interactions between Hypoloma fasciculare and Resinicium bicolor. J. Chem. Ecol. 2007, 33, 43–57. [Google Scholar] [CrossRef]
- Crowther, T.W.; Boddy, L.; Jones, T.H. Outcomes of fungal interactions are determined by soil invertebrate grazers. Ecol. Lett. 2011, 14, 1134–1142. [Google Scholar] [CrossRef]
- Crowther, T.W.; Littleboy, A.; Jones, T.H.; Boddy, L. Interactive effects of warming and invertebrate grazing on the outcomes of competitive fungal interactions. FEMS Microbiol. Ecol. 2012, 81, 419–426. [Google Scholar] [CrossRef] [Green Version]
- A’Bear, A.D.; Crowther, T.W.; Ashfield, R.; Chadwick, D.D.; Dempsey, J.; Meletiou, L.; Rees, C.L.; Jones, T.H.; Boddy, L. Localised invertebrate grazing moderates the effect of warming on competitive fungal interactions. Fungal Ecol. 2013, 6, 137–140. [Google Scholar] [CrossRef]
- Hiscox, J.; Clarkson, G.; Savoury, M.; Powell, G.; Savva, I.; Lloyd, M.; Shipcott, J.; Choimes, A.; Cumbriu, X.A.; Boddy, L. Effects of pre-colonisation and temperature on interspecific fungal interactions in wood. Fungal Ecol. 2016, 21, 32–42. [Google Scholar] [CrossRef] [Green Version]
- Hanson, K.L.; Nicolau, D.V.; Filipponi, L.; Wang, L.; Lee, A.P.; Nicolau, D.V. Fungi use efficient algorithms for the exploration of microfluidic networks. Small 2006, 2, 1212–1220. [Google Scholar] [CrossRef]
- Held, M.; Edwards, C.; Nicolau, D.V. Probing the growth dynamics of Neurospora crassa with microfluidic structures. Fungal Biol. 2011, 115, 493–505. [Google Scholar] [CrossRef] [PubMed]
- Aleklett, K.; Ohlsson, P.; Bengtsson, M.; Hammer, E.C. Fungal foraging behaviour and hyphal space exploration in micro-structured Soil Chips. ISME J. 2021, 15, 1782–1793. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fukasawa, Y.; Kaga, K. Timing of Resource Addition Affects the Migration Behavior of Wood Decomposer Fungal Mycelia. J. Fungi 2021, 7, 654. https://doi.org/10.3390/jof7080654
Fukasawa Y, Kaga K. Timing of Resource Addition Affects the Migration Behavior of Wood Decomposer Fungal Mycelia. Journal of Fungi. 2021; 7(8):654. https://doi.org/10.3390/jof7080654
Chicago/Turabian StyleFukasawa, Yu, and Koji Kaga. 2021. "Timing of Resource Addition Affects the Migration Behavior of Wood Decomposer Fungal Mycelia" Journal of Fungi 7, no. 8: 654. https://doi.org/10.3390/jof7080654