
Antimycotic Effects of 11 Essential Oil Components and Their Combinations on 13 Food Spoilage Yeasts and Molds


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Chemicals and Cultivation
2.2. Antifungal Susceptibility Tests
2.3. Synergy Tests via Checkerboard Method
2.4. Growth Kinetics
2.5. Flow Cytometric Analyses
2.6. Statistical Analysis
3. Results
3.1. Cinnamaldehyde Showed Highest MIC
3.2. Synergistic Effects of Cinnamaldehyde with EOCs
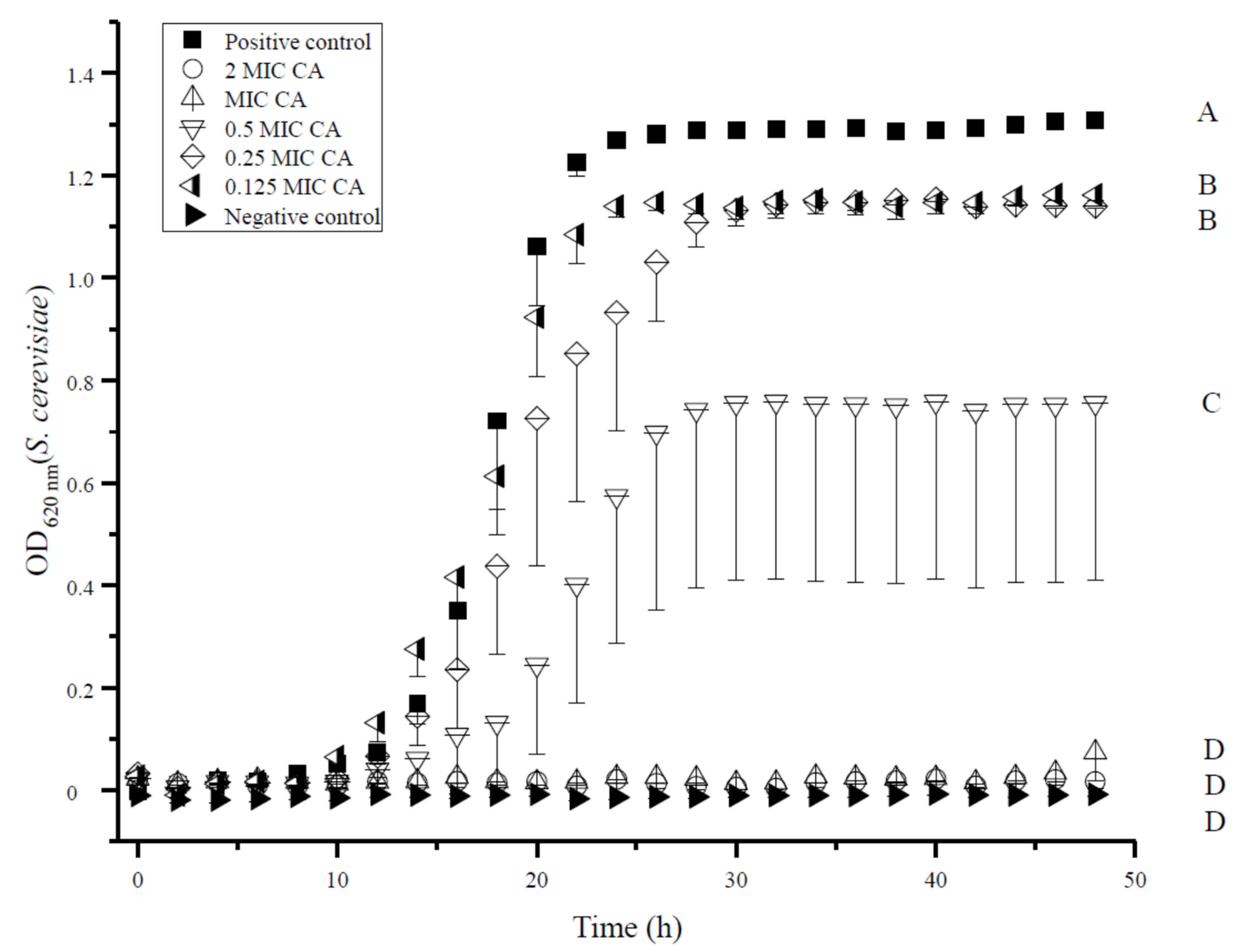
3.3. Growth Kinetics of S. cerevisiae were Impaired by CA
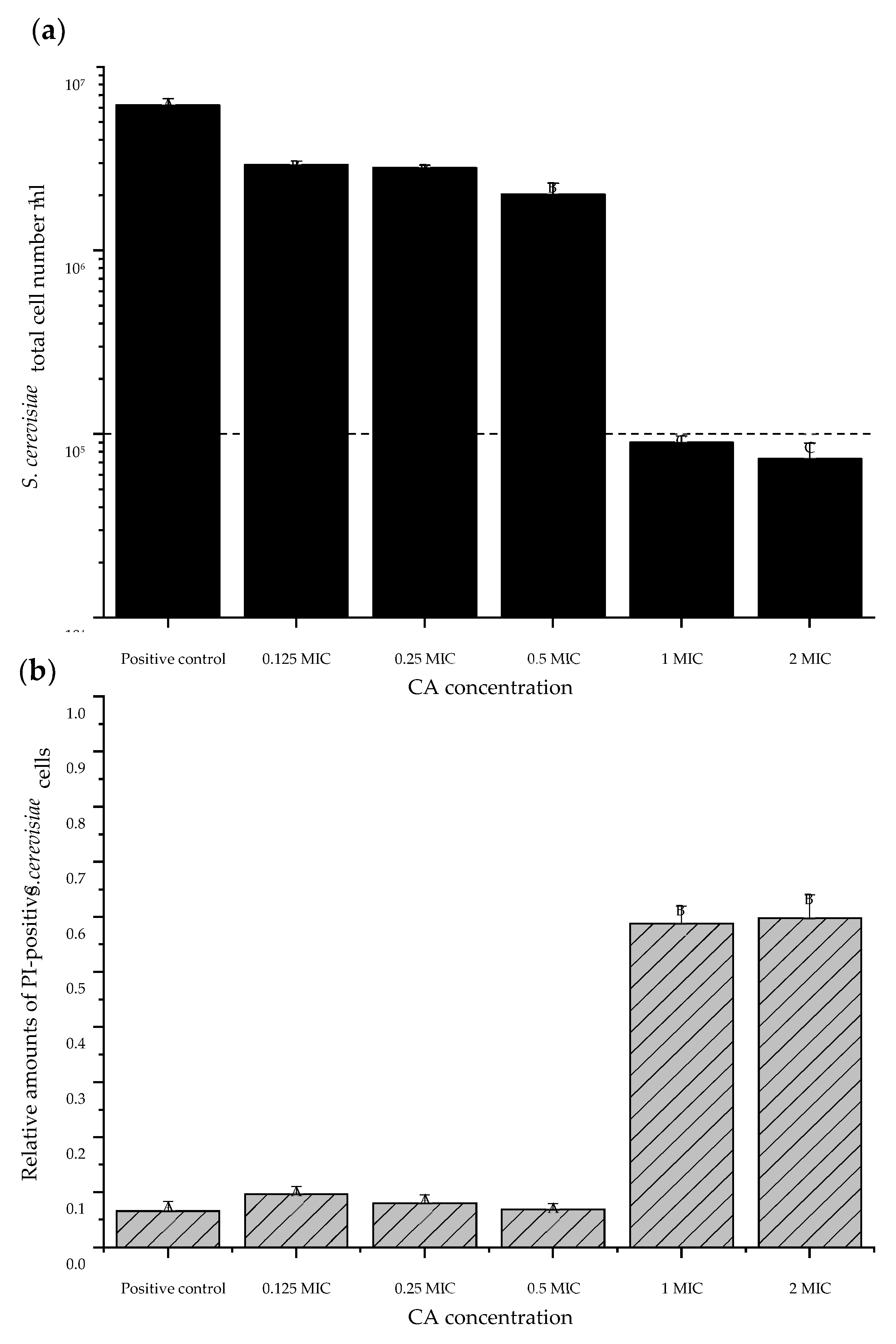
3.4. Cinnamaldehyde Showed Fungicidal Effects
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bueno, D.J.; Silva, J.O.; Oliver, G. Fungal isolation and enumeration in foods. Met. Mol. Biol. 2004, 268, 127–131. [Google Scholar] [CrossRef]
- Fung, F.; Clark, R.F. Health effects of mycotoxins: A toxicological overview. J. Toxicol. Clin. Toxicol. 2004, 42, 217–234. [Google Scholar] [CrossRef] [PubMed]
- Valle Garcia, M.; Sonnenstrahl Bregão, A.; Parussolo, G.; Olivier Bernardi, A.; Stefanello, A.; Venturini Copetti, M. Incidence of spoilage fungi in the air of bakeries with different hygienic status. Int. J. Food Microbiol. 2019, 290, 254–261. [Google Scholar] [CrossRef]
- Tian, J.; Wang, Y.; Zeng, H.; Li, Z.; Zhang, P.; Tessema, A.; Peng, X. Efficacy and possible mechanisms of perillaldehyde in control of Aspergillus niger causing grape decay. Int. J. Food Microbiol. 2015, 202, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Omidbeygi, M.; Barzegar, M.; Hamidi, Z.; Naghdibadi, H. Antifungal activity of thyme, summer savory and clove essential oils against Aspergillus flavus in liquid medium and tomato paste. Food Control 2007, 18, 1518–1523. [Google Scholar] [CrossRef]
- de Souza, E.L.; da Cruz Almeida, E.T.; de Sousa Guedes, J.P. The potential of the incorporation of essential oils and their individual constituents to improve microbial safety in juices: A review. Compr. Rev. Food Sci. Food Saf. 2016, 15, 753–772. [Google Scholar] [CrossRef] [Green Version]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils--a review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Bishop, C.D. Antiviral activity of the essential oil of Melaleuca alternifolia (Maiden amp; Betche) cheel (tea tree) against tobacco mosaic virus. J. Essent. Oil Res. 1995, 7, 641–644. [Google Scholar] [CrossRef]
- Juglal, S.; Govinden, R.; Odhav, B. Spice oils for the control of co-occurring mycotoxin-producing fungi. J. Food Prot. 2002, 65, 683–687. [Google Scholar] [CrossRef] [PubMed]
- Ultee, A.; Smid, E.J. Influence of carvacrol on growth and toxin production by Bacillus cereus. Int. J. Food Microbiol. 2001, 64, 373–378. [Google Scholar] [CrossRef]
- Pessoa, L.M.; Morais, S.M.; Bevilaqua, C.M.L.; Luciano, J.H.S. Anthelmintic activity of essential oil of Ocimum gratissimum Linn. and eugenol against Haemonchus contortus. Vet. Parasitol. 2002, 109, 59–63. [Google Scholar] [CrossRef]
- Pandey, R.; Kalra, A.; Tandon, S.; Mehrotra, N.; Singh, H.N.; Kumar, S. Essential oils as potent source of nematicidal compounds. J. Phytopathol. 2000, 148, 501–502. [Google Scholar] [CrossRef]
- Karpouhtsis, I.; Pardali, E.; Feggou, E.; Kokkini, S.; Scouras, Z.G.; Mavragani-Tsipidou, P. Insecticidal and genotoxic activities of oregano essential oils. J. Agric. Food Chem. 1998, 46, 1111–1115. [Google Scholar] [CrossRef]
- Konstantopoulou, I.; Vassilopoulou, L.; Mavragani-Tsipidou, P.; Scouras, Z.G. Insecticidal effects of essential oils. A study of the effects of essential oils extracted from eleven Greek aromatic plants on Drosophila auraria. Experientia 1992, 48, 616–619. [Google Scholar] [CrossRef]
- Valero, M.; Giner, M.J. Effects of antimicrobial components of essential oils on growth of Bacillus cereus INRA L2104 in and the sensory qualities of carrot broth. Int. J. Food Microbiol. 2006, 106, 90–94. [Google Scholar] [CrossRef]
- Oussalah, M.; Caillet, S.; Lacroix, M. Mechanism of action of Spanish oregano, Chinese cinnamon, and savory essential oils against cell membranes and walls of Escherichia coli O157:H7 and Listeria monocytogenes. J. Food Prot. 2006, 69, 1046–1055. [Google Scholar] [CrossRef]
- Utchariyakiat, I.; Surassmo, S.; Jaturanpinyo, M.; Khuntayaporn, P.; Chomnawang, M.T. Efficacy of cinnamon bark oil and cinnamaldehyde on anti-multidrug resistant Pseudomonas aeruginosa and the synergistic effects in combination with other antimicrobial agents. BMC Complement. Altern. Med. 2016, 16, 158. [Google Scholar] [CrossRef] [Green Version]
- Pinto, E.; Vale-Silva, L.; Cavaleiro, C.; Salgueiro, L. Antifungal activity of the clove essential oil from Syzygium aromaticum on Candida, Aspergillus and dermatophyte species. J. Med. Microbiol. 2009, 58, 1454–1462. [Google Scholar] [CrossRef]
- Jantan, I.b.; Karim Moharam, B.A.; Santhanam, J.; Jamal, J.A. Correlation between chemical composition and antifungal activity of the essential oils of eight Cinnamomum species. Pharmaceut. Biol. 2008, 46, 406–412. [Google Scholar] [CrossRef] [Green Version]
- Paster, N.; Menasherov, M.; Ravid, U.; Juven, B. Antifungal activity of oregano and thyme essential oils applied as fumigants against fungi attacking stored grain. J. Food Prot. 1995, 58, 81–85. [Google Scholar] [CrossRef]
- Marino, M.; Bersani, C.; Comi, G. Impedance measurements to study the antimicrobial activity of essential oils from Lamiaceae and Compositae. Int. J. Food Microbiol. 2001, 67, 187–195. [Google Scholar] [CrossRef]
- Marino, M.; Bersani, C.; Comi, G. Antimicrobial activity of the essential oils of Thymus vulgaris L. measured using a bioimpedometric method. J. Food Prot. 1999, 62, 1017–1023. [Google Scholar] [CrossRef]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods--a review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef]
- Lv, F.; Liang, H.; Yuan, Q.; Li, C. In vitro antimicrobial effects and mechanism of action of selected plant essential oil combinations against four food-related microorganisms. Food Res. Int. 2011, 44, 3057–3064. [Google Scholar] [CrossRef]
- Braga, M.E.M.; Leal, P.F.; Carvalho, J.E.; Meireles, M.A.A. Comparison of yield, composition, and antioxidant activity of turmeric (Curcuma longa L.) extracts obtained using various techniques. J. Agric. Food Chem. 2003, 51, 6604–6611. [Google Scholar] [CrossRef]
- Prakash, B.; Singh, P.; Mishra, P.K.; Dubey, N.K. Safety assessment of Zanthoxylum alatum Roxb. essential oil, its antifungal, antiaflatoxin, antioxidant activity and efficacy as antimicrobial in preservation of Piper nigrum L. fruits. Int. J. Food Microbiol. 2012, 153, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Jordán, M.J.; Martínez, R.M.; Goodner, K.L.; Baldwin, E.A.; Sotomayor, J.A. Seasonal variation of Thymus hyemalis Lange and Spanish Thymus vulgaris L. essential oils composition. Ind. Crop. Prod. 2006, 24, 253–263. [Google Scholar] [CrossRef]
- Fratini, F.; Mancini, S.; Turchi, B.; Friscia, E.; Pistelli, L.; Giusti, G.; Cerri, D. A novel interpretation of the Fractional Inhibitory Concentration Index: The case Origanum vulgare L. and Leptospermum scoparium J. R. et G. Forst essential oils against Staphylococcus aureus strains. Microbiol. Res. 2017, 195, 11–17. [Google Scholar] [CrossRef]
- Renneberg, J. Definitions of antibacterial interactions in animal infection models. J. Antimicrob. Chemother. 1993, 31 (Suppl. D), 167–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, P.M.; Parish, M.E. Methods for testing the efficacy of food antimicrobials. Food Technol. 1989, 43, 148–155. [Google Scholar]
- Jiang, Y.; Wu, N.; Fu, Y.-J.; Wang, W.; Luo, M.; Zhao, C.-J.; Zu, Y.-G.; Liu, X.-L. Chemical composition and antimicrobial activity of the essential oil of Rosemary. Environ. Toxicol. Pharmacol. 2011, 32, 63–68. [Google Scholar] [CrossRef]
- Noll, M.; Trunzer, K.; Vondran, A.; Vincze, S.; Dieckmann, R.; Al Dahouk, S.; Gold, C. Benzalkonium chloride induces a VBNC state in Listeria monocytogenes. Microorganisms 2020, 8, 184. [Google Scholar] [CrossRef] [Green Version]
- Vanhauteghem, D.; Demeyere, K.; Callaert, N.; Boelaert, A.; Haesaert, G.; Audenaert, K.; Meyer, E. Flow cytometry is a powerful tool for assessment of the viability of fungal conidia in metalworking fluids. Appl. Environ. Microbiol. 2017, 83, e00938-17. [Google Scholar] [CrossRef] [Green Version]
- Hyldgaard, M.; Mygind, T.; Meyer, R.L. Essential oils in food preservation: Mode of action, synergies, and interactions with food matrix components. Front. Microbiol. 2012, 3, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moleyar, V.; Narasimham, P. Antibacterial activity of essential oil components. Int. J. Food Microbiol. 1992, 16, 337–342. [Google Scholar] [CrossRef]
- Visvalingam, J.; Hernandez-Doria, J.D.; Holley, R.A. Examination of the genome-wide transcriptional response of Escherichia coli O157:H7 to cinnamaldehyde exposure. Appl. Environ. Microbiol. 2013, 79, 942–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arendrup, M.C.; Guinea, J.; Cuenca-Estrella, M.; Meletiadis, J.; Mouton, J.W.; Lagrou, K.; Howard, S.J. EUCAST Definitive Document E.DEF 9.3. Method for the Determination of Broth Dilution Minimum Inhibitory Concentrations of Antifungal Agents for Conidia Forming Moulds. 2015. Available online: https://www.aspergillus.org.uk/wp-content/uploads/2016/03/EUCAST_E_Def_9_3_Mould_testing_definitive_0.pdf (accessed on 7 October 2021).
- Arendrup, M.C.; Guinea, J.; Cuenca-Estrella, M.; Meletiadis, J.; Mouton, J.W.; Lagrou, K.; Howard, S.J. EUCAST DEFINITIVE DOCUMENT E.DEF 7.3.1 Method for the Determination of Broth Dilution Minimum Inhibitory Concentrations of Antifungal Agents for Yeasts. 2015. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/AFST/Files/EUCAST_E_Def_7_3_1_Yeast_testing__definitive.pdf (accessed on 7 October 2021).
- Bonapace, C.R.; White, R.L.; Friedrich, L.V.; Bosso, J.A. Evaluation of antibiotic synergy against Acinetobacter baumannii: A comparison with Etest, time-kill, and checkerboard methods. Diagn. Microbiol. Infect. Dis. 2000, 38, 43–50. [Google Scholar] [CrossRef]
- Bonapace, C.R.; Bosso, J.A.; Friedrich, L.V.; White, R.L. Comparison of methods of interpretation of checkerboard synergy testing. Diagn. Microbiol. Infect. Dis. 2002, 44, 363–366. [Google Scholar] [CrossRef]
- Heymich, M.-L.; Nißl, L.; Hahn, D.; Noll, M.; Pischetsrieder, M. Antioxidative, antifungal and additive activity of the antimicrobial peptides Leg1 and Leg2 from chickpea. Foods 2021, 10, 585. [Google Scholar] [CrossRef]
- Qu, S.; Yang, K.; Chen, L.; Liu, M.; Geng, Q.; He, X.; Li, Y.; Liu, Y.; Tian, J. Cinnamaldehyde, a promising natural preservative against Aspergillus flavus. Front. Microbiol. 2019, 10, 2895. [Google Scholar] [CrossRef] [Green Version]
- Shreaz, S.; Sheikh, R.A.; Rimple, B.; Hashmi, A.A.; Nikhat, M.; Khan, L.A. Anticandidal activity of cinnamaldehyde, its ligand and Ni(II) complex: Effect of increase in ring and side chain. Microb. Pathog. 2010, 49, 75–82. [Google Scholar] [CrossRef]
- Gehan, I. Kh, M.; Samir, A.M., A. Antifungal potential and biochemical effects of monoterpenes and phenylpropenes on plant. Plant Protect. Sci. 2017, 54, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Mitropoulou, G.; Nikolaou, A.; Santarmaki, V.; Sgouros, G.; Kourkoutas, Y. Citrus medica and Cinnamomum zeylanicum essential oils as potential biopreservatives against spoilage in low alcohol wine products. Foods 2020, 9, 577. [Google Scholar] [CrossRef] [PubMed]
- Hua, H.; Xing, F.; Selvaraj, J.N.; Wang, Y.; Zhao, Y.; Zhou, L.; Liu, X.; Liu, Y. Inhibitory effect of essential oils on Aspergillus ochraceus growth and ochratoxin A production. PLoS ONE 2014, 9, e108285. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.; Kong, J.; Ju, J.; Zhang, Y.; Guo, Y.; Cheng, Y.; Qian, H.; Xie, Y.; Yao, W. Membrane damage mechanism contributes to inhibition of trans-cinnamaldehyde on Penicillium italicum using Surface-Enhanced Raman Spectroscopy (SERS). Sci. Rep. 2019, 9, 490. [Google Scholar] [CrossRef]
- Wei, J.; Bi, Y.; Xue, H.; Wang, Y.; Zong, Y.; Prusky, D. Antifungal activity of cinnamaldehyde against Fusarium sambucinum involves inhibition of ergosterol biosynthesis. J. Appl. Microbiol. 2020, 129, 256–265. [Google Scholar] [CrossRef]
- Bakhtiari, S.; Jafari, S.; Taheri, J.B.; Kashi, T.S.J.; Namazi, Z.; Iman, M.; Poorberafeyi, M. The effects of cinnamaldehyde (cinnamon derivatives) and nystatin on Candida albicans and Candida glabrata. Open Access Maced. J. Med. Sci. 2019, 7, 1067–1070. [Google Scholar] [CrossRef] [PubMed]
- Ciani, M.; Menghini, L.; Mariani, F.; Pagiotti, R.; Menghini, A.; Fatichenti, F. Antimicrobial properties of essential oil of Satureja montana L. on pathogenic and spoilage yeasts. Biotechnol. Lett. 2000, 22, 1007–1010. [Google Scholar] [CrossRef]
- Xiang, F.; Zhao, Q.; Zhao, K.; Pei, H.; Tao, F. The efficacy of composite essential oils against aflatoxigenic fungus Aspergillus flavus in maize. Toxins 2020, 12, 562. [Google Scholar] [CrossRef]
- Wang, Y.; Feng, K.; Yang, H.; Yuan, Y.; Yue, T. Antifungal mechanism of cinnamaldehyde and citral combination against Penicillium expansum based on FT-IR fingerprint, plasma membrane, oxidative stress and volatile profile. RSC Adv. 2018, 8, 5806–5815. [Google Scholar] [CrossRef] [Green Version]
- Helal, G.A.; Sarhan, M.M.; Abu Shahla, A.N.K.; Abou El-Khair, E.K. Effects of Cymbopogon citratus L. essential oil on the growth, lipid content and morphogenesis of Aspergillus niger ML2-strain. J. Basic Microbiol. 2006, 46, 456–469. [Google Scholar] [CrossRef]
- Wang, S.; Zheng, Y.; Xiang, F.; Li, S.; Yang, G. Antifungal activity of Momordica charantia seed extracts toward the pathogenic fungus Fusarium solani L. J. Food Drug Anal. 2016, 24, 881–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OuYang, Q.; Duan, X.; Li, L.; Tao, N. Cinnamaldehyde exerts its antifungal activity by disrupting the cell wall integrity of Geotrichum citri-aurantii. Front. Microbiol. 2019, 10, 55. [Google Scholar] [CrossRef]
- Bang, K.H.; Lee, D.W.; Park, H.M.; Rhee, Y.H. Inhibition of fungal cell wall synthesizing enzymes by trans-cinnamaldehyde. Biosci. Biotechnol. Biochem. 2000, 64, 1061–1063. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Wang, D.; Liu, L.; Chen, J.; Xue, Y.; Shi, Z. Ca(2+) efflux is involved in cinnamaldehyde-induced growth inhibition of Phytophthora capsici. PLoS ONE 2013, 8, e76264. [Google Scholar] [CrossRef] [Green Version]
- Chami, F.; Chami, N.; Bennis, S.; Bouchikhi, T.; Remmal, A. Oregano and clove essential oils induce surface alteration of Saccharomyces cerevisiae. Phytother. Res. 2005, 19, 405–408. [Google Scholar] [CrossRef] [PubMed]
- Höferl, M.; Stoilova, I.; Schmidt, E.; Wanner, J.; Jirovetz, L.; Trifonova, D.; Krastev, L.; Krastanov, A. Chemical composition and antioxidant properties of Juniper Berry (Juniperus communis L.) essential oil. Action of the essential oil on the antioxidant protection of Saccharomyces cerevisiae model organism. Antioxidants 2014, 3, 81–98. [Google Scholar] [CrossRef] [Green Version]
- López, P.; Sánchez, C.; Batlle, R.; Nerín, C. Development of flexible antimicrobial films using essential oils as active agents. J. Agric. Food Chem. 2007, 55, 8814–8824. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| EOCs and Control | Amphotericin B | CAR | CA | CI | CLA | CLO | EU | GE | IEU | LM | LN | Vanillin |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A. niger | 4 | 3904 | 262.5 | 888 | 1714 | 1710 | 2134 | 1758 | 1082 | >3440 | 1740 | 1000 |
| F. solani | 8 | 3904 | 262.5 | 1776 | 1714 | 3420 | 2134 | 1758 | 2164 | >3440 | 3480 | 2000 |
| P. funiculosum | >16 | 3904 | 131.3 | 444 | 428.5 | 855 | 1067 | 879 | 1082 | 1720 | 870 | 500 |
| C. parapsilosis | 2 | 3904 | 262.5 | 888 | 1714 | 3420 | 4268 | 1758 | 4328 | >3440 | 6960 | 2000 |
| D. hansenii | >4 | 1952 | 131.3 | 444 | 857 | 855 | 2134 | 879 | 2164 | 860 | 870 | 1000 |
| D. bruxellensis | 0.25 | 1952 | 32.81 | 222 | 107.1 | 3420 | 2134 | 1758 | 2164 | 3440 | 3480 | 2000 |
| H. anomala | 0.5 | 3904 | 32.81 | 222 | 428.5 | 3420 | 4268 | 1758 | 4328 | 1720 | 3480 | 1000 |
| I. orientalis | 2 | >3904 | 262.5 | 888 | 1714 | 6840 | 4268 | 3516 | 4328 | 3440 | 6960 | 4000 |
| K. marxianus | 2 | 3904 | 262.5 | 444 | 857 | 3420 | 4268 | 1758 | 4328 | 3440 | 3480 | 4000 |
| P. membranifaciens | 4 | 1952 | 131.3 | 222 | 428.5 | 1710 | 2134 | 439.5 | 2164 | 1720 | 1740 | 1000 |
| S. cerevisiae | 4 | 3904 | 131.3 | 222 | 428.5 | 3420 | 4268 | 1758 | 4328 | 3440 | 3480 | 4000 |
| S. octosporus | 1 | 3904 | 65.63 | 111 | 428.5 | 1710 | 2134 | 1758 | 4328 | 3440 | 3480 | 2000 |
| S. pombe | 1 | 3904 | 328.1 | 111.5 | 428.5 | 3420 | 2134 | 1758 | 4328 | 3440 | 3480 | 2000 |
| Mold and Yeast Strains | CA + CI | CA + CLA | CA + CLO | CA + EU | CA + GE | CA+ IEU | CA + LN |
|---|---|---|---|---|---|---|---|
| A. niger | 1.0 ± 0.1 | 1.0 ± 0.1 | 1.5 ± 0.2 | 1.3 ± 0.1 | 1.1 ± 0.1 | 1.2 ± 0.0 | 1.1 ± 0.1 |
| F. solani | 1.0 ± 0.1 | 1.0 ± 0.0 | 1.1 ± 0.1 | 1.1 ± 0.1 | 1.1 ± 0.1 | 1.1 ± 0.1 | 1.0 ± 0.1 |
| P. funiculosum | 0.9 ± 0.2 | 1.0 ± 0.1 | 1.1 ± 0.1 | 1.1 ± 0.0 | 1.2 ± 0.2 | 1.0 ± 0.1 | 1.1 ± 0.3 |
| C. parapsilosis | 0.9 ± 0.1 | 1.0 ± 0.1 | 1.0 ± 0.0 | 1.0 ± 0.1 | 1.1 ± 0.0 | 1.2 ± 0.1 | 0.9 ± 0.1 |
| D. hansenii | 1.1 ± 0.1 | 1.0 ± 0.1 | 1.1 ± 0.1 | 1.2 ± 0.1 | 1.0 ± 0.1 | 1.2 ± 0.1 | 1.2 ± 0.1 |
| D. bruxellensis | 0.8 ± 0.1 | 0.9 ± 0.1 | 1.1 ± 0.0 | 1.0 ± 0.1 | 0.9 ± 0.1 | 1.1 ± 0.1 | 0.9 ± 0.0 |
| H. anomala | 1.3 ± 0.1 | 1.1 ± 0.1 | 1.5 ± 0.1 | 1.3 ± 0.2 | 1.5 ± 0.1 | 1.5 ± 0.3 | 1.4 ± 0.3 |
| I. orientalis | 0.9 ± 0.1 | 0.8 ± 0.1 | 1.1 ± 0.1 | 1.2 ± 0.1 | 1.0 ± 0.0 | 1.1 ± 0.1 | 1.0 ± 0.1 |
| K. marxianus | 1.0 ± 0.1 | 1.1 ± 0.1 | 1.1 ± 0.1 | 1.1 ± 0.1 | 1.0 ± 0.1 | 1.3 ± 0.1 | 1.1 ± 0.0 |
| P. membranifaciens | 0.9 ± 0.1 | 0.6 ± 0.1 | 1.1 ± 0.1 | 0.8 ± 0.0 | 1.1 ± 0.1 | 0.8 ± 0.0 | 0.9 ± 0.2 |
| S. cerevisiae | 1.0 ± 0.1 | 0.6 ± 0.1 | 1.1 ± 0.1 | 0.9 ± 0.1 | 1.1 ± 0.0 | 1.0 ± 0.1 | 1.1 ± 0.0 |
| S. octosporus | 1.0 ± 0.1 | 0.9 ± 0.1 | 0.9 ± 0.1 | 0.7 ± 0.1 | 0.7 ± 0.1 | 0.7 ± 0.1 | 1.0 ± 0.1 |
| S. pombe | 0.9 ± 0.1 | 0.5 ± 0.1 | 1.0 ± 0.1 | 0.8 ± 0.1 | 0.9 ± 0.0 | 0.9 ± 0.1 | 0.9 ± 0.1 |
| Treatment | NC | 2 MIC | 1 MIC | 0.5 MIC | 0.25 MIC | 0.125 MIC | PC |
|---|---|---|---|---|---|---|---|
| PC | 16 h | 17 h | 17 h | 17 h | n.s. | n.s. | |
| 0.125 MIC | 13 h | 14 h | 14 h | 19 h | n.s. | ||
| 0.25 MIC | 19 h | 19 h | 19 h | n.s. | |||
| 0.5 MIC | 25 h | 25 h | 25 h | ||||
| 1 MIC | n.s. | n.s. | |||||
| 2 MIC | n.s. | ||||||
| NC | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nißl, L.; Westhaeuser, F.; Noll, M. Antimycotic Effects of 11 Essential Oil Components and Their Combinations on 13 Food Spoilage Yeasts and Molds. J. Fungi 2021, 7, 872. https://doi.org/10.3390/jof7100872
Nißl L, Westhaeuser F, Noll M. Antimycotic Effects of 11 Essential Oil Components and Their Combinations on 13 Food Spoilage Yeasts and Molds. Journal of Fungi. 2021; 7(10):872. https://doi.org/10.3390/jof7100872
Chicago/Turabian StyleNißl, Laura, Florian Westhaeuser, and Matthias Noll. 2021. "Antimycotic Effects of 11 Essential Oil Components and Their Combinations on 13 Food Spoilage Yeasts and Molds" Journal of Fungi 7, no. 10: 872. https://doi.org/10.3390/jof7100872