
Polyphasic Approach Utilized for the Identification of Two New Toxigenic Members of Penicillium Section Exilicaulis, P. krskae and P. silybi spp. nov.


 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Sample Collection and Isolation of the Fungi
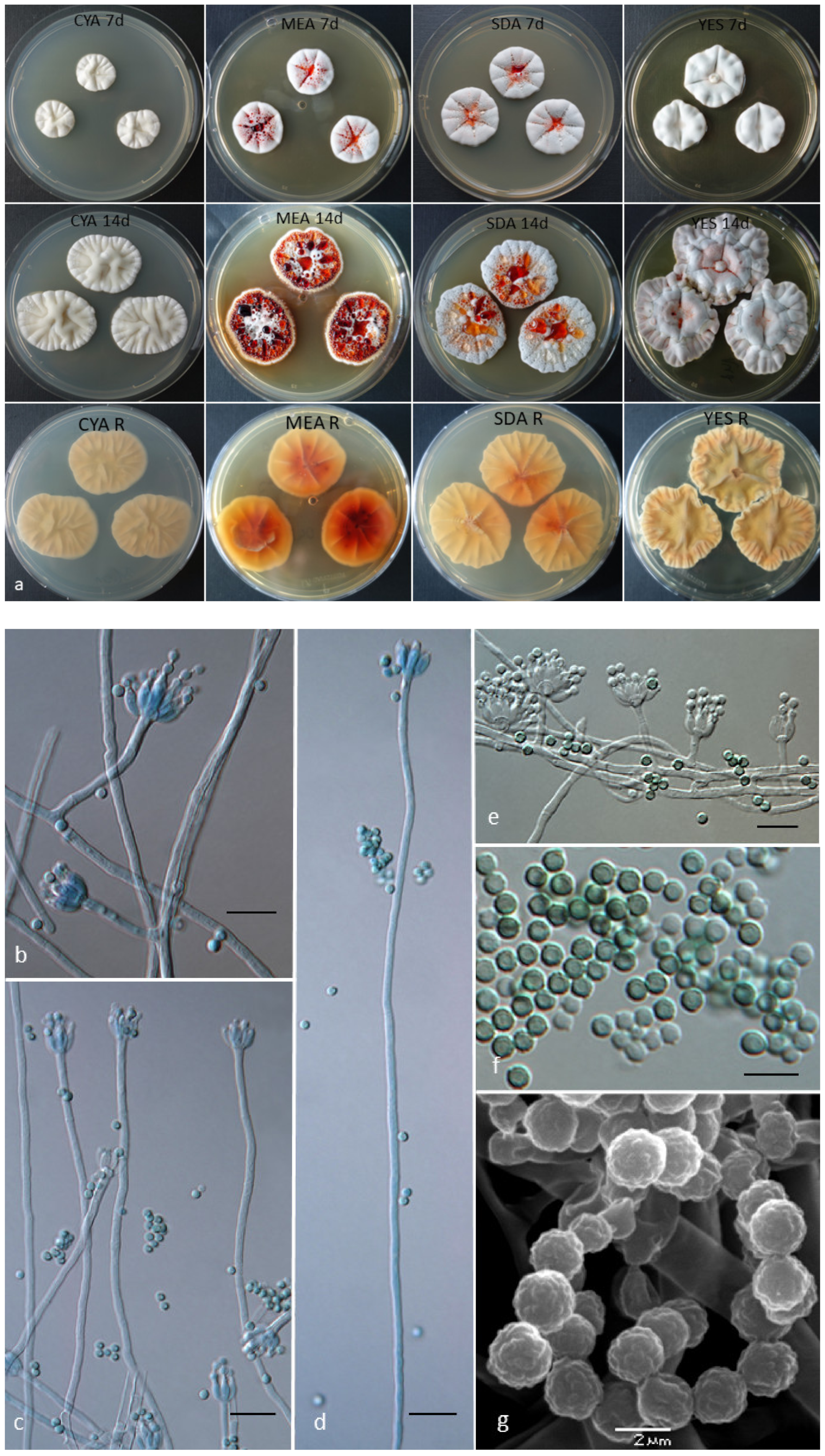
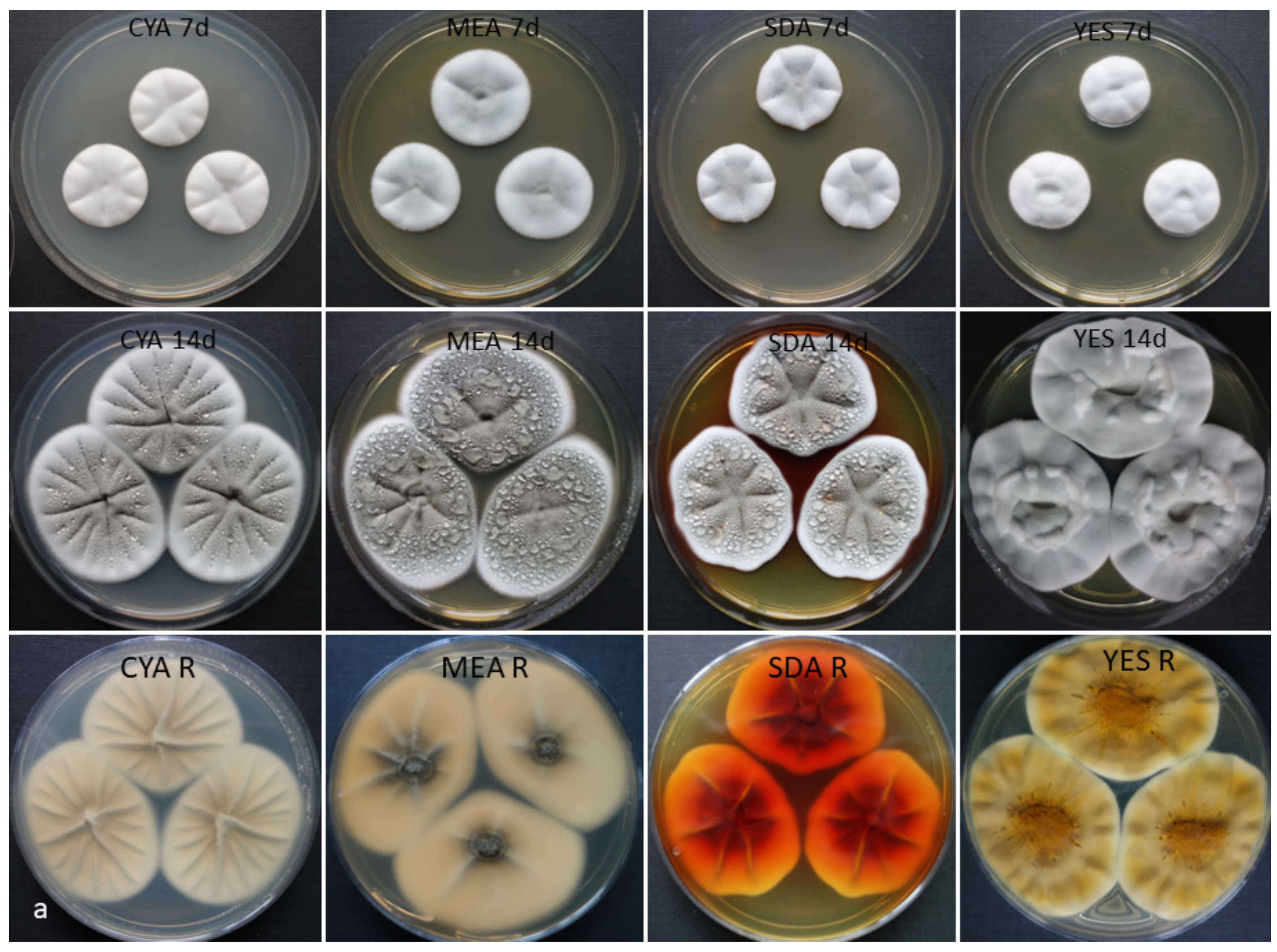
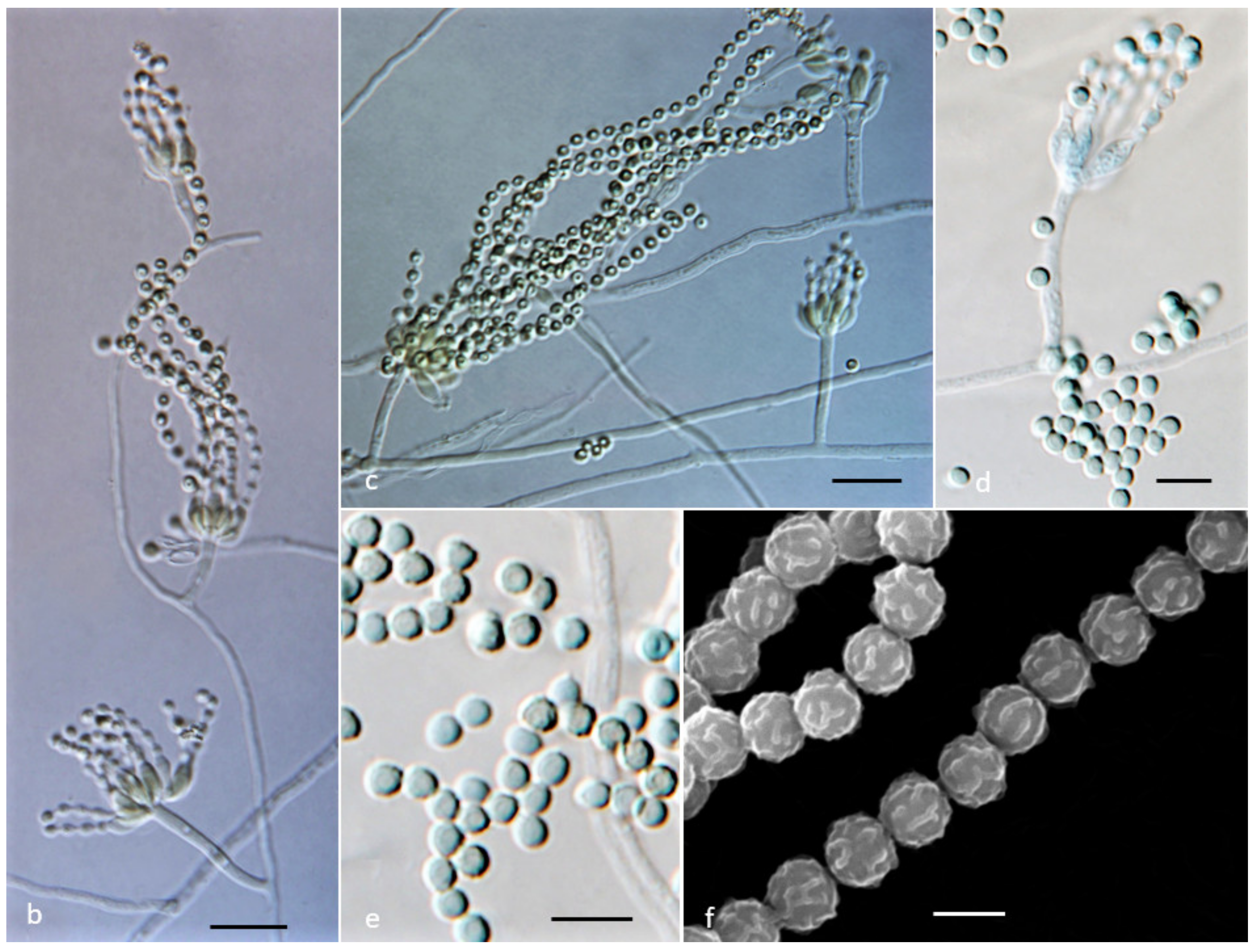
2.2. Cultivation of the Strains, Media, and Morphological Analysis
2.3. DNA Extraction, PCR Amplification, and Sequencing
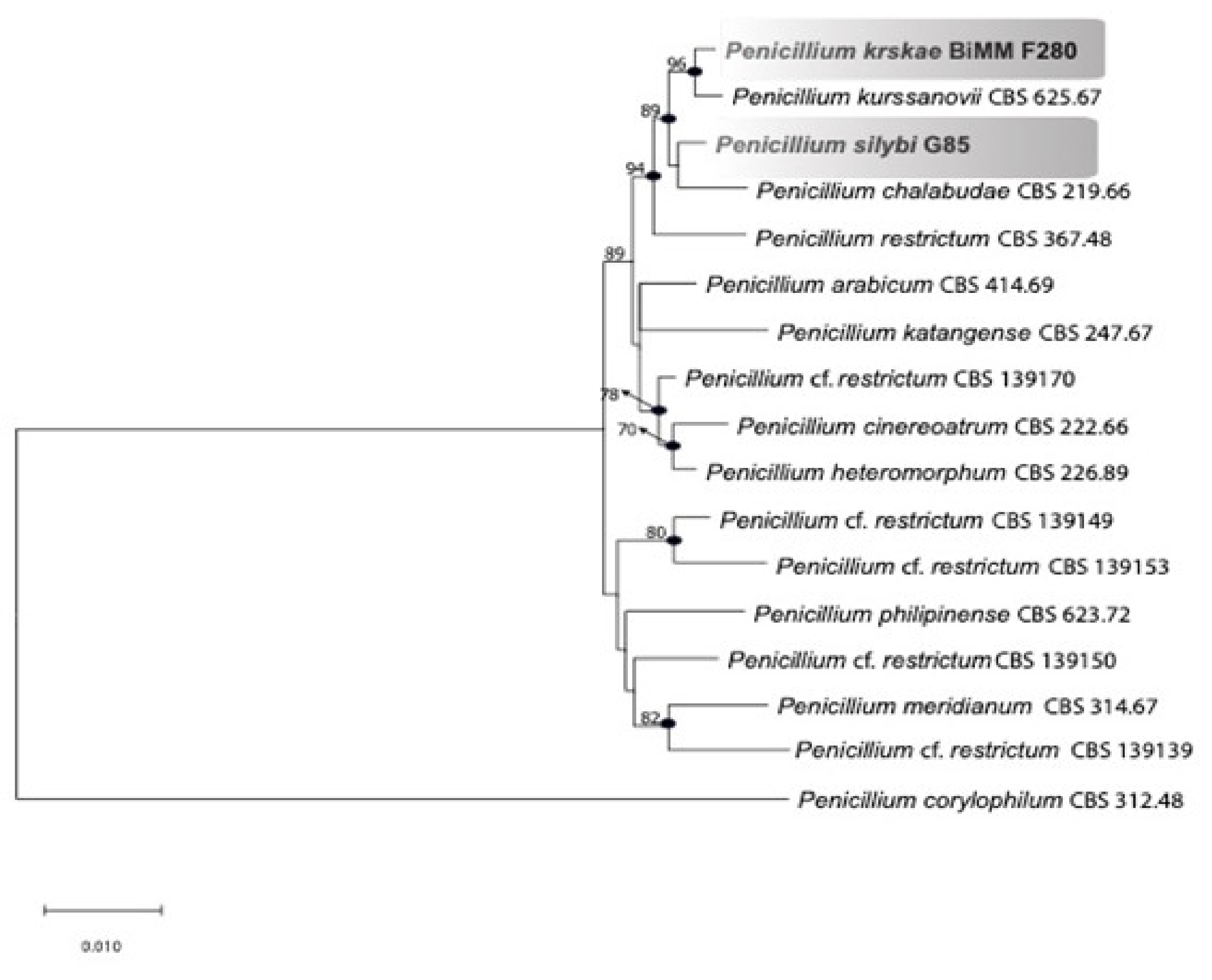
2.4. Phylogenetic Analysis
2.4.1. Evolutionary Analysis Using Maximum Likelihood
2.4.2. Bayesian Analysis
2.5. Chemical Analysis of the Red Exudate and Metabolic Profiling
2.5.1. Fermentation and Extraction
2.5.2. Micro-Extraction of YES Cultures for Metabolic Profiling and Quantification
2.5.3. Isolation of Secondary Metabolites from Red Exudate
2.5.4. LC–MS Analysis of the Exudate
2.5.5. Multi-Toxin LC–MS/MS Method for the YES Extract Quantification
2.5.6. NMR
2.5.7. Confirmation of Novel Compounds by LC–MS
3. Results
3.1. Taxonomy
3.2. Phylogenetic Analysis
3.3. Phenotype and Physiological Characteristics
3.4. Secondary Metabolites and Novel Chloroemodic Acid Derivatives
3.4.1. Chemical Profiling of the Exudate
3.4.2. Metabolic Profile of the Related Penicillium Species in Series Restricta
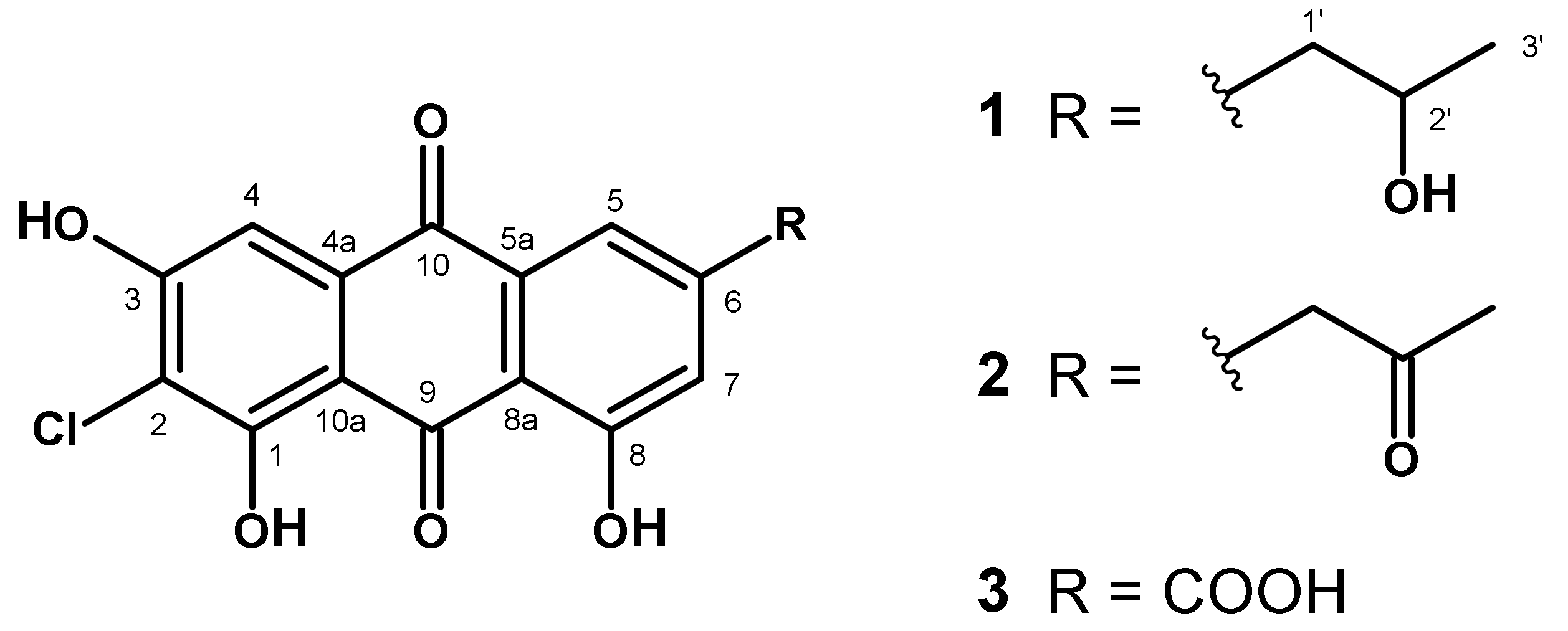
3.4.3. Two Novel Chloroemodic Acid Derivatives
4. Discussion
4.1. Phylogeny
4.2. Phenotype and Physiology
4.3. Extrolites
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Houbraken, J.; Kocsube, S.; Visagie, C.M.; Yilmaz, N.; Wang, X.C.; Meijer, M.; Kraak, B.; Hubka, V.; Bensch, K.; Samson, R.A.; et al. Classification of Aspergillus, Penicillium, Talaromyces and related genera (Eurotiales): An overview of families, genera, subgenera, sections, series and species. Stud. Mycol. 2020, 95, 5–169. [Google Scholar] [CrossRef]
- Cole, R.A.; Cox, R.H. Handbook of Toxic Fungal Metabolites; Academic Press: New York, NY, USA, 1981; p. 937. [Google Scholar]
- Betina, V. Mycotoxins: Production, Isolation, Separation, and Purification; Elsevier: Amsterdam, The Netherlands, 1989. [Google Scholar]
- Pitt, J.I. Biology and ecology of toxigenic Penicillium species. In Mycotoxins and Food Safety; DeVries, J.W., Trucksess, M.W., Jackson, L.S., Eds.; Kluwer Academic: New York, NY, USA, 2002; pp. 29–41. [Google Scholar]
- Nielsen, J.C.; Grijseels, S.; Prigent, S.; Ji, B.; Dainat, J.; Nielsen, K.F.; Frisvad, J.C.; Workman, M.; Nielsen, J. Global analysis of biosynthetic gene clusters reveals vast potential of secondary metabolite production in Penicillium species. Nat. Microbiol. 2017, 2, 17044. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.W.; Klich, M. Mycotoxins. Clinical Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarvis, B.B.; Miller, J.D. Mycotoxins as harmful indoor air contaminants. Appl. Microbiol. Biotechnol. 2005, 66, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Samson, R.A.; Houbraken, J.; Thrane, U.; Frisvad, J.C.; Andersen, B. Food and Indoor Fungi, 2nd ed.; Westerdijk Fungal Biodiversity Institute: Utrecht, The Netherlands, 2019; p. 481. [Google Scholar]
- Houbraken, J.; Samson, R.A. Phylogeny of Penicillium and the segregation of Trichocomaceae into three families. Stud. Mycol. 2011, 70, 1–51. [Google Scholar] [CrossRef]
- Houbraken, J.; Visagie, C.M.; Meijer, M.; Frisvad, J.C.; Busby, P.E.; Pitt, J.I.; Seifert, K.A.; Louis-Seize, G.; Demirel, R.; Yilmaz, N.; et al. A taxonomic and phylogenetic revision of Penicillium section Aspergilloides. Stud. Mycol. 2014, 78, 373–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visagie, C.M.; Seifert, K.A.; Houbraken, J.; Samson, R.A.; Jacobs, K. A phylogenetic revision of Penicillium sect. Exilicaulis, including nine new species from fynbos in South Africa. IMA Fungus 2016, 7, 75–117. [Google Scholar] [CrossRef] [Green Version]
- Pitt, J.I. The Genus Penicillium and its Teleomorphic States Eupenicillium and Talaromyces; Academic Press: London, UK, 1980; p. 634. [Google Scholar]
- Domsch, K.H.; Gams, W.; Anderson, T.H. Compendium of Soil Fungi; Academic Press: London, UK, 1980. [Google Scholar]
- Guarro, J.; Gené, J.; Stchigel, A.M.; Figueras, M.J. Atlas of Soil Ascomycetes; APS Press: St. Paul, MN, USA, 2012. [Google Scholar]
- De Hoog, G.S.; Guarro, J.G.; Figueras, M.J. Atlas of Clinical Fungi, 2nd ed.; Centraalbureau voor Schimmelcultures and Universitat Rovira i Virgili: Utrecht, The Netherlands, 2000. [Google Scholar]
- De Hoog, G.S.; Guarro, J.; Gené, J.; Ahmed, S.; Al-Hatmi, A.M.S.; Figueras, M.J.; Vitale, R.G. Atlas of Clinical Fungi, 4th ed.; Reus: Utrecht, The Netherlands, 2019; p. 1599. [Google Scholar]
- Figueroa, M.; Jarmusch, A.K.; Raja, H.A.; El-Elimat, T.; Kavanaugh, J.S.; Horswill, A.R.; Cooks, R.G.; Cech, N.B.; Oberlies, N.H. Polyhydroxyanthraquinones as quorum sensing inhibitors from the guttates of Penicillium restrictum and their analysis by desorption electrospray ionization mass spectrometry. J. Nat. Prod. 2014, 77, 1351–1358. [Google Scholar] [CrossRef] [Green Version]
- Raja, H.A.; Kaur, A.; El-Elimat, T.; Figueroa, M.; Kumar, R.; Deep, G.; Agarwal, R.; Faeth, S.H.; Cech, N.B.; Oberlies, N.H. Phylogenetic and chemical diversity of fungal endophytes isolated from Silybum marianum (L.) Gaertn. (milk thistle). Mycology 2015, 6, 8–27. [Google Scholar] [CrossRef]
- Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Hong, S.B.; Klaassen, C.H.; Perrone, G.; Seifert, K.A.; Varga, J.; Yaguchi, T.; Samson, R.A. Identification and nomenclature of the genus Penicillium. Stud. Mycol. 2014, 78, 343–371. [Google Scholar] [CrossRef] [Green Version]
- Samson, R.A.; Houbraken, J.; Thrane, U.; Frisvad, J.C.; Andersen, B. Food and Indoor Fungi. In CBS Laboratory Manual Series; CBS-KNAW Fungal Biodiversity Centre Utrecht: Utrecht, The Netherlands, 2010; p. 310. [Google Scholar]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.E.; Crane, C.; Barrett, S.; Cano-Lira, J.F.; Le Roux, J.J.; Thangavel, R.; Guarro, J.; et al. Fungal Planet description sheets: 469–557. Persoonia 2016, 37, 218–403. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.; Barber, P.A.; Alvarado, P.; Barnes, C.W.; Buchanan, P.K.; Heykoop, M.; Moreno, G.; et al. Fungal Planet description sheets: 558–624. Persoonia 2017, 38, 240–384. [Google Scholar] [CrossRef] [PubMed]
- Kozakiewicz, Z. Aspergillus species on stored products. Mycol. Pap. 1989, 161, 1–188. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Pagès, H.; Aboyoun, P.; Gentleman, R.; DebRoy, S. Biostrings: Efficient manipulation of biological strings. In R Package Version 2.58.0; R Foundation for Statistical Computing: Vienna, Austria, version 2.58.50.
- Jukes, T.H.; Cantor, C.R. Evolution of protein molecules. In Mammalian Protein Metabolism; Munro, H.N., Ed.; Academic Press: New York, NY, USA, 1969; pp. 21–132. [Google Scholar]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Gao, F.; Jakovlic, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*. Phylogenetic analysis using parsimony (*and other methods). Version 4; Sinauer Associates: Sunderland, UK, 2002. [Google Scholar]
- El-Elimat, T.; Figueroa, M.; Ehrmann, B.M.; Cech, N.B.; Pearce, C.J.; Oberlies, N.H. High-resolution MS, MS/MS, and UV database of fungal secondary metabolites as a dereplication protocol for bioactive natural products. J. Nat. Prod. 2013, 76, 1709–1716. [Google Scholar] [CrossRef] [Green Version]
- Sulyok, M.; Stadler, D.; Steiner, D.; Krska, R. Validation of an LC-MS/MS-based dilute-and-shoot approach for the quantification of > 500 mycotoxins and other secondary metabolites in food crops: Challenges and solutions. Anal. Bioanal. Chem. 2020, 412, 2607–2620. [Google Scholar] [CrossRef] [Green Version]
- Sulyok, M. The European Commission. Guidance document on identification of mycotoxins in food and feed. Off. J. Eur. Union. 2017, 1–4, 1–4. [Google Scholar]
- Paguigan, N.D.; El-Elimat, T.; Kao, D.; Raja, H.A.; Pearce, C.J.; Oberlies, N.H. Enhanced dereplication of fungal cultures via use of mass defect filtering. J. Antibiot. 2017, 70, 553–561. [Google Scholar] [CrossRef] [Green Version]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metab. Off. J. Metab. Soc. 2007, 3, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Harrington, T.C. Cycloheximide sensitivity as a taxonomic character in Ceratocystis. Mycologia 1981, 73, 1123–1129. [Google Scholar] [CrossRef]
- Seifert, K.A.; Giuseppin, S. Cycloheximide tolerance as a taxonomic character in Penicillium. In Integration of Modern Taxonomic Methods for Penicillium and Aspergillus Classification; Samson, R.A., Pitt, J.I., Eds.; Harword Publishers: Amsterdam, The Netherlands, 2000; pp. 259–263. [Google Scholar]
- Labuda, R.; Piovarciova, Z.; Tancinova, D. Sensitivity of the Alternaria infectoria species-group 1 to cycloheximide. Lett. Appl. Microbiol. 2008, 46, 673–675. [Google Scholar] [CrossRef]
- Colotelo, N. Fungal exudates. Can. J. Microbiol. 1978, 24, 1173–1181. [Google Scholar] [CrossRef]
- Wilson, J.K. An examination of the exudate and juice of certain fungi found in their native environment. Mycologia 1948, 40, 605–613. [Google Scholar] [CrossRef]
- Jennings, D.H. The role of droplets in helping to maintain a constant growth rate of aerial hyphae. Mycol. Res. 1991, 95, 883–884. [Google Scholar] [CrossRef]
- Hutwimmer, S.; Wang, H.; Strasser, H.; Burgstaller, W. Formation of exudate droplets by Metarhizium anisopliae and the presence of destruxins. Mycologia 2010, 102, 1–10. [Google Scholar] [CrossRef] [PubMed]
- McPhee, W.J.; Colotelo, N. Fungal exudates. I. Characteristics of hyphal exudates in Fusarium culmorum. Can. J. Bot. 1977, 55, 358–365. [Google Scholar] [CrossRef]
- Koulman, A.; Lane, G.A.; Christensen, M.J.; Fraser, K.; Tapper, B.A. Peramine and other fungal alkaloids are exuded in the guttation fluid of endophyte-infected grasses. Phytochemistry 2007, 68, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sena Filho, J.G.; Hoover, A.R.; King, J.B.; Ellis, T.K.; Powell, D.R.; Cichewicz, R.H. Chemical epigenetics alters the secondary metabolite composition of guttate excreted by an atlantic-forest-soil-derived Penicillium citreonigrum. J. Nat. Prod. 2010, 73, 942–948. [Google Scholar] [CrossRef] [Green Version]
- Sica, V.P.; Figueroa, M.; Raja, H.A.; El-Elimat, T.; Darveaux, B.A.; Pearce, C.J.; Oberlies, N.H. Optimizing production and evaluating biosynthesis in situ of a herbicidal compound, mevalocidin, from Coniolariella sp. J. Ind. Microbiol. Biotechnol. 2016, 43, 1149–1157. [Google Scholar] [CrossRef] [Green Version]
- Daly, S.M.; Elmore, B.O.; Kavanaugh, J.S.; Triplett, K.D.; Figueroa, M.; Raja, H.A.; El-Elimat, T.; Crosby, H.A.; Femling, J.K.; Cech, N.B.; et al. omega-Hydroxyemodin limits Staphylococcus aureus quorum sensing-mediated pathogenesis and inflammation. Antimicrob. Agents Chemother. 2015, 59, 2223–2235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graf, T.N.; Kao, D.; Rivera-Chavez, J.; Gallagher, J.M.; Raja, H.A.; Oberlies, N.H. Drug leads from endophytic fungi: Lessons learned via scaled production. Planta Med. 2020, 86, 988–996. [Google Scholar] [CrossRef] [PubMed]
- Gessler, N.N.; Egorova, A.S.; Belozerskaya, T.A. Fungal anthraquinones. Appl. Biochem. Microbiol. 2013, 49, 85–99. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Strain Numbers | Origin | GenBank Accession Numbers | |||
|---|---|---|---|---|---|---|
| ITS | BenA | CaM | RPB2 | |||
| P. krskae | BiMM-F280 a | Lab contaminant, Austria; ex-type | MW794123 | MW774594 | MW774595 | MW774593 |
| P. silybi | G85 | Asymptomatic plant (Silybum marianum), USA; ex-type | KF367458 | MW774592 | MW774591 | AB860248 |
| P. arabicum | CBS 414.69 | Soil, Syria; ex-type | KC411758 | KP016750 | KP016770 | KP064574 |
| P. chalabudae | CBS 219.66 | Soil, Ukraine; ex-type | KP016811 | KP016748 | KP016767 | KP064572 |
| P. cinereoatrum | CBS 222.66 | Soil, Ukraine; ex-type | KC411700 | KJ834442 | KP125335 | JN406608 |
| P. corylophilum | CBS 312.48 | Unknown; ex-type | AF033450 | JX141042 | KP016780 | KP064631 |
| P. heteromorphum | CBS 226.89 | Soil, China; ex-type | KC411702 | KJ834455 | KP016786 | JN406605 |
| P. katangense | CBS 247.67 | Soil, DR Congo; ex-type | AF033458 | KP016757 | KP016788 | KP064646 |
| P. kurssanovii | CBS 625.67 | Soil, Ukraine; ex-type | EF422849 | KP016758 | KP016789 | KP064647 |
| P. meridianum | CBS 314.67 | Soil, South Africa; ex-type | AF033451 | KJ834472 | KP016794 | JN406576 |
| P. philippinense | CBS 623.72 | Soil, Philippines; ex-type | KC411770 | KJ834482 | KP016799 | JN406543 |
| P. restrictum | CBS 367.48 | Soil, Honduras; ex-type | AF033457 | KJ834486 | KP016803 | JN121506 |
| P. cf. restrictum | CBS 139139 | Soil, South Africa; fynbos | JX140930 | JX141055 | JX157419 | KP064607 |
| P. cf. restrictum | CBS 139149 | Soil, South Africa; fynbos | KP016819 | JX141060 | JX157467 | KP064609 |
| P. cf. restrictum | CBS 139150 | Soil, South Africa; fynbos | KP016816 | JX141061 | JX157468 | KP064610 |
| P. cf. restrictum | CBS 139153 | Soil, South Africa; fynbos | KP016818 | JX141064 | JX157474 | KP064612 |
| P. cf. restrictum | CBS 139170 | Soil, South Africa; fynbos | - | JX141056 | JX157420 | KP064644 |
| Fungus | Growth on CYA a | Growth on MEA-CX250 b | Growth and Acid Production on CREA c | Red Exudate or Pigment Present | Conidial Morphology | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 15 °C | 25 °C | 30 °C | 37 °C | Average Size in µm | Surface Ornamentation | |||||
| LM | SEM | |||||||||
| P. krskae | 0 | 18–20 | 24–26 | 8–10 | 5 (15) | 12–14 / + | Yes | 2.1 | sm-r-ro | verruc |
| P. silybi | 5–7 | 26–28 | 25–27 | 3–5 | 10 (25) | 20–22 /++ | Yes ** | 2.0 | ro-str | lob-retic |
| P. chalabudae | 5–7 | 13–16 | 18–20 | 10–12 | 0 * | 8–10 / ++ | No | 2.1 | ro | tuber |
| P. kurssanovii | 3–5 | 12–15 | 15–18 | 0 | 0 * | 8–10 / - (±) | No | 2.3 | ro | tuber |
| P. restrictum | 2–4 | 18–20 | 22–24 | 8–10 | 0 * | 9–11 /- (±) | No | 2.4 | ro-ech | acul-ech |
| Fungus | MEA Mm * | MEA-CX in mm (% Inhibition) | ||
|---|---|---|---|---|
| 100 | 250 | 500 | ||
| P. krskae | 20–30 | 8 (60)–20 (33) | 5 (75)–15 (50) | 0 (100)–3 (90) |
| P. silybi | 28–48 | 18 (36)–35 (27) | 12 (57)–25 (48) | 6 (78)–18 (62) |
| P. chalabudae | 18–40 | 2 (89)–18 (55) | 0 (100)–0 (100) | 0 (100)–0 (100) |
| P. kurssanovii | 22–42 | 0 (100)–0 (100) | 0 (100)–0 (100) | 0 (100)–0 (100) |
| P. restricum | 18–30 | 0 (100)–5 (83) | 0 (100)–0 (100) | 0 (100)–0 (100) |
| Compound Name a | Molecular Formula | P. krskae | P. silybi | ||
|---|---|---|---|---|---|
| PDA | MEA | PDA | MEA | ||
| 2-hydroxyemodic acid | C15H8O8 | − | − | + | − |
| (+)-2’S-Isorhodoptilometrin | C17H14O6 | + | + | + | + |
| 1’-Hydroxy-2’-ketoisorhodoptilometrin | C17H12O7 | + | − | + | + |
| 7-Chloro-1’-hydroxyisorhodoptilometrin | C17H13O7Cl | + b | + | − | + |
| 1’-Hydroxyisorhodoptilometrin | C17H14O7 | + b | + | + | + |
| 2-Chloroemodic acid | C15H7ClO7 | + b | + | + | + |
| 2-Chloro-isorhodoptilometrin * | C17H13O6Cl | + b | + | − | + |
| 2-Chloro-desmethyl dermoquinone * | C17H11O6Cl | + b | + | − | + |
| 7-Chlorocitreosein | C15H9ClO6 | + b | + | − | + |
| O-Demethyldermoquinone | C17H12O6 | + | + | + | + |
| Emodic acid | C15H8O7 | + | + | + | + |
| Emodin | C15H10O5 | + | − | + | + |
| ω-Hydroxyemodin (citreorosein) | C15H10O6 | + b | + | + | + |
| Compound a | P. krsk | P. silb | P. chal | P. kurs | P. rest |
|---|---|---|---|---|---|
| 15-Hydroxyculmorin | - | - | - | - | 1.10 * |
| 7-Hydroxypestalotin | 68.39 | 8.69 | 0.95 | - | - |
| Chlorocitreorosein | 0.54 | 355.20 | 0.12 | 0.19 | 0.08 |
| Citreorosein | 0.49 | 751.35 | 0.48 | 1.11 | 0.62 |
| Emodin | - | 36.02 | - | 0.01 | 0.03 |
| Endocrocin | - | 93.71 * | - | - | - |
| Hydroxysydonic acid | - | - | - | - | 5.83 * |
| Iso-rhodoptilometrin | 0.16 | 264.90 | 0.20 | 0.26 | 0.22 |
| NP1243 | 1.67 | 5.66 | 0.28 | 0.62 | 0.18 |
| Oxyskyrin | - | 0.05* | - | - | - |
| Paxilline | 4.31 * | - | - | - | - |
| Pestalotin | 1.26 | 0.15 | 0.26 | - | - |
| Skyrin | - | 0.15 * | - | - | - |
| Tryptophol | - | - | 1.52 * | - | - |
| Methylorsellinic acid | - | - | - | - | 0.30 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Labuda, R.; Bacher, M.; Rosenau, T.; Gasparotto, E.; Gratzl, H.; Doppler, M.; Sulyok, M.; Kubátová, A.; Berger, H.; Cank, K.; et al. Polyphasic Approach Utilized for the Identification of Two New Toxigenic Members of Penicillium Section Exilicaulis, P. krskae and P. silybi spp. nov. J. Fungi 2021, 7, 557. https://doi.org/10.3390/jof7070557
Labuda R, Bacher M, Rosenau T, Gasparotto E, Gratzl H, Doppler M, Sulyok M, Kubátová A, Berger H, Cank K, et al. Polyphasic Approach Utilized for the Identification of Two New Toxigenic Members of Penicillium Section Exilicaulis, P. krskae and P. silybi spp. nov. Journal of Fungi. 2021; 7(7):557. https://doi.org/10.3390/jof7070557
Chicago/Turabian StyleLabuda, Roman, Markus Bacher, Thomas Rosenau, Erika Gasparotto, Hannes Gratzl, Maria Doppler, Michael Sulyok, Alena Kubátová, Harald Berger, Kristof Cank, and et al. 2021. "Polyphasic Approach Utilized for the Identification of Two New Toxigenic Members of Penicillium Section Exilicaulis, P. krskae and P. silybi spp. nov." Journal of Fungi 7, no. 7: 557. https://doi.org/10.3390/jof7070557