
Bioinformatics and Transcriptome Analysis of CFEM Proteins in Fusarium graminearum


Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture of Fungal Strain
2.2. Plant Growth Conditions and Inoculation
2.3. RNA Extraction and Microarray Hybridization
2.4. Bioinformatic Analysis of Common in Fungal Extracellular Membrane (CFEM)-Containing Proteins in F. Graminearum
2.4.1. Identification of CFEM-Containing Proteins in F. Graminearum Genome
2.4.2. Signal Peptide, Transmembrane Domain, and Subcellular Localization Prediction
2.4.3. Phylogenetic Analysis and Multiple Sequences Alignment
2.4.4. Domain Analysis of FgCFEM Proteins
2.4.5. Glycosylphosphatidylinositol (GPI) Modification Site Prediction
2.4.6. Analysis of Candidate Effectors of CFEM-Containing Proteins
3. Results
3.1. Bioinformatics Identification of CFEM Proteins in F. Graminearum
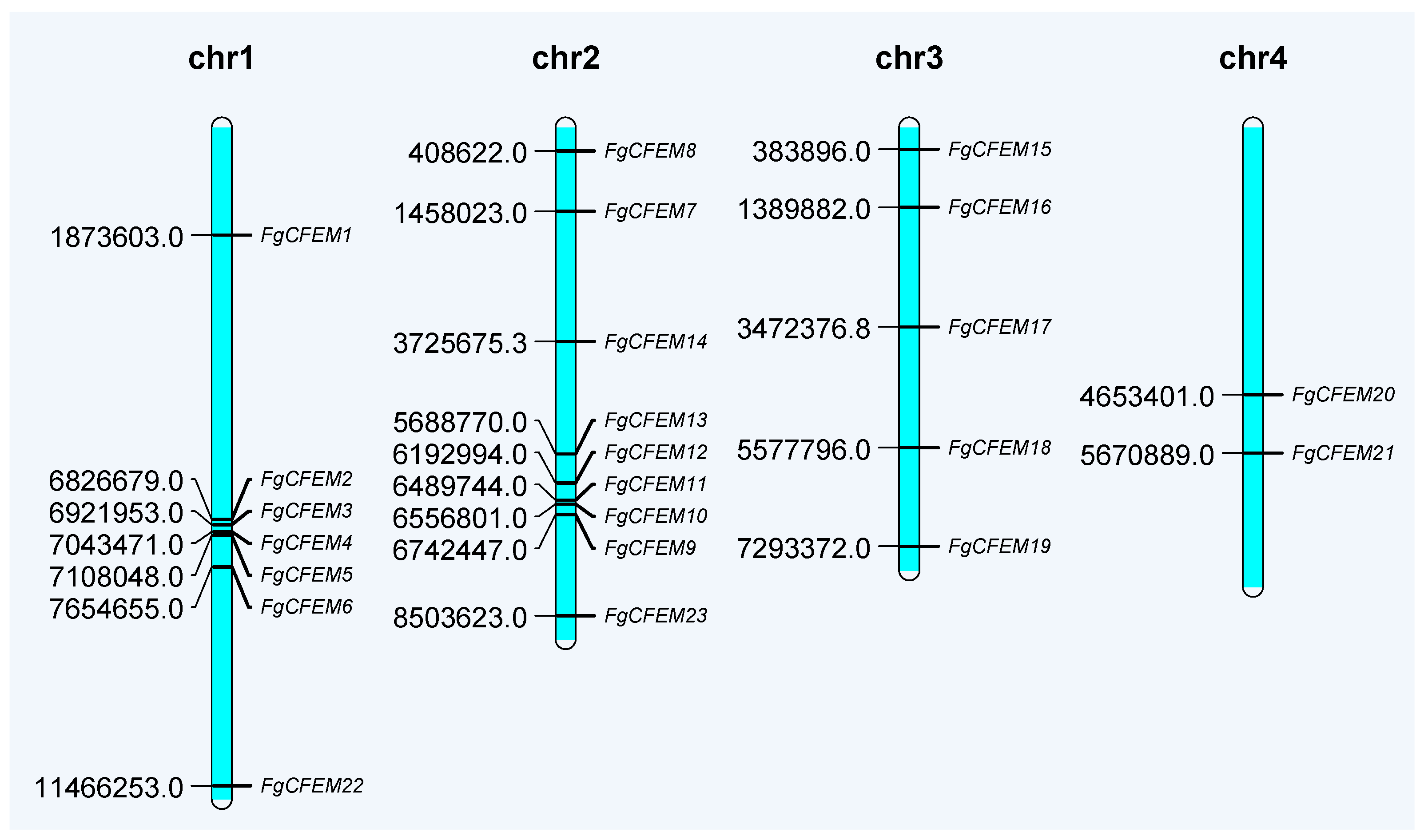
3.2. Chromosomal Distribution of CFEM-Containing Genes
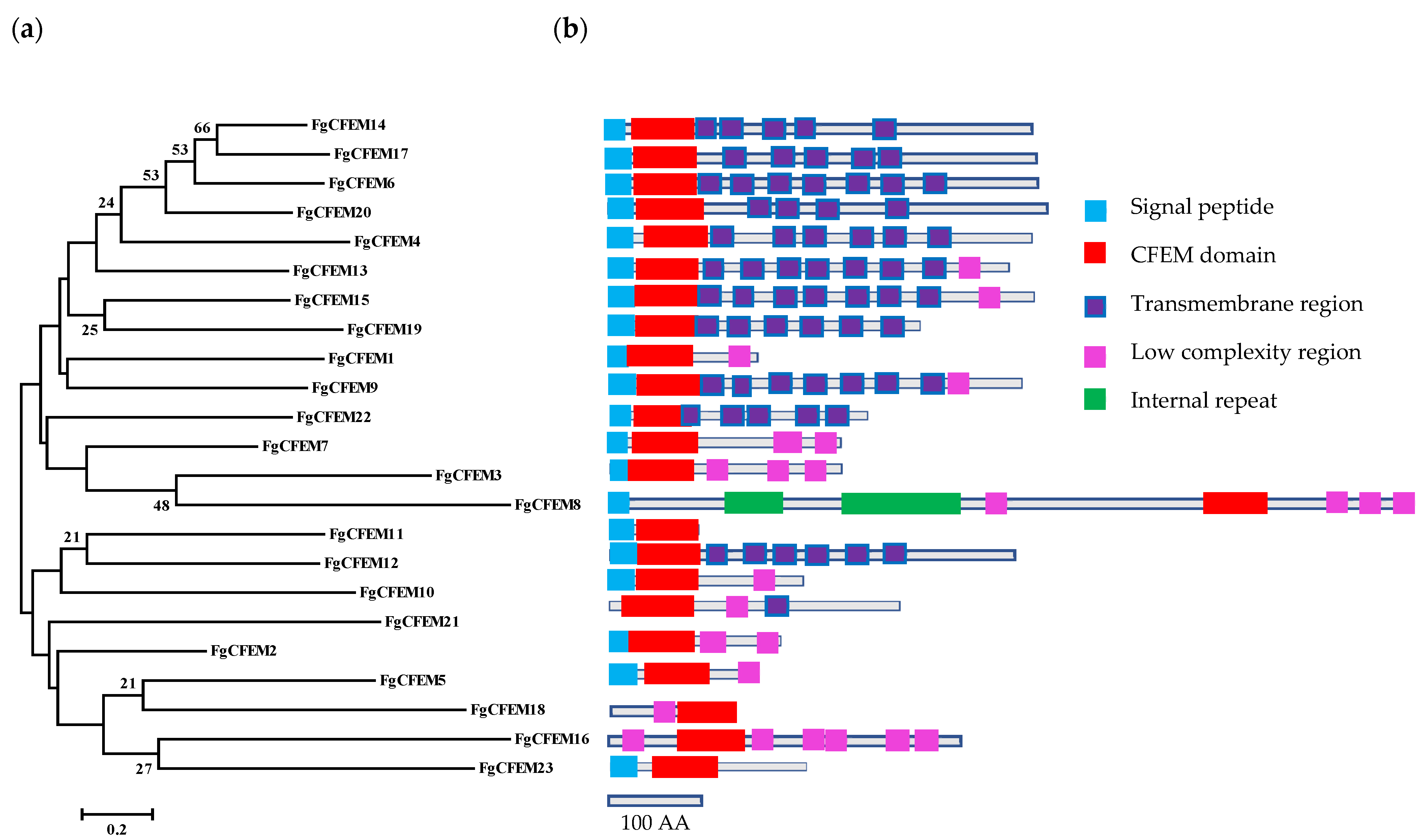
3.3. Feature Characterization of CFEM-Containing Proteins
3.4. Identification of Potential CFEM Effectors in F. Graminearum
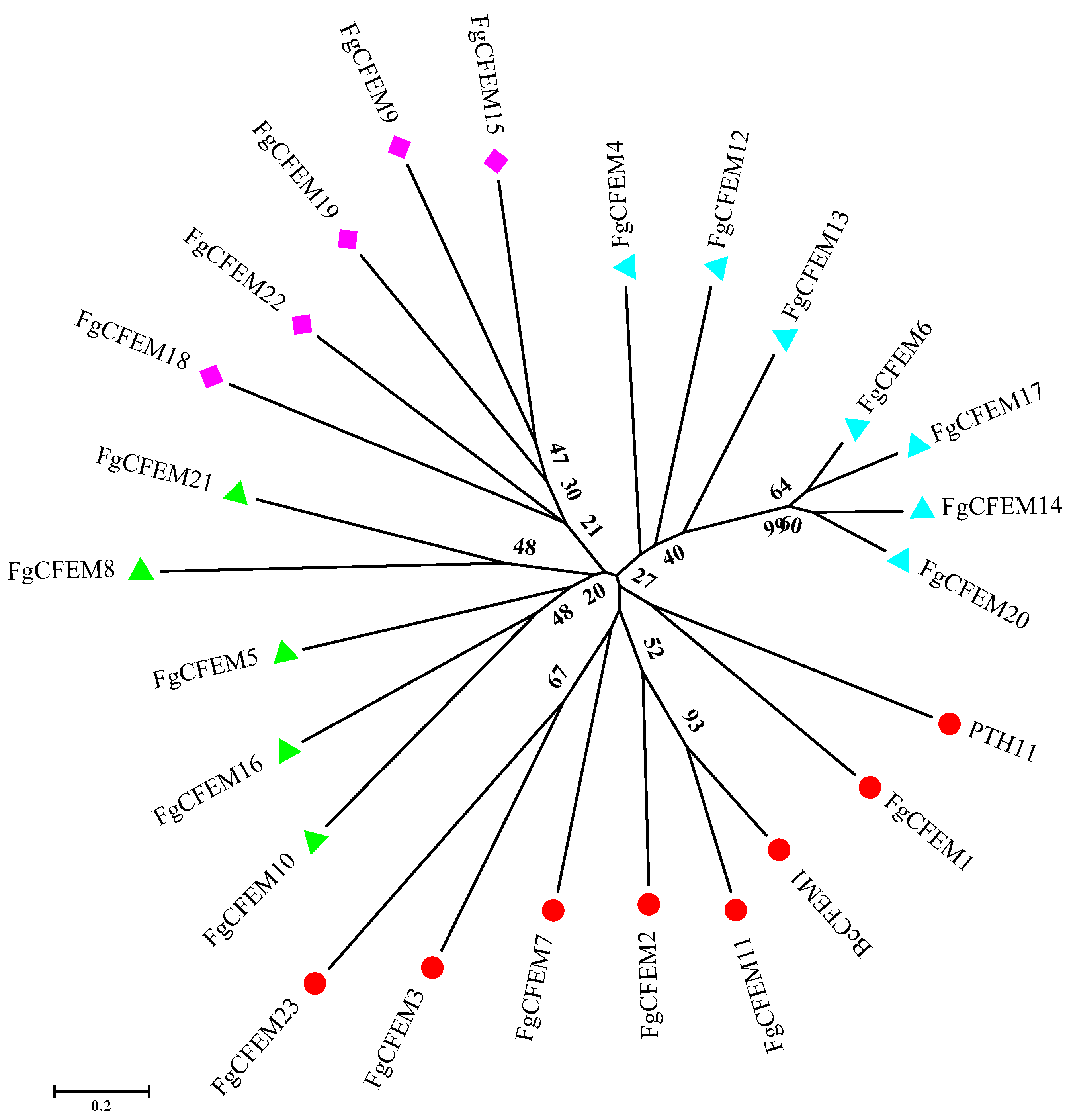
3.5. Phylogenetic Analysis of CFEM Proteins from F. Graminearum
3.6. Conserved Motif Analysis
3.7. The Transcriptometrics Analysis of CFEM Genes in Planta
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Garmendiaa, G.; Pattarinoa, L.; Negrína, C.; Martínez-Silveiraa, A.; Pereyrab, S.; Wardc, T.J.; Vero, S. Species composition, toxigenic potential and aggressiveness of Fusarium isolates causing Head Blight of barley in Uruguay. Food Microbiol. 2018, 76, 426–433. [Google Scholar] [CrossRef]
- Starkey, D.E.; Ward, T.J.; Aoki, T.; Gale, L.R.; Corby Kistler, H.; Geiser, D.M.; Suga, H.; Toth, B.; Varga, J.; O’Donnell, K. Global molecular surveillance reveals novel Fusarium head blight species and trichothecene toxin diversity. Fungal Genet. Biol. 2007, 44, 1191–1204. [Google Scholar] [CrossRef]
- Trail, F. For blighted waves of grain: Fusarium graminearum in the postgenomics era. Plant Physiol. 2009, 149, 103–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Pietro, A.D.; Spana, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]
- Desjardins, A.E.; Proctor, R.H.; Bai, G.; McCormick, S.P.; Shaner, G.; Buechely, G.; Hohn, T.M. Reduced virulence of trichothecene-nonproducing mutants of Gibberella zeae in wheat field tests. Mol. Plant Microbe Interact. 1996, 9, 775–781. [Google Scholar] [CrossRef]
- Jansen, C.; von Wettstein, D.; Schafer, W.; Kogel, K.H.; Felk, A.; Maier, F.J. Infection patterns in barley and wheat spikes inoculated with wild-type and trichodiene synthase gene disrupted Fusarium graminearum. Pro. Natl. Acad. Sci. USA 2005, 102, 16892–16897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proctor, R.H.; Hohn, T.M.; McCormick, S.P. Reduced virulence of Gibberella zeae caused by disruption of a trichothecene toxin biosynthetic gene. Mol. Plant Microbe Interact. 1995, 8, 593–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.W.; Sun, S.; Ge, W.Y.; Zhao, L.F.; Hou, B.Q.; Wang, K.; Lyu, Z.F.; Chen, L.Y.; Xu, S.S.; Guo, J.; et al. Horizontal gene transfer of Fhb7 from fungus underlies Fusarium head blight resistance in wheat. Science 2020, 368, 844. [Google Scholar] [CrossRef]
- Narayanan, S.P.; Lung, S.C.; Liao, P.; Lo, C.; Chye, M.L. The overexpression of OsACBP5 protects transgenic rice against necrotrophic, hemibiotrophic and biotrophic pathogens. Sci. Rep. 2020, 10, 14918. [Google Scholar] [CrossRef]
- Lu, S.; Edwards, M.C. Genome-wide analysis of small secreted cysteine-rich proteins identifies candidate effector proteins potentially involved in Fusarium graminearum-wheat interactions. Phytopathology 2016, 106, 166–176. [Google Scholar] [CrossRef] [Green Version]
- Appella, E.; Weber, I.T.; Blasi, F. Structure and function of epidermal growth factor-like regions in proteins. FEBS Lett. 1988, 231, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, R.D.; Kelkar, H.S.; Dean, R.A. An eight-cysteine-containing CFEM domain unique to a group of fungal membrane proteins. Trends Biochem. Sci. 2003, 28, 118–121. [Google Scholar] [CrossRef]
- Zhang, Z.N.; Wu, Q.Y.; Zhang, G.Z.; Zhu, Y.Y.; Murphy, R.W.; Liu, Z.; Zou, C.G. Systematic analyses reveal uniqueness and origin of the CFEM domain in fungi. Sci. Rep. 2015, 5, 13032. [Google Scholar] [CrossRef] [Green Version]
- Pérez, A.; Pedrós, B.; Murgui, A.; Casanova, M.; López-Ribot, J.L.; Martínez, J.P. Biofilm formation by Candida albicans mutants for genes coding fungal proteins exhibiting the eight-cysteine-containing CFEM domain. FEMS Yeast Res. 2006, 6, 1074–1084. [Google Scholar] [CrossRef] [Green Version]
- Pérez, A.; Ramage, G.; Blanes, R.; Murgui, A.; Casanova, M.; Martínez, J.P. Some biological features of Candida albicans mutants for genes coding fungal proteins containing the CFEM domain. FEMS Yeast Res. 2011, 11, 273–284. [Google Scholar] [CrossRef] [Green Version]
- Vaknin, Y.; Shadkchan, Y.; Levdansky, E.; Morozow, M.; Romano, J.; Osherov, N. The three Aspergillus fumigatus CFEM-domain GPI-anchored proteins (CfmA-C) affect cell-well stability but do not play a role in fungal virulence. Fungal Genet. Biol. 2014, 63, 55–64. [Google Scholar] [CrossRef]
- Kou, Y.; Tan, Y.H.; Ramanujam, R.; Naqvi, N. Structure-function analyses of the Pth11 receptor reveal an important role for CFEM motif and redox regulation in rice blast. New Phytol. 2017, 214, 330–342. [Google Scholar] [CrossRef]
- Ling, J.; Zeng, F.; Gao, Y.X.; Zhang, J.X.; Chen, G.H.; Mao, Z.C.; Yang, Y.H.; Xie, B.Y. Identification of a class of CFEM proteins containing a new conserved motif in Fusarium oxyporum. Physiol. Mol. Plant Pathol. 2015, 89, 41–48. [Google Scholar] [CrossRef]
- Zhao, S.; Shang, X.; Bi, W.; Yu, X.; Liu, D.; Kang, Z.; Wang, X.; Wang, X. Genome-wide identification of effector candidates with conserved motifs from the wheat leaf rust fungus Puccinia triticina. Front. Microbiol. 2020, 11, 1188. [Google Scholar] [CrossRef]
- Ding, C.; Vidanes, G.M.; Maguire, S.L.; Guida, A.; Synnott, J.M.; Andes, D.R.; Butler, G. Conserved and divergent roles of Bcr1 and CFEM proteins in Candida parapsilosis and Candida albicans. PLoS ONE 2011, 6, e28151. [Google Scholar] [CrossRef] [Green Version]
- Weissman, Z.; Kornitzer, D. A family of Candida cell surface haem-binding proteins involved in haemin and haemoglobin-iron utilization. Mol. Microbiol. 2004, 53, 1209–1220. [Google Scholar] [CrossRef]
- Weissman, Z.; Shemer, R.; Conibear, E.; Kornitzer, D. An endocytic mechanism for haemoglobin-iron acquisition in Candida albicans. Mol. Microbiol. 2008, 69, 201–217. [Google Scholar] [CrossRef]
- Choi, W.; Dean, R.A. The adenylate cyclase gene MAC1 of Magnaporthe grisea controls appressorium formation and other aspects of growth and development. Plant Cell 1997, 9, 1973–1983. [Google Scholar]
- DeZwaan, T.M.; Carroll, A.M.; Valent, B.; Sweigard, J.A. Magnaporthe grisea Pth11p Is a Novel Plasma Membrane Protein That Mediates Appressorium Differentiation in Response to Inductive Substrate Cues. Plant Cell 1999, 11, 2013–2030. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.J.; Wei, W.; Wu, Y.Y.; Zhou, Y.; Peng, F.; Zhang, S.P.; Chen, P.; Xu, X.W. BcCFEM1, a CFEM Domain-Containing Protein with Putative GPI-Anchored Site, Is Involved in Pathogenicity, Conidial Production, and Stress Tolerance in Botrytis cinerea. Front. Microbiol. 2017, 8, 1807. [Google Scholar] [CrossRef]
- Arya, G.C.; Srivastava, D.A.; Pandaranayaka, E.P.J.; Manasherova, E.; Prusky, D.B.; Elad, Y.; Frenkel, O.; Dvir, H.; Harel, A. Characterization of the role of a non-GPCR membrane-bound CFEM protein in the pathogenicity and germination of Botrytis cinerea. Microorganisms 2020, 8, 1043. [Google Scholar] [CrossRef]
- Hao, G.; McMormick, S.; Usgaard, T.; Tiley, H.; Vaughan, M.M. Characterization of three Fusarium graminearum effectors and their roles during fusarium head blight. Front. Plant Sci. 2020, 11, 579553. [Google Scholar] [CrossRef]
- King, R.; Urban, M.; Hammond-Kosack, M.C.U.; Hassani-Pak, K.; Hammond-Kosack, K.E. The completed genome sequence of the pathogenic ascomycete fungus Fusarium graminearum. BMC Genom. 2015, 16, 544. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Jia, L.; Zhang, Y.; Jiang, G.; Li, X.; Zhang, D.; Tang, W. In planta stage-specific fungal gene profiling elucidates the molecular strategies of Fusarium graminearum growing inside wheat coleoptiles. Plant Cell 2012, 24, 5159–5176. [Google Scholar] [CrossRef] [Green Version]
- Wu, A.B.; Li, H.P.; Zhao, C.S.; Liao, Y.C. Comparative pathogenicity of Fusarium graminearum isolates from China revealed by wheat coleoptile and floret inoculations. Mycopathologia 2005, 160, 75–83. [Google Scholar] [CrossRef]
- Zhang, Y.; He, J.; Jia, L.; Yuan, T.; Zhang, D.; Guo, Y.; Wang, Y.; Tang, W. Cellular tracking and gene profiling of Fusarium graminearum during maize stalk rot disease development elucidates its strategies in confronting phosphorus limitation in the host apoplast. PLoS Pathog. 2016, 12, e1005485. [Google Scholar] [CrossRef] [Green Version]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Williams, N.; Misleh, C.; Li, W.W. MEME: Discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006, 34, W369e73. [Google Scholar] [CrossRef]
- Eisenhaber, B.; Schneider, G.; Wildpaner, M.; Eisenhaber, F. A sensitive predictor for potential GPI lipid modification sites in fungal protein sequences and its application to genome-wide studies for Aspergillus nidulans, Candida albicans, Neurospora crassa, Saccharomyces cerevisiae, and Schizosaccharomyces pombe. J. Mol. Biol. 2004, 337, 243–253. [Google Scholar] [CrossRef]
- Brown, N.A.; Antoniw, J.; Hammond-Kosack, K.E. The predicted secretome of the plant pathogenic fungus Fusarium graminearum: A refined comparative analysis. PLoS ONE 2012, 7, e33731. [Google Scholar] [CrossRef] [Green Version]
- Sperschneider, J.; Gardiner, D.M.; Dodds, P.N.; Tini, F.; Covarelli, L.; Singh, K.B.; Manners, J.M.; Taylor, J.M. EffectorP: Predicting fungal effector proteins from secretomes using machine learning. New Phytol. 2016, 210, 743–761. [Google Scholar] [CrossRef] [Green Version]
- Krüger, A.T.; Engel, J.; Buettner, F.F.; Routier, F.H. Aspergillus fumigatus Cap59-like protein A is involved in α1,3-mannosylation of GPI-anchors. Glycobiology 2016, 26, 30–38. [Google Scholar]
- Lo Presti, L.; Lanver, D.; Schweizer, G.; Tanaka, S.; Liang, L.; Tollot, M.; Zuccaro, A.; Reissmann, S.; Kahmann, R. Fungal effectors and plant susceptibility. Annu. Rev. Plant Biol. 2015, 66, 513–545. [Google Scholar] [CrossRef]
- Nasser, L.; Weissman, Z.; Pinsky, M.; Amartely, H.; Dvir, H.; Kornitzer, D. Structural basis of haem-iron acquisition by fungal pathogens. Nat. Microbiol. 2016, 1, 16156. [Google Scholar] [CrossRef]
- Gong, A.D.; Jing, Z.Y.; Zhang, K.; Tan, Q.Q.; Wang, G.L.; Liu, W.D. Bioinformatic analysis and functional characterization of the CFEM proteins in maize anthracnose fungus Colletotrichum graminicola. J. Integr. Agric. 2020, 19, 541–550. [Google Scholar] [CrossRef]
- Moukadiri, I.; Armero, J.; Abad, A.; Sentandreu, R.; Zueco, J. Identification of a mannoprotein present in the inner layer of the cell wall of Saccharomyces cerevisiae. J. Bacteriol. 1997, 179, 2154–2162. [Google Scholar] [CrossRef] [Green Version]
- Sabnam, N.; Roy-Barman, S. WISH, a novel CFEM GPCR is indispensable for surface sensing, asexual and pathogenic differentiation in rice blast fungus. Fungal Genet. Biol. 2017, 105, 37–51. [Google Scholar] [CrossRef]
- Yang, F.; Jensen, J.D.; Svensson, B.; Jorgensen, H.J.L.; Collinge, D.B.; Finnie, C. Secretomics identifies Fusarium graminearum proteins involved in the interaction with barley and wheat. Mol. Plant Pathol. 2012, 13, 445–453. [Google Scholar] [CrossRef]
- Nombela, C.; Gil, C.; Chaffin, W.L. Non-conventional protein secretion in yeast. Trends Microbiol. 2006, 14, 15–21. [Google Scholar] [CrossRef]
- Eisenhaber, B.; Bork, P.; Eisenhaber, F. Post-translational GPI lipid anchor modification of proteins in kingdoms of life: Analysis of protein sequence data from complete genomes. Protein Eng. Des. Sel. 2001, 14, 17–25. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Protein ID | Amino Acid (AA) | No. of Cys | Cys% in Matured Protein | Position of CFEM Domain | SP Cleavage 1 | mTP 2 | SP | Other 3 | Loc | TM 4 no. | GPI-Anchored | Effector |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FgCFEM1 | FGRAMPH1_01G01499 | 160 | 10 | 6.99% | 18–85 | 17–18 | 0 | 0.9998 | 0.0002 | S 5 | 0 | N139/G131 | Y 7 |
| FgCFEM2 | FGRAMPH1_01G05009 | 184 | 8 | 4.79% | 19–86 | 17–18 | 0 | 0.9999 | 0.0001 | S | 0 | N163/G164 | - |
| FgCFEM3 | FGRAMPH1_01G05085 | 250 | 9 | 3.90% | 17–84 | 19–20 | 0 | 0.9998 | 0.0002 | S | 0 | S220/A228 | - |
| FgCFEM4 | FGRAMPH1_01G05193 | 457 | 12 | 2.73% | 25–90 | 17–18 | 0 | 0.9992 | 0.0008 | - 6 | 6 | - | - |
| FgCFEM5 | FGRAMPH1_01G05255 | 161 | 8 | 5.80% | 35–101 | 20–21 | 0.0005 | 0.9993 | 0.0002 | S | 0 | N138/A139 | Y |
| FgCFEM6 | FGRAMPH1_01G05701 | 463 | 18 | 4.05% | 28–92 | 19–20 | 0.0001 | 0.9925 | 0.0075 | - | 7 | - | - |
| FgCFEM7 | FGRAMPH1_01G10249 | 207 | 8 | 4.17% | 17–84 | 15–16 | 0 | 0.9999 | 0.0001 | S | 0 | G176/G186 | - |
| FgCFEM8 | FGRAMPH1_01G10975 | 864 | 22 | 2.60% | 640–705 | 17–18 | 0 | 0.9996 | 0.0003 | S | 0 | - | - |
| FgCFEM9 | FGRAMPH1_01G13033 | 440 | 15 | 3.58% | 23–87 | 21–22 | 0.0003 | 0.7624 | 0.2374 | - | 8 | - | - |
| FgCFEM10 | FGRAMPH1_01G13195 | 210 | 8 | 4.23% | 27–90 | 21–22 | 0 | 0.9997 | 0.0003 | S | 0 | S186/D185 | - |
| FgCFEM11 | FGRAMPH1_01G13253 | 95 | 10 | 12.99% | 29–92 | 18–19 | 0 | 1 | 0 | S | 0 | - | Y |
| FgCFEM12 | FGRAMPH1_01G13513 | 435 | 17 | 4.10% | 26–90 | 20–21 | 0 | 0.9991 | 0.0008 | - | 6 | - | - |
| FgCFEM13 | FGRAMPH1_01G13985 | 430 | 18 | 4.38% | 25–88 | 19–20 | 0 | 0.9993 | 0.0007 | - | 7 | - | - |
| FgCFEM14 | FGRAMPH1_01G15521 | 458 | 15 | 3.42% | 26–90 | 19–20 | 0.0004 | 0.9986 | 0.001 | - | 5 | - | - |
| FgCFEM15 | FGRAMPH1_01G16401 | 458 | 9 | 2.05% | 26–90 | 20–21 | 0.0005 | 0.9338 | 0.0657 | - | 7 | - | - |
| FgCFEM16 | FGRAMPH1_01G17281 | 379 | 8 | 2.11% | 73–142 | −6 | 0.2608 | 0.0319 | 0.7073 | S | 0 | N355/S347 | - |
| FgCFEM17 | FGRAMPH1_01G18835 | 461 | 15 | 3.40% | 28–92 | 20–21 | 0.0009 | 0.9978 | 0.0013 | - | 5 | - | - |
| FgCFEM18 | FGRAMPH1_01G21361 | 129 | 7 | 5.43% | 65–125 | - | 0.0007 | 0.4664 | 0.5329 | S | 0 | - | Y |
| FgCFEM19 | FGRAMPH1_01G21947 | 334 | 9 | 2.87% | 24–88 | 20–21 | 0 | 0.9543 | 0.0457 | - | 6 | - | - |
| FgCFEM20 | FGRAMPH1_01G25789 | 472 | 18 | 3.97% | 27–96 | 19–20 | 0.0001 | 0.9956 | 0.0043 | - | 4 | - | - |
| FgCFEM21 | FGRAMPH1_01G26539 | 312 | 8 | 2.56% | 9–83 | - | 0 | 0 | 1 | - | 1 | - | - |
| FgCFEM22 | FGRAMPH1_01G08575 | 280 | 8 | 3.10% | 26–88 | 22–23 | 0 | 0.906 | 0.094 | - | 5 | - | - |
| FgCFEM23 | FGRAMPH1_01G11435 | 189 | 10 | 5.92% | 28–95 | 20–21 | 0 | 0.9994 | 0.0006 | S | 0 | N168/G169 | Y |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Wang, H.; Yang, J.; Yang, X.; Zhang, M.; Zhao, Z.; Fan, Y.; Wang, C.; Wang, J. Bioinformatics and Transcriptome Analysis of CFEM Proteins in Fusarium graminearum. J. Fungi 2021, 7, 871. https://doi.org/10.3390/jof7100871
Chen L, Wang H, Yang J, Yang X, Zhang M, Zhao Z, Fan Y, Wang C, Wang J. Bioinformatics and Transcriptome Analysis of CFEM Proteins in Fusarium graminearum. Journal of Fungi. 2021; 7(10):871. https://doi.org/10.3390/jof7100871
Chicago/Turabian StyleChen, Lingqiao, Haoyu Wang, Junhua Yang, Xianli Yang, Mengyuan Zhang, Zhihui Zhao, Yingying Fan, Cheng Wang, and Jianhua Wang. 2021. "Bioinformatics and Transcriptome Analysis of CFEM Proteins in Fusarium graminearum" Journal of Fungi 7, no. 10: 871. https://doi.org/10.3390/jof7100871