
Geometry Does Impact on the Plane Strain Directions of the Human Left Ventricle, Irrespective of Disease
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
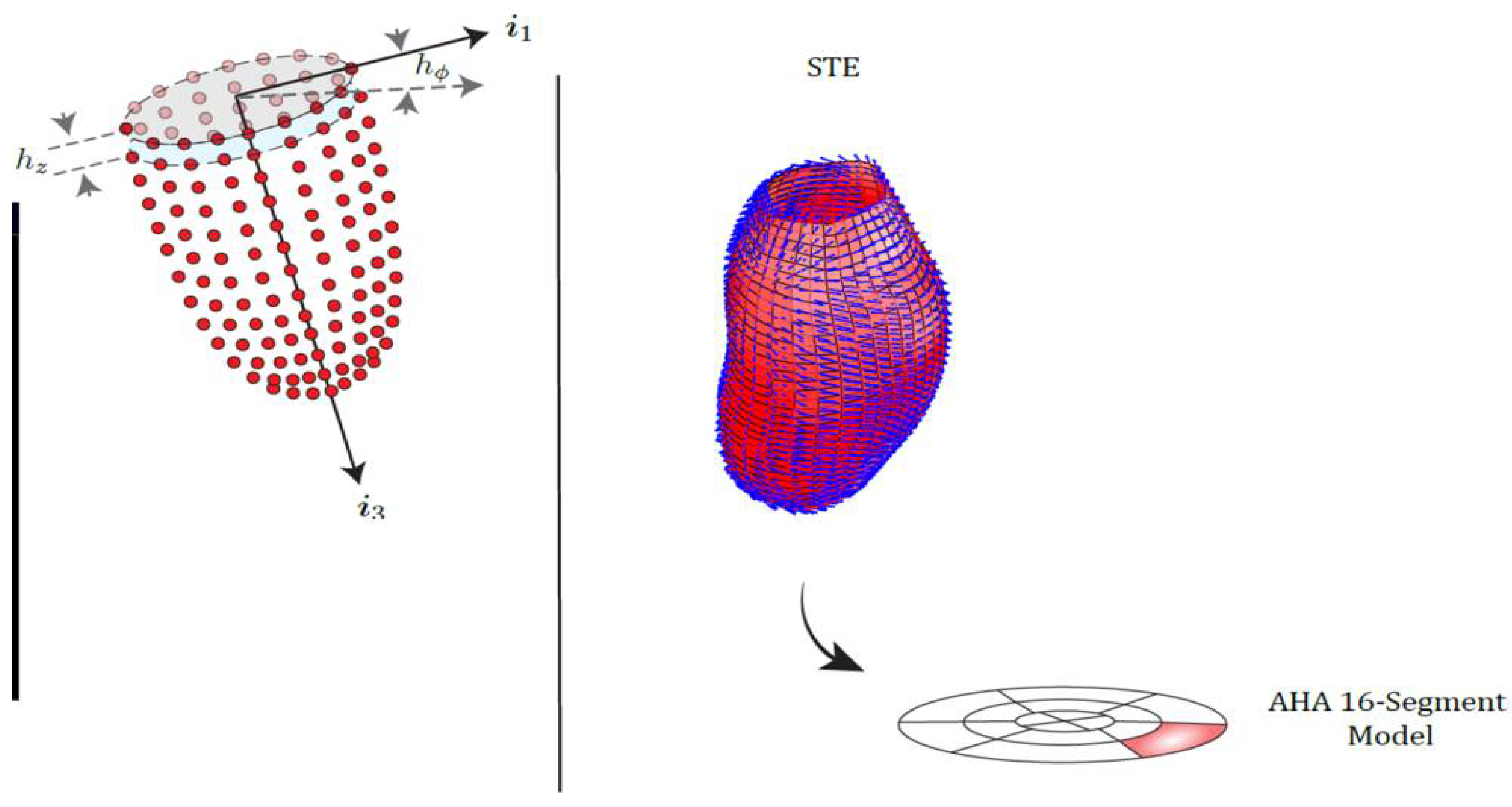
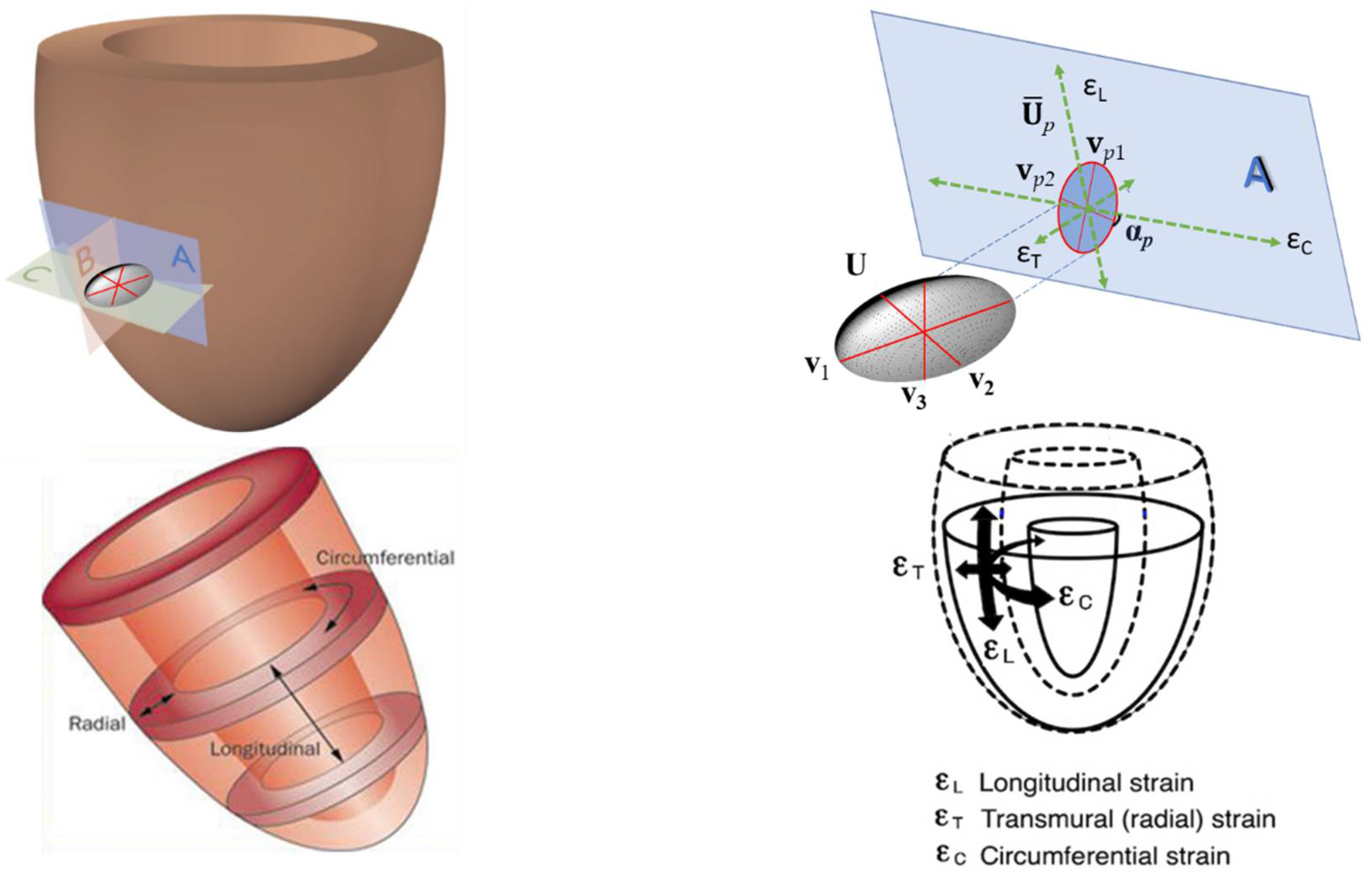
2.2. Strains, Tensors, Strain Directions and More
2.3. Statistical Analysis and Visualization
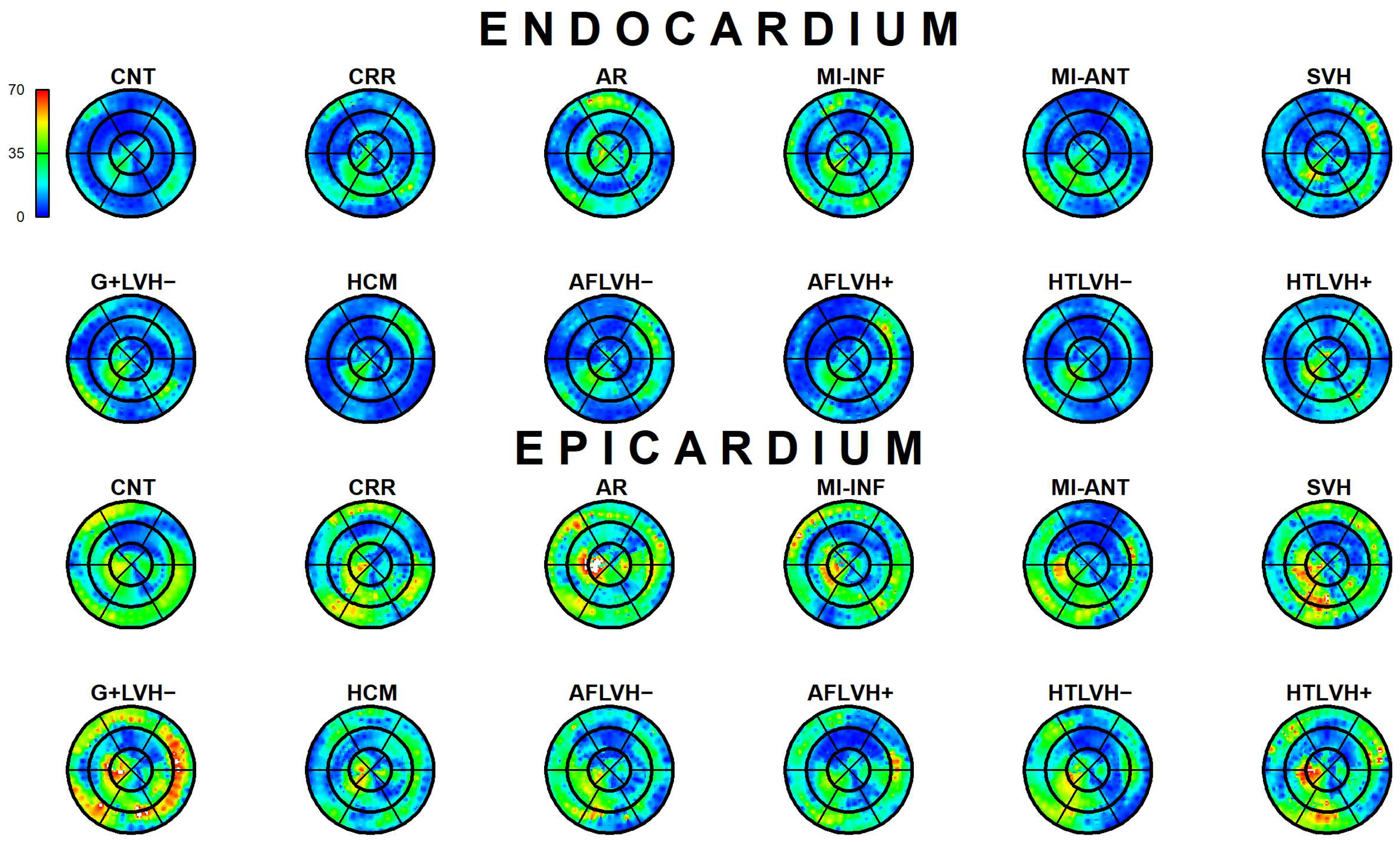
2.3.1. Epi-Endo Comparison
2.3.2. Control vs. Diseased Patients Comparison
2.3.3. PSD Angle Differences vs. Static Curvature Ratio
2.3.4. Results Visualization
3. Results
3.1. Epi-Endo Comparison
3.2. Control vs. Diseased Comparison
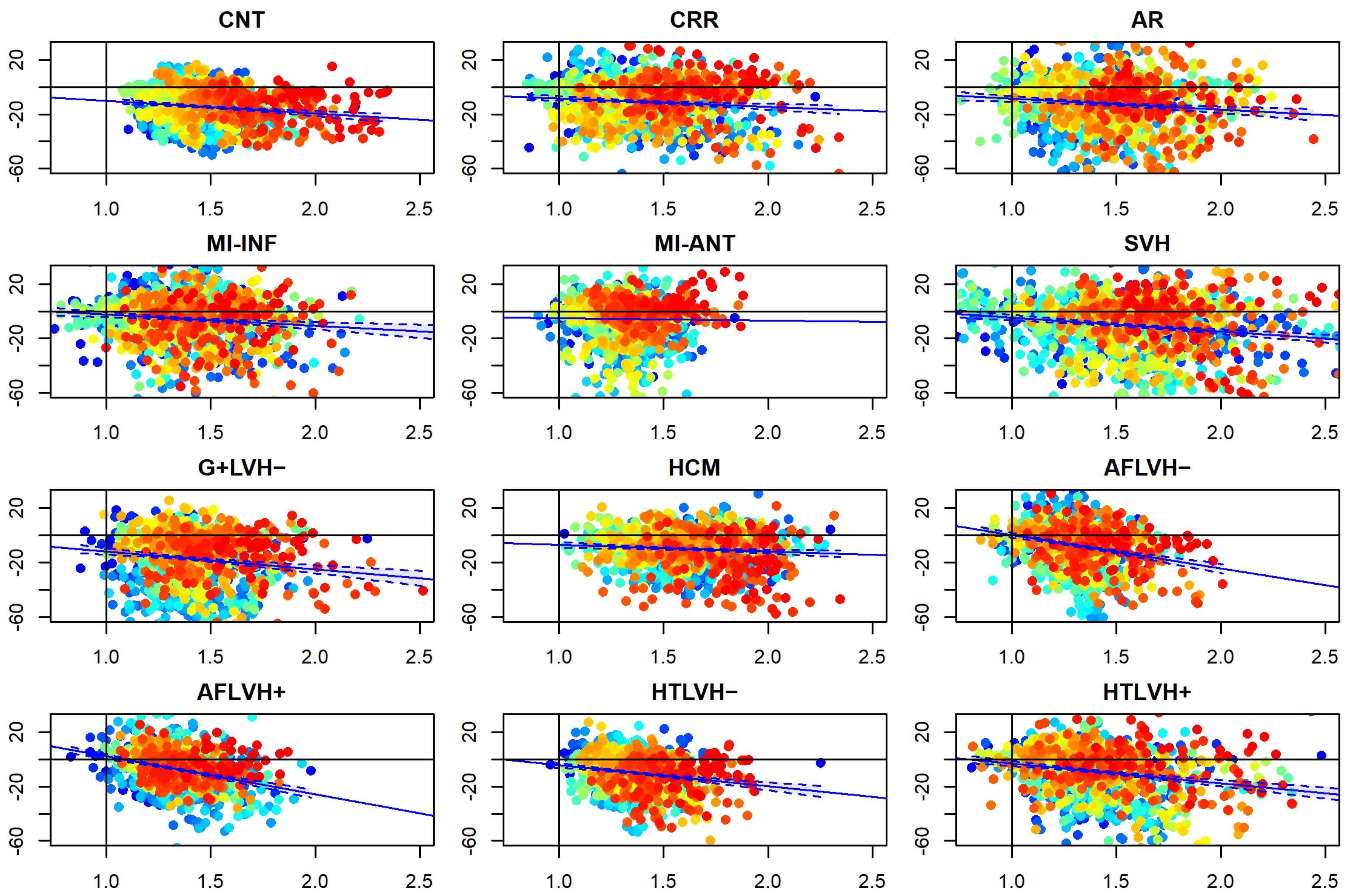
3.3. αp Differences vs. Static Curvature’ Ratio
4. Discussion
5. Limitation of the Study
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arts, T.; Bovendeerd, P.; Delhaas, T.; Prinzen, F. Modeling the relation between cardiac pump function and myofiber mechanics. J. Biomech. 2003, 36, 731–736. [Google Scholar] [CrossRef]
- Bovendeerd, P.H.M.; Arts, T.; Huyghe, J.M.; Van Campen, D.H.; Reneman, R.S. Dependence of local left ventricular wall mechanics on myocardial fiber orientation: A model study. J. Biomech. 1992, 25, 1129–1140. [Google Scholar] [CrossRef]
- Costa, K.D.; Holmes, J.W.; McCulloch, A.D. Modelling cardiac mechanical properties in three dimensions. Phil. Trans. Roy. Soc. Lond. A Math. Phys. Eng. Sci. 2001, 359, 1233–1250. [Google Scholar] [CrossRef]
- Freeman, G.L.; LeWinter, M.M.; Engler, R.L.; Covell, J.W. Relationship between myocardial fiber direction and segment shortening in the midwall of the canine left ventricle. Circ. Res. 1985, 56, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Vetter, F.J.; McCulloch, A.D. Three-dimensional stress and strain in passive rabbit left ventricle: A model study. Ann. Biomed. Eng. 2000, 28, 781–792. [Google Scholar] [CrossRef]
- Hooks, D.A.; Tomlinson, K.A.; Marsden, S.G.; LeGrice, I.J.; Smaill, B.H.; Pullan, A.J.; Hunter, P.J. Cardiac microstructure: Implications for electrical propagation and defibrillation in the heart. Circ. Res. 2002, 91, 331–338. [Google Scholar] [CrossRef] [Green Version]
- Arts, T.; Prinzen, F.W.; Snoeckx, L.H.; Rijcken, J.M.; Reneman, R.S. Adaptation of cardiac structure by mechanical feedback in the environment of the cell: A model study. Biophys. J. 1994, 66, 953–961. [Google Scholar] [CrossRef] [Green Version]
- Van Der Bel-Kahn, J. Muscle fiber disarray in common heart diseases. Am. J. Cardiol. 1977, 40, 355–364. [Google Scholar] [CrossRef]
- Engelmayr, G.C., Jr.; Cheng, M.; Bettinger, C.J.; Borenstein, J.T.; Langer, R.; Freed, L.E. Accordion-like honeycombs for tissue engineering of cardiac anisotropy. Nat. Mater. 2008, 7, 1003–1010. [Google Scholar] [CrossRef]
- Agger, P.; Stephenson, R.S. Assessing Myocardial Architecture: The Challenges and Controversies. J. Cardiovasc. Dev. Dis. 2020, 7, 47. [Google Scholar] [CrossRef]
- Torrent-Guasp, F. Anatomia Funcional del Corazon; Paz Montalvo: Madrid, Spain, 1957; pp. 62–68. [Google Scholar]
- Gilbert, S.H.; Benson, A.P.; Li, P.; Holden, A.V. Regional localisation of left ventricular sheet structure: Integration with current models of cardiac fibre, sheet and band structure. Eur. J. Cardio-Thorac. Surg. 2007, 32, 231–249. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.H.; Ho, S.Y.; Sanchez-Quintana, D.; Redmann, K.; Lunkenheimer, P.P. Heuristic problems in defining the three-dimensional arrangement of the ventricular myocytes. Anat. Rec. Part A Discov. Mol. Cell. Evol. Biol. 2006, 288, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Smerup, M.; Nielsen, E.; Agger, P.; Frandsen, J.; Vestergaard-Poulsen, P.; Andersen, J.; Nyengaard, J.; Pedersen, M.; Ringgaard, S.; Hjortdal, V.; et al. The three-dimensional arrangement of the myocytes aggregated together within the mammalian ventricular myocardium. Anat. Rec. Adv. Int. Anat. Evol. Biol. 2009, 292, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Pedrizzetti, G.; Kraigher-Krainer, E.; De Luca, A.; Caracciolo, G.; Mangual, J.O.; Shah, A.; Toncelli, L.; Domenichini, F.; Tonti, G.; Galanti, G.; et al. Functional strain-line pattern in the human left ventricle. Phys. Rev. Lett. 2012, 109, 048103. [Google Scholar] [CrossRef]
- Gabriele, S.; Nardinocchi, P.; Varano, V. Evaluation of the strain-line patterns in a human left ventricle: A simulation study. Comp. Methods Biomech. Biomed. Eng. 2015, 18, 790–798. [Google Scholar] [CrossRef]
- Gabriele, S.; Teresi, L.; Varano, V.; Evangelista, A.; Nardinocchi, P.; Puddu, P.E.; Torromeo, C. A comparative analysis of the strain-line pattern in the human left ventricle: Experiments vs modelling. Comp. Methods Biomech. Biomed. Eng. Imaging Vis. 2014, 4, 164–173. [Google Scholar]
- Lee, S.; Choi, S.; Kim, S.; Jeong, Y.; Lee, K.; Hur, S.H.; Lee, S.R.; Lee, E.J.; Sin, M.J.; Kim, N.; et al. Validation of three-dimensional echocardiographic principal strain analysis for assessing left ventricular contractility: An animal study. Med. Phys. 2019, 46, 2137–2144. [Google Scholar] [CrossRef]
- Colorado-Cervantes, J.I.; Nardinocchi, P.; Piras, P.; Sansalone, V.; Teresi, L.; Torromeo, C.; Puddu, P.E. Patient-specific modeling of left ventricle mechanics. Acta Mech. Sin. 2022, 38, 621211. [Google Scholar] [CrossRef]
- Piras, P.; Torromeo, C.; Evangelista, A.; Esposito, G.; Nardinocchi, P.; Teresi, L.; Madeo, A.; Re, F.; Chialastri, C.; Schiariti, M.; et al. Non-invasive prediction of genotype positive–phenotype negative in hypertrophic cardiomyopathy by 3D modern shape analysis. Exp. Phys. 2019, 104, 1688–1700. [Google Scholar] [CrossRef]
- Evangelista, A.; Gabriele, S.; Nardinocchi, P.; Piras, P.; Puddu, P.E.; Teresi, L.; Torromeo, C.; Varano, V. Non-invasive assessment of functional strain lines in the real human left ventricle via speckle tracking echocardiography. J. Biomech. 2015, 48, 465–471. [Google Scholar] [CrossRef]
- Piras, P.; Profico, A.; Pandolfi, L.; Raia, P.; Di Vincenzo, F.; Mondanaro, A.; Castiglione, S.; Varano, V. Current options for visualization of local deformation in modern shape analysis applied to paleobiological case studies. Front. Earth Sci. 2020, 8, 66. [Google Scholar] [CrossRef]
- Varano, V.; Piras, P.; Gabriele, S.; Teresi, L.; Nardinocchi, L.; Dryden, I.L.; Torromeo, C.; Puddu, P.E. The decomposition of deformation: New metrics to enhance shape analysis in medical imaging. Med. Image Anal. 2018, 46, 35–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piras, P.; Torromeo, C.; Evangelista, A.; Gabriele, S.; Esposito, G.; Nardinocchi, P.; Teresi, L.; Madeo, A.; Schiariti, M.; Varano, V.; et al. Homeostatic Left Heart integration and disintegration links atrio-ventricular covariation’s dyshomeostasis in Hypertrophic Cardiomyopathy. Sci. Rep. 2017, 24, 6257. [Google Scholar] [CrossRef] [PubMed]
- Mangiafico, S. Summary and Analysis of Extension Program Evaluation in R; Rutgers Cooperative Extension: New Brunswick, NJ, USA, 2016. [Google Scholar]
- Azhari, H.; Weiss, J.L.; Rogers, W.J.; Siu, C.O.; Zerhouni, E.A.; Shapiro, E.P. Noninvasive quantification of principal strains in normal canine hearts using tagged mri images in 3-D. Am. J. Phys. 1993, 264, 205–216. [Google Scholar] [CrossRef]
- MacGowan, G.A.; Shapiro, E.P.; Azhari, H.; Siu, C.O.; Hees, P.S.; Hutchins, G.M.; Weiss, J.L.; Rademakers, F.E. Noninvasive measurement of shortening in the fiber and cross-fiber directions in the normal human left ventricle and in idiopathic dilated cardiomyopathy. Circulation 1997, 96, 535–541. [Google Scholar] [CrossRef]
- Waldman, L.K.; Nosan, D.; Villarreal, F.; Covell, J.W. Relation between transmural deformation and local myofiber direction in canine left ventricle. Circ. Res. 1988, 63, 550–562. [Google Scholar] [CrossRef] [Green Version]
- Esposito, G.; Piras, P.; Evangelista, A.; Nuzzi, V.; Nardinocchi, P.; Pannarale, G.; Torromeo, C.; Puddu, P.E. Improving performance values of 3D speckle tracking in arterial hypertension and paroxysmal atrial fibrillation by using novel strain parameters. Sci. Rep. 2019, 9, 7382. [Google Scholar] [CrossRef] [Green Version]
- Piras, P.; Teresi, L.; Puddu, P.E.; Torromeo, C.; Young, A.A.; Suinesiaputra, A.; Medrano-Gracia, P. Morphologically normalized left ventricular motion indicators from MRI feature tracking characterize myocardial infarction. Sci. Rep. 2017, 25, 12259. [Google Scholar] [CrossRef] [Green Version]
- Muraru, D.; Niero, A.; Rodriguez-Zanella, H.; Cherata, D.; Badano, L. Three-dimensional speckle-tracking echocardiography: Benefits and limitations of integrating myocardial mechanics with three-dimensional imaging. Cardiovasc. Diagn. Ther. 2018, 8, 101–117. [Google Scholar] [CrossRef] [Green Version]
- Henglin, M.; Stein, G.; Hushcha, P.V.; Snoek, J.; Wiltschko, A.B.; Cheng, S. Machine learning approaches in cardiovascular imaging. Circ. Cardiovasc. Imaging 2017, 10, e005614. [Google Scholar] [CrossRef]
- Nayak, R.; Schifitto, G.; Doyley, M.M. Visualizing angle-independent principal strains in the longitudinal view of the carotid artery: Phantom and in vivo evaluation. Ultrasound Med. Biol. 2018, 44, 1379–1391. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Description | N |
|---|---|---|
| CNT | control healthy subjects | 82 |
| CRR | cirrhosis | 13 |
| AR | aortic regurgitation | 4 |
| MI-INF | myocardial infarction of inferior wall | 4 |
| MI-ANT | myocardial infarction of anterior wall | 11 |
| SVH | secondary ventricular hypertrophy | 7 |
| G+LVH− | mutation carriers for hypertrophy without LV hypertrophy | 7 |
| HCM | mutation carriers for hypertrophy with LV hypertrophy | 18 |
| AFLVH− | atrial fibrillation without ventricular hypertrophy | 9 |
| AFLVH+ | atrial fibrillation with ventricular hypertrophy | 11 |
| HTLVH− | hypertension without LV hypertrophy | 18 |
| HTLVH+ | hypertension with LV hypertrophy | 9 |
| Group\Variables | CNT N = 82 | CRR N = 13 | AR N = 4 | MI-INF N = 4 | MI-ANT N = 11 | SVH N = 7 | G+LVH− N = 7 | HCM N = 18 | AFLVH− N = 9 | AFLVH+ N = 11 | HTLVH− N = 18 | HTLVH+ N = 9 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sex (M/F) | 48/34 | n = 82 | 11/2 | n = 13 | 2/2 | n = 4 | 3/1 | n = 4 | 10/1 | n = 11 | 6/1 | n = 7 | 3/4 | n = 7 | 12/6 | n = 18 | 8/1 | n = 9 | 6/5 | n = 11 | 10/8 | n = 18 | 8/1 | n = 9 |
| Mean ± sd | n | Mean ± sd | n | Mean ± sd | n | Mean ± sd | n | Mean ± sd | n | Mean ± sd | n | Mean ± sd | n | Mean ± sd | n | Mean ± sd | n | Mean ± sd | n | Mean ± sd | n | Mean ± sd | n | |
| Age, years | 38.531 ± 7.877 | n = 81 | 57.077 ± 6.689 | n = 13 | 61 ± 22.316 | n = 4 | 58.25 ± 8.382 | n = 4 | 60 ± 9.274 | n = 11 | 57.286 ± 12.645 | n = 7 | 45.143 ± 12.812 | n = 7 | 48.5 ± 13.544 | n = 18 | 59.778 ± 13.8 | n = 9 | 71.636 ± 3.957 | n = 11 | 56.389 ± 7.578 | n = 18 | 57.667 ± 10.186 | n = 9 |
| Ejection Fraction, % | 54.82 ± 8.679 | n = 82 | 54.435 ± 7.321 | n = 13 | 53.081 ± 9.729 | n = 4 | 59.858 ± 6.912 | n = 4 | 52.023 ± 5.123 | n = 11 | 51.896 ± 9.184 | n = 7 | 59.061 ± 3.598 | n = 7 | 58.653 ± 7.828 | n = 18 | 54.143 ± 6.921 | n = 9 | 56.399 ± 7.5 | n = 11 | 55.643 ± 10.143 | n = 18 | 56.357 ± 3.921 | n = 9 |
| Weight, kg | 71.409 ± 12.27 | n = 66 | 71.385 ± 10.389 | n = 13 | 64.25 ± 15.521 | n = 4 | 83.25 ± 18.392 | n = 4 | 81.909 ± 9.235 | n = 11 | 76.286 ± 6.473 | n = 7 | 72 ± 13.416 | n = 7 | 76.111 ± 14.768 | n = 18 | 85.444 ± 10.039 | n = 9 | 74.455 ± 13.299 | n = 11 | 75.944 ± 9.716 | n = 18 | 80.556 ± 10.477 | n = 9 |
| BSA, m2 | 1.841 ± 0.193 | n = 66 | 1.831 ± 0.157 | n = 13 | 1.663 ± 0.216 | n = 4 | 1.94 ± 0.244 | n = 4 | 2.06 ± 0.111 | n = 6 | 1.853 ± 0.065 | n = 7 | 1.813 ± 0.166 | n = 7 | 1.869 ± 0.175 | n = 15 | 2.001 ± 0.203 | n = 7 | 1.908 ± 0.175 | n = 4 | 1.874 ± 0.155 | n = 18 | 1.944 ± 0.15 | n = 9 |
| BMI, Kg/m2 | 23.909 ± 3.091 | n = 66 | 24.103 ± 2.67 | n = 13 | 23.208 ± 1.461 | n = 4 | 29.105 ± 3.221 | n = 4 | 26.666 ± 2.327 | n = 11 | 27.396 ± 3.694 | n = 7 | 25.993 ± 4.569 | n = 7 | 25.869 ± 3.9 | n = 18 | 27.437 ± 2.178 | n = 9 | 25.985 ± 3.97 | n = 11 | 26.447 ± 2.938 | n = 18 | 27.039 ± 2.547 | n = 9 |
| Interv. Septum, mm | 8.197 ± 1.449 | n = 66 | 10.154 ± 1.345 | n = 13 | 10.25 ± 2.5 | n = 4 | 10.5 ± 2.082 | n = 4 | 10.091 ± 1.375 | n = 11 | 9.286 ± 2.289 | n = 7 | 9.571 ± 1.618 | n = 7 | 18 ± 4.256 | n = 18 | 9.444 ± 1.81 | n = 9 | 11.636 ± 2.111 | n = 11 | 9.444 ± 1.542 | n = 18 | 11.667 ± 1.225 | n = 9 |
| PAS, mmHg | 116.406 ± 9.614 | n = 64 | 125.615 ± 9.124 | n = 13 | 118.75 ± 6.292 | n = 4 | 121.25 ± 10.308 | n = 4 | 114.091 ± 8.006 | n = 11 | 134.286 ± 11.701 | n = 7 | 122.857 ± 4.88 | n = 7 | 123.824 ± 6.002 | n = 17 | 127.778 ± 11.756 | n = 9 | 128.182 ± 10.787 | n = 11 | 129.722 ± 8.309 | n = 18 | 130 ± 11.18 | n = 9 |
| PAD, mmHg | 73.984 ± 7.724 | n = 64 | 77.538 ± 8.038 | n = 13 | 70 ± 8.165 | n = 4 | 75 ± 9.129 | n = 4 | 70 ± 8.062 | n = 11 | 84.286 ± 4.499 | n = 7 | 75.714 ± 5.345 | n = 7 | 78.235 ± 5.574 | n = 17 | 79.444 ± 3.909 | n = 9 | 75.455 ± 5.222 | n = 11 | 82.778 ± 7.117 | n = 18 | 81.667 ± 7.906 | n = 9 |
| Group | Intercept | p-Value_Intercept | Beta | p-Value_Beta | R_Squared |
|---|---|---|---|---|---|
| Systole | |||||
| CNT | −0.607 | 0.817 | −9.427 | 7.57 × 10−8 | 0.024 |
| CRR | −2.112 | 0.391 | −6.164 | 3.24 × 10−4 | 0.011 |
| AR | −0.691 | 0.822 | −8.018 | 1.25 × 10−4 | 0.012 |
| MI-INF | 5.708 | 0.053 | −8.185 | 9.51 × 10−5 | 0.013 |
| SVH | −3.593 | 0.342 | −1.579 | 0.576 | 0 |
| MI-ANT | 5.647 | 0.02 | −10.307 | 1.08 × 10−11 | 0.038 |
| G+LVH− | 1.467 | 0.691 | −13.208 | 2.81 × 10−7 | 0.022 |
| HCM | −2.248 | 0.394 | −4.833 | 0.003 | 0.007 |
| AFLVH− | 24.488 | 1.76 × 10−13 | −24.431 | 3.37 × 10−22 | 0.076 |
| HTLVH− | 29.934 | 9.26 × 10−20 | −27.755 | 5.87 × 10−29 | 0.1 |
| HTLVH+ | 11.208 | 4.00 × 10−4 | −15.447 | 1.65 × 10−11 | 0.038 |
| AFLVH+ | 11.592 | 2.64 × 10−5 | −14.542 | 4.19 × 10−14 | 0.047 |
| Diastole | |||||
| CNT | 23.409 | 1.15 × 10−6 | −29.695 | 3.7 × 10−15 | 0.051 |
| CRR | 5.217 | 0.333 | −12.75 | 0.003 | 0.007 |
| AR | 32.119 | 6.05 × 10−6 | −34.933 | 4.1 × 10−10 | 0.032 |
| MI-INF | 43.368 | 1.88 × 10−14 | −40.097 | 4.7 × 10−8 | 0.061 |
| SVH | 8.12 | 0.172 | −11.331 | 0.02 | 0.005 |
| MI-ANT | 12.818 | 0.025 | −17.726 | 4.4 × 10−5 | 0.014 |
| G+LVH− | 14.257 | 0.03 | −25.162 | 1.4 × 10−6 | 0.019 |
| HCM | 14.313 | 2.18 × 10−4 | −17.576 | 3.6 × 10−10 | 0.033 |
| AFLVH− | 39.764 | 2.81 × 10−10 | −39.072 | 5.3 × 10−14 | 0.047 |
| HTLVH− | 49.612 | 7.53 × 10−16 | −46.261 | 5.1 × 10−20 | 0.068 |
| HTLVH+ | 35.213 | 5.54 × 10−12 | −36.41 | 9.9 × 10−19 | 0.064 |
| AFLVH+ | 29.519 | 7.87 × 10−8 | −30.599 | 2.3 × 10−12 | 0.041 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piras, P.; Colorado-Cervantes, I.; Nardinocchi, P.; Gabriele, S.; Varano, V.; Esposito, G.; Teresi, L.; Torromeo, C.; Puddu, P.E. Geometry Does Impact on the Plane Strain Directions of the Human Left Ventricle, Irrespective of Disease. J. Cardiovasc. Dev. Dis. 2022, 9, 393. https://doi.org/10.3390/jcdd9110393
Piras P, Colorado-Cervantes I, Nardinocchi P, Gabriele S, Varano V, Esposito G, Teresi L, Torromeo C, Puddu PE. Geometry Does Impact on the Plane Strain Directions of the Human Left Ventricle, Irrespective of Disease. Journal of Cardiovascular Development and Disease. 2022; 9(11):393. https://doi.org/10.3390/jcdd9110393
Chicago/Turabian StylePiras, Paolo, Ivan Colorado-Cervantes, Paola Nardinocchi, Stefano Gabriele, Valerio Varano, Giuseppe Esposito, Luciano Teresi, Concetta Torromeo, and Paolo Emilio Puddu. 2022. "Geometry Does Impact on the Plane Strain Directions of the Human Left Ventricle, Irrespective of Disease" Journal of Cardiovascular Development and Disease 9, no. 11: 393. https://doi.org/10.3390/jcdd9110393