
Profile of Membrane Cargo Trafficking Proteins and Transporters Expressed under N Source Derepressing Conditions in Aspergillus nidulans

 and
and 
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Media, Strains, Growth Conditions and A. nidulans Transformation
2.2. Protein Extraction
2.3. Enzymatic Digestion
2.4. One-Dimensional Liquid Chromatography (1D-LC) Separation
2.5. MS/MS Analysis
2.6. Data Analysis
2.7. Manual Curation and Protein Classification
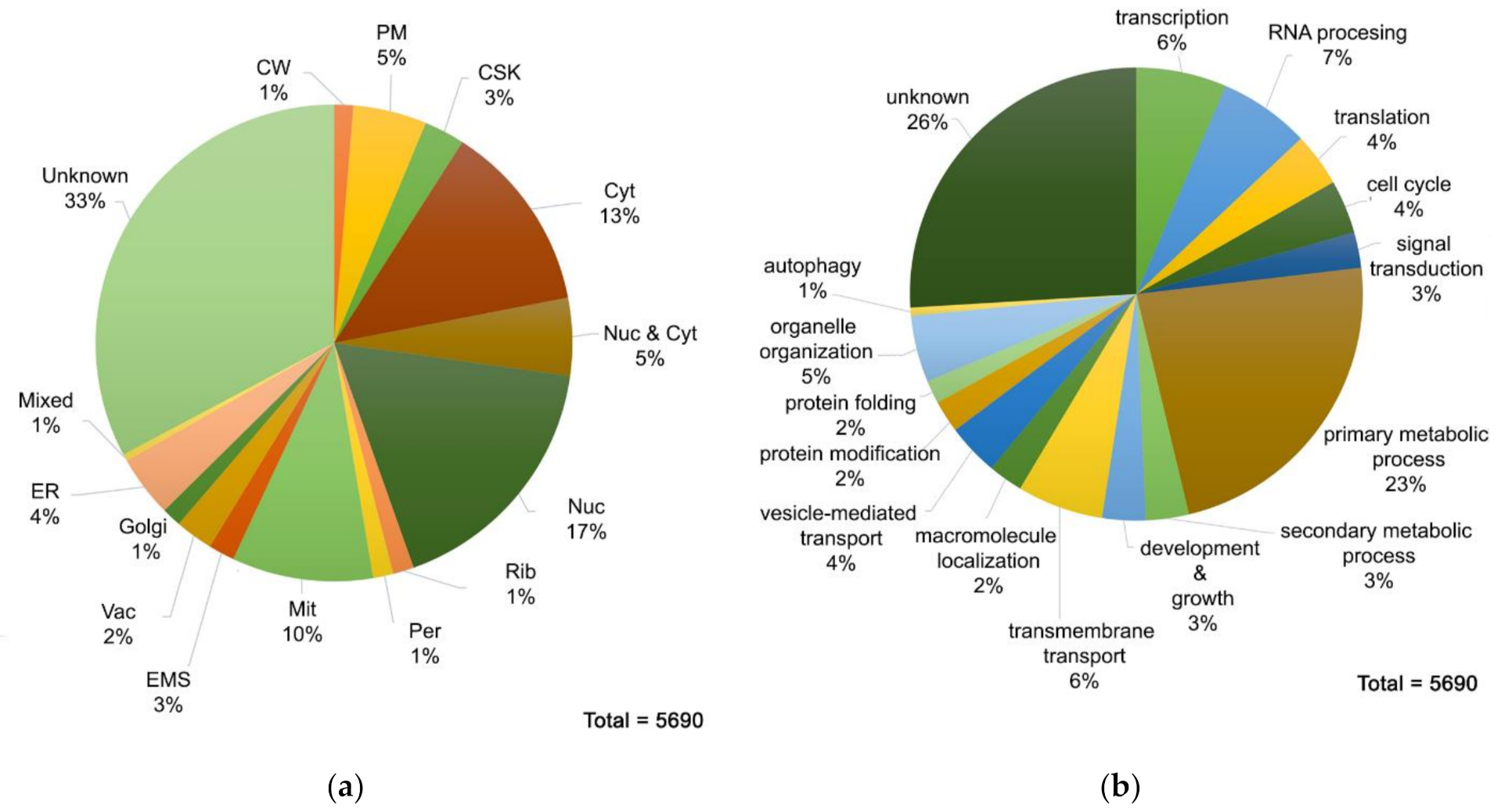
3. Results and Discussion
3.1. Development of a Protocol for Protein Extraction for Proteomic Analysis in A. nidulans
3.2. Proteome MS Analysis of A. nidulans Grown in Glucose MM Containing Nitrate as N Source
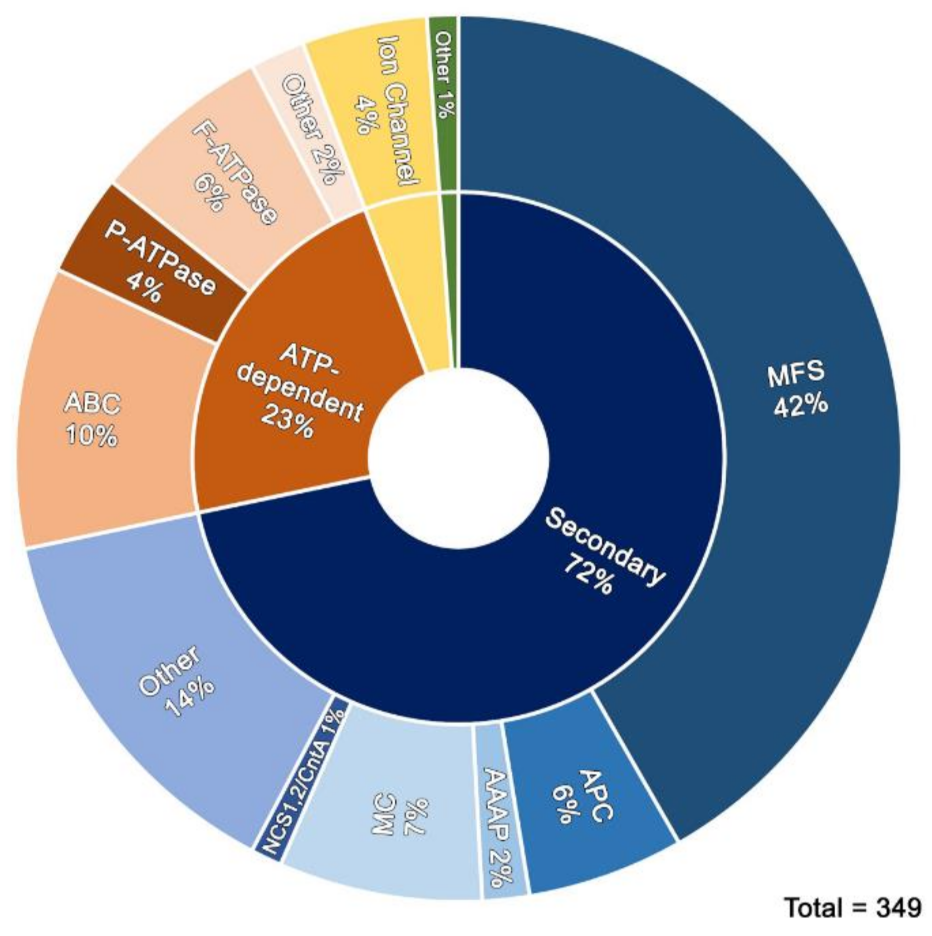
3.3. Transporters Identified
3.3.1. Sugar and Related Drug Efflux Transporters
3.3.2. Amino Acid Transporters
3.3.3. Nucleobase/Nucleoside Transporters
3.3.4. Other Transporters
3.3.5. Ion Channels
3.3.6. Mitochondrial Solute Transporters
3.3.7. Primary Active Transporters (ATP-Binding)
3.4. The Profile of Proteins Involved in Cargo Membrane Trafficking and Turnover Validates the High Sensitivity of the Proteome Identified
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Robey, R.W.; Pluchino, K.M.; Hall, M.D.; Fojo, A.T.; Bates, S.E.; Gottesman, M.M. Revisiting the role of ABC transporters in multidrug-resistant cancer. Nat. Rev. Cancer 2018, 18, 452–464. [Google Scholar] [CrossRef]
- Yahyaoui, R.; Pérez-Frías, J. Amino acid transport defects in human inherited metabolic disorders. Int. J. Mol. Sci. 2020, 21, 119. [Google Scholar] [CrossRef] [Green Version]
- Bowers, K.; Srai, S.K.S. The trafficking of metal ion transporters of the Zrt- and Irt-like protein family. Traffic 2018, 19, 813–822. [Google Scholar] [CrossRef] [Green Version]
- Bu, M.; Farrer, M.J.; Khoshbouei, H. Dynamic control of the dopamine transporter in neurotransmission and homeostasis. NPJ Park. Dis. 2021, 7, 1–11. [Google Scholar] [CrossRef]
- Evangelinos, M.; Martzoukou, O.; Chorozian, K.; Amillis, S.; Diallinas, G. BsdABsd2-dependent vacuolar turnover of a misfolded version of the UapA transporter along the secretory pathway: Prominent role of selective autophagy. Mol. Microbiol. 2016, 100, 893–911. [Google Scholar] [CrossRef]
- Cullen, P.J.; Steinberg, F. To degrade or not to degrade: Mechanisms and significance of endocytic recycling. Nat. Rev. Mol. Cell Biol. 2018, 19, 679–696. [Google Scholar] [CrossRef]
- Estadella, I.; Pedrós-Gámez, O.; Colomer-Molera, M.; Bosch, M.; Sorkin, A.; Felipe, A. Endocytosis: A Turnover Mechanism Controlling Ion Channel Function. Cells 2020, 9, 1833. [Google Scholar] [CrossRef]
- Dimou, S.; Diallinas, G. Life and death of fungal transporters under the challenge of polarity. Int. J. Mol. Sci. 2020, 21, 5376. [Google Scholar] [CrossRef]
- Barata-Antunes, C.; Alves, R.; Talaia, G.; Casal, M.; Gerós, H.; Mans, R.; Paiva, S. Endocytosis of nutrient transporters in fungi: The ART of connecting signaling and trafficking. Comput. Struct. Biotechnol. J. 2021, 21, 5376. [Google Scholar]
- Agasid, M.T.; Robinson, C.V. Probing membrane protein–lipid interactions. Curr. Opin. Struct. Biol. 2021, 69, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Feyder, S.; De Craene, J.O.; Bär, S.; Bertazzi, D.L.; Friant, S. Membrane trafficking in the yeast Saccharomyces cerevisiae model. Int. J. Mol. Sci. 2015, 16, 1509–1525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, N.J.; Booth, P.J. Co-Translational Protein Folding in Lipid Membranes. Trends Biochem. Sci. 2019, 44, 729–730. [Google Scholar] [CrossRef]
- Dimou, S.; Martzoukou, O.; Dionysopoulou, M.; Bouris, V.; Amillis, S.; Diallinas, G. Translocation of nutrient transporters to cell membrane via Golgi bypass in Aspergillus nidulans. EMBO Rep. 2020, 21, e49929. [Google Scholar] [CrossRef] [PubMed]
- Karachaliou, M.; Amillis, S.; Evangelinos, M.; Kokotos, A.C.; Yalelis, V.; Diallinas, G. The arrestin-like protein ArtA is essential for ubiquitination and endocytosis of the UapA transporter in response to both broad-range and specific signals. Mol. Microbiol. 2013, 88, 301–317. [Google Scholar] [CrossRef]
- Martzoukou, O.; Amillis, S.; Zervakou, A.; Christoforidis, S.; Diallinas, G. The AP-2 complex has a specialized clathrin-independent role in apical endocytosis and polar growth in fungi. Elife 2017, 6, e20083. [Google Scholar] [CrossRef] [PubMed]
- Gingras, A.C.; Abe, K.T.; Raught, B. Getting to know the neighborhood: Using proximity-dependent biotinylation to characterize protein complexes and map organelles. Curr. Opin. Chem. Biol. 2019, 48, 44–54. [Google Scholar] [CrossRef]
- Arora, D.; Abel, N.B.; Liu, C.; van Damme, P.; Yperman, K.; Eeckhout, D.; Vu, L.D.; Wang, J.; Tornkvist, A.; Impens, F.; et al. Establishment of proximity-dependent biotinylation approaches in different plant model systems. Plant Cell 2020, 32, 3388–3407. [Google Scholar] [CrossRef] [PubMed]
- Pantazopoulou, A.; Lemuh, N.D.; Hatzinikolaou, D.G.; Drevet, C.; Cecchetto, G.; Scazzocchio, C.; Diallinas, G. Differential physiological and developmental expression of the UapA and AzgA purine transporters in Aspergillus nidulans. Fungal Genet. Biol. 2007, 44, 627–640. [Google Scholar] [CrossRef]
- Pantazopoulou, A.; Diallinas, G. Fungal nucleobase transporters. FEMS Microbiol. Rev. 2007, 31, 657–675. [Google Scholar] [CrossRef] [Green Version]
- Koukaki, M.; Giannoutsou, E.; Karagouni, A.; Diallinas, G. A novel improved method for Aspergillus nidulans transformation. J. Microbiol. Methods 2003, 55, 687–695. [Google Scholar] [CrossRef]
- Diallinas, G. Biochemistry: An almost-complete movie. Science 2008, 322, 1644–1645. [Google Scholar] [CrossRef]
- Anagnostopoulos, A.K.; Stravopodis, D.J.; Tsangaris, G.T. Yield of 6,000 proteins by 1D nLC–MS/MS without pre-fractionation. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2017, 1047, 92–96. [Google Scholar] [CrossRef]
- Forment, J.V.; Flipphi, M.; Ventura, L.; González, R.; Ramón, D.; MacCabe, A.P. High-affinity glucose transport in Aspergillus nidulans is mediated by the products of two related but differentially expressed genes. PLoS ONE 2014, 9, e94662. [Google Scholar] [CrossRef] [Green Version]
- dos Reis, T.F.; Menino, J.F.; Bom, V.L.P.; Brown, N.A.; Colabardini, A.C.; Savoldi, M.; Goldman, M.H.S.; Rodrigues, F.; Goldman, G.H. Identification of glucose transporters in aspergillus nidulans. PLoS ONE 2013, 8, e81412. [Google Scholar] [CrossRef] [Green Version]
- Forment, J.V.; Flipphi, M.; Ramón, D.; Ventura, L.; MacCabe, A.P. Identification of the mstE gene encoding a glucose-inducible, low affinity glucose transporter in Aspergillus nidulans. J. Biol. Chem. 2006, 281, 8339–8346. [Google Scholar] [CrossRef] [Green Version]
- Fekete, E.; Karaffa, L.; Seiboth, B.; Fekete, É.; Kubicek, C.P.; Flipphi, M. Identification of a permease gene involved in lactose utilisation in Aspergillus nidulans. Fungal Genet. Biol. 2012, 49, 415–425. [Google Scholar] [CrossRef]
- Colabardini, A.C.; Ries, L.N.A.; Brown, N.A.; Dos Reis, T.F.; Savoldi, M.; Goldman, M.H.S.; Menino, J.F.; Rodrigues, F.; Goldman, G.H. Functional characterization of a xylose transporter in Aspergillus nidulans. Biotechnol. Biofuels 2014, 7, 46. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.; Vienken, K.; Weber, R.; Bunting, S.; Requena, N.; Fischer, R. A putative high affinity hexose transporter, hxtA, of Aspergillus nidulans is induced in vegetative hyphae upon starvation and in ascogenous hyphae during cleistothecium formation. Fungal Genet. Biol. 2004, 41, 148–156. [Google Scholar] [CrossRef]
- Dos Reis, T.F.; De Lima, P.B.A.; Parachin, N.S.; Mingossi, F.B.; De Castro Oliveira, J.V.; Ries, L.N.A.; Goldman, G.H. Identification and characterization of putative xylose and cellobiose transporters in Aspergillus nidulans. Biotechnol. Biofuels 2016, 9, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Balaska, S.; Myrianthopoulos, V.; Tselika, M.; Hatzinikolaou, D.G.; Mikros, E.; Diallinas, G. NmeA, a novel efflux transporter specific for nucleobases and nucleosides, contributes to metal resistance in Aspergillus nidulans. Mol. Microbiol. 2017, 05, 426–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, H. Molecular genetics of fungal siderophore biosynthesis and uptake: The role of siderophores in iron uptake and storage. Appl. Microbiol. Biotechnol. 2003, 62, 316–330. [Google Scholar] [CrossRef]
- Gournas, C.; Evangelidis, T.; Athanasopoulos, A.; Mikros, E.; Sophianopoulou, V. The Aspergillus nidulans proline permease as a model for understanding the factors determining substrate binding and specificity of fungal amino acid transporters. J. Biol. Chem. 2015, 290, 6141–6155. [Google Scholar] [CrossRef] [Green Version]
- Apostolaki, A.; Erpapazoglou, Z.; Harispe, L.; Billini, M.; Kafasla, P.; Kizis, D.; Peñalva, M.A.; Scazzocchio, C.; Sophianopoulou, V. AgtA, the dicarboxylic amino acid transporter of Aspergillus nidulans, is concertedly down-regulated by exquisite sensitivity to nitrogen metabolite repression and ammonium-elicited endocytosis. Eukaryot. Cell 2009, 8, 339–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutchings, H.; Stahmann, K.P.; Roels, S.; Espeso, E.A.; Timberlake, W.E.; Arst, H.N.; Tilburn, J. The multiply-regulated gabA gene encoding the GABA permease of Aspergillus nidulans: A score of exons. Mol. Microbiol. 1999, 32, 557–568. [Google Scholar] [CrossRef] [PubMed]
- Russnak, R.; Konczal, D.; McIntire, S.L. A Family of Yeast Proteins Mediating Bidirectional Vacuolar Amino Acid Transport. J. Biol. Chem. 2001, 276, 23849–23857. [Google Scholar] [CrossRef] [Green Version]
- Krypotou, E.; Evangelidis, T.; Bobonis, J.; Pittis, A.A.; Gabaldón, T.; Scazzocchio, C.; Mikros, E.; Diallinas, G. Origin, diversification and substrate specificity in the family of NCS1/FUR transporters. Mol. Microbiol. 2015, 96, 927–950. [Google Scholar] [CrossRef]
- Sioupouli, G.; Lambrinidis, G.; Mikros, E.; Amillis, S.; Diallinas, G. Cryptic purine transporters in Aspergillus nidulans reveal the role of specific residues in the evolution of specificity in the NCS1 family. Mol. Microbiol. 2017, 103, 319–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gournas, C.; Papageorgiou, I.; Diallinas, G. The nucleobase-ascorbate transporter (NAT) family: Genomics, evolution, structure-function relationships and physiological role. Mol. Biosyst. 2008, 4, 404–416. [Google Scholar] [CrossRef]
- Diallinas, G.; Gournas, C. Structure-function relationships in the nucleobase-ascorbate transporter (NAT) family: Lessons from model microbial genetic systems. Channels 2008, 2, 363–372. [Google Scholar] [CrossRef]
- Hamari, Z.; Amillis, S.; Drevet, C.; Apostolaki, A.; Vágvölgyi, C.; Diallinas, G.; Scazzocchio, C. Convergent evolution and orphan genes in the Fur4p-like family and characterization of a general nucleoside transporter in Aspergillus nidulans. Mol. Microbiol. 2009, 73, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Branon, T.C.; Bosch, J.A.; Sanchez, A.D.; Udeshi, N.D.; Svinkina, T.; Carr, S.A.; Feldman, J.L.; Perrimon, N.; Ting, A.Y. Efficient proximity labeling in living cells and organisms with TurboID. Nat. Biotechnol. 2018, 36, 880–887. [Google Scholar] [CrossRef]
- Wang, Y.; Li, W.; Siddiqi, Y.; Symington, V.F.; Kinghorn, J.R.; Unkles, S.E.; Glass, A.D.M. Nitrite transport is mediated by the nitrite-specific high-affinity NitA transporter and by nitrate transporters NrtA, NrtB in Aspergillus nidulans. Fungal Genet. Biol. 2008, 45, 94–102. [Google Scholar] [CrossRef]
- Sá-Pessoa, J.; Amillis, S.; Casal, M.; Diallinas, G. Expression and specificity profile of the major acetate transporter AcpA in Aspergillus nidulans. Fungal Genet. Biol. 2015, 76, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Monahan, B.J.; Askin, M.C.; Hynes, M.J.; Davis, M.A. Differential expression of Aspergillus nidulans ammonium permease genes is regulated by GATA transcription factor AreA. Eukaryot. Cell 2006, 5, 226–237. [Google Scholar] [CrossRef] [Green Version]
- Del Sorbo, G.; Andrade, A.C.; Van Nistelrooy, J.G.M.; Van Kan, J.A.L.; Balzi, E.; De Waard, M.A. Multidrug resistance in Aspergillus nidulans involves novel ATP-binding cassette transporters. Mol. Gen. Genet. 1997, 254, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Andrade, A.C.; Del Sorbo, G.; Van Nistelrooy, J.G.M.; De Waard, M.A. The ABC transporter AtrB from Aspergillus nidulans mediates resistance to all major classes of fungicides and some natural toxic compounds. Microbiology 2000, 146, 1987–1997. [Google Scholar] [CrossRef] [Green Version]
- Chiang, Y.M.; Ahuja, M.; Oakley, C.E.; Entwistle, R.; Asokan, A.; Zutz, C.; Wang, C.C.C.; Oakley, B.R. Development of Genetic Dereplication Strains in Aspergillus nidulans Results in the Discovery of Aspercryptin. Angew. Chemie Int. Ed. 2016, 55, 1662–1665. [Google Scholar] [CrossRef] [Green Version]
- Reoyo, E.; Espeso, E.A.; Peñalva, M.A.; Suárez, T. The essential Aspergillus nidulans gene pmaA encodes an homologue of fungal plasma membrane H+-ATPases. Fungal Genet. Biol. 1998, 23, 288–299. [Google Scholar] [CrossRef]
- Clutterbuck, A.J. The genetics of conidiophore pigmentation in Aspergillus nidulans. J. Gen. Microbiol. 1990, 136, 1731–1738. [Google Scholar] [CrossRef] [Green Version]
- Markina-Iñarrairaegui, A.; Spielvogel, A.; Etxebeste, O.; Ugalde, U.; Espeso, E.A. Tolerance to alkaline ambient pH in Aspergillus nidulans depends on the activity of ENA proteins. Sci. Rep. 2020, 10, 14325. [Google Scholar] [CrossRef]
- Melin, P.; Schnürer, J.; Wagner, E.G.H. Disruption of the gene encoding the V-ATPase subunit A results in inhibition of normal growth and abolished sporulation in Aspergillus nidulans. Microbiology 2004, 150, 743–748. [Google Scholar] [CrossRef]
- Bravo-Plaza, I.; Hernández-González, M.; Pinar, M.; Díaz, J.F.; Peñalva, M.A. Identification of the guanine nucleotide exchange factor for SAR1 in the filamentous fungal model Aspergillus nidulans. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 118551. [Google Scholar] [CrossRef]
- Hovsepian, J.; Albanèse, V.; Becuwe, M.; Ivashov, V.; Teis, D.; Léon, S. The yeast arrestin-related protein Bul1 is a novel actor of glucose-induced endocytosis. Mol. Biol. Cell 2018, 29, 1012–1020. [Google Scholar] [CrossRef]
- Peñalva, M.A.; Lucena-Agell, D.; Arst, H.N. Liaison alcaline: Pals entice non-endosomal ESCRTs to the plasma membrane for pH signaling. Curr. Opin. Microbiol. 2014, 22, 49–59. [Google Scholar] [CrossRef]
- Tsukada, M.; Ohsumi, Y. Isolation and characterization of autophagy-defective mutants of Saccharomyces cerevisiae. FEBS Lett. 1993, 333, 169–174. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dimou, S.; Georgiou, X.; Sarantidi, E.; Diallinas, G.; Anagnostopoulos, A.K. Profile of Membrane Cargo Trafficking Proteins and Transporters Expressed under N Source Derepressing Conditions in Aspergillus nidulans. J. Fungi 2021, 7, 560. https://doi.org/10.3390/jof7070560
Dimou S, Georgiou X, Sarantidi E, Diallinas G, Anagnostopoulos AK. Profile of Membrane Cargo Trafficking Proteins and Transporters Expressed under N Source Derepressing Conditions in Aspergillus nidulans. Journal of Fungi. 2021; 7(7):560. https://doi.org/10.3390/jof7070560
Chicago/Turabian StyleDimou, Sofia, Xenia Georgiou, Eleana Sarantidi, George Diallinas, and Athanasios K. Anagnostopoulos. 2021. "Profile of Membrane Cargo Trafficking Proteins and Transporters Expressed under N Source Derepressing Conditions in Aspergillus nidulans" Journal of Fungi 7, no. 7: 560. https://doi.org/10.3390/jof7070560