
White Sparkling Wine Proteins and Glycoproteins and Their Behavior in Foam Expansion and Stability

Abstract
:1. Introduction
2. Materials and Methods
2.1. Extraction of Protein Fractions
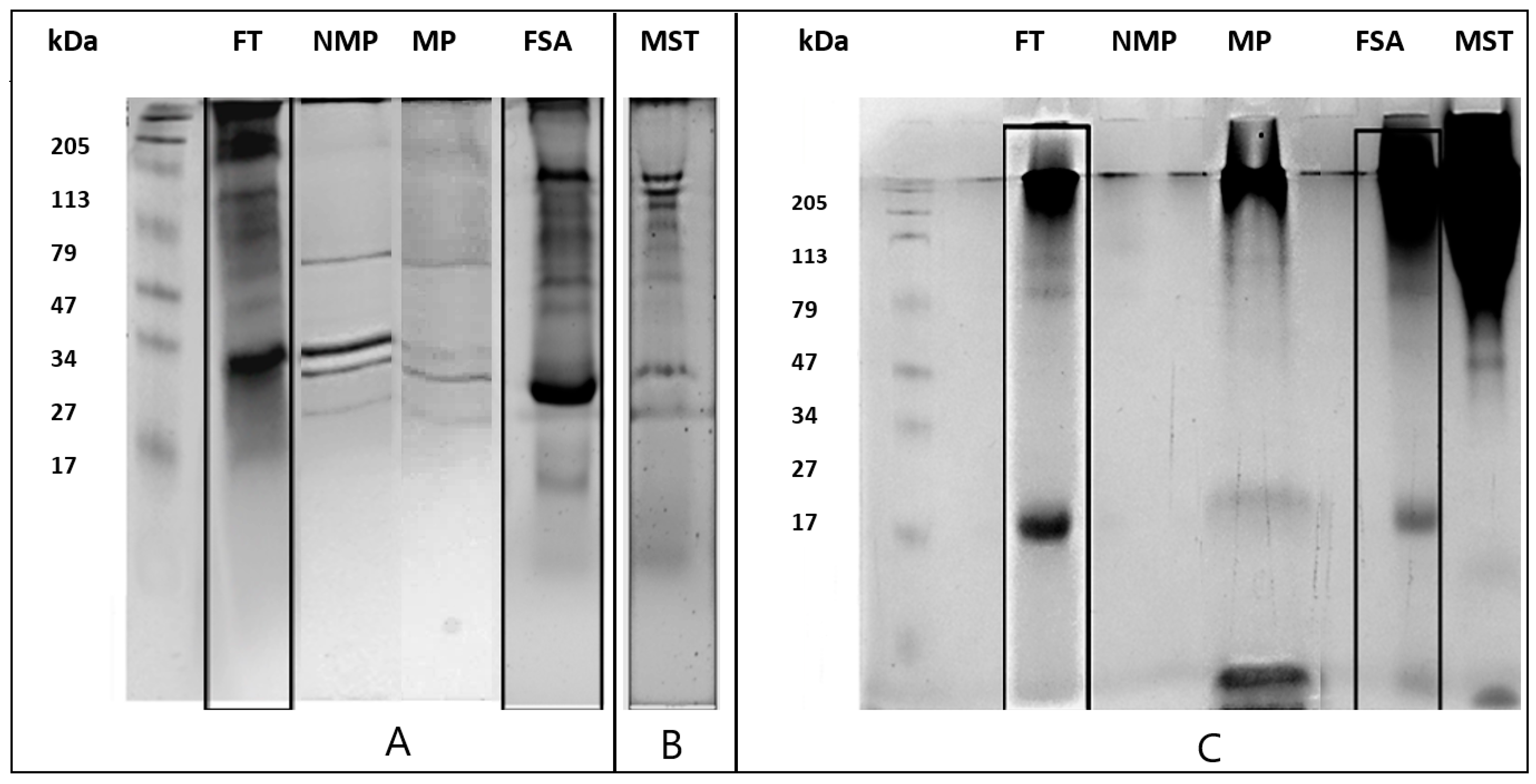
- Chardonnay white wine (500 mL) was dialyzed for 12 h at 4 °C against distilled water using a 3.5 kDa tubular membrane (Spectrum Chemical, New Brunswick, NJ, USA). Subsequently, the clear wine was frozen and freeze-dried. The powder obtained at the end of this process, containing the total wine macromolecules, was called the total fraction (FT).
- Chardonnay white wine (500 mL) was dialyzed as described in point 1. The dialyzed solution was then diluted 1:1 v/v with a saline buffer at pH 7.4 (Tris 20 mM, NaCl 0.5 M, CaCl2 1 mM, 1 mM MgCl2, and 1 mM MnCl2) and filtered with PTFE filters with MWCO 0.45 µm (Sartorius, Goettingen, Germany). Two distinct protein fractions were purified from this sample using the ÄKTApurifier affinity chromatography system (UP900 by Amersham, Chalfont St. Giles, UK). This system managed using the Unicorn 5.1 Software, mounted a column packed with 30 mL of Sepharose conjugated with Concanavalin A (Sigma-Aldrich, St. Louis, MO, USA). The sample was loaded via a 150 mL Superloop, and peaks were acquired at 214 nm. The first fraction collected corresponded to proteins having no affinity for Concanavalin A and, therefore, not mannosylated (NMP). Instead, for the elution of the mannosylated fraction (MP), a 0.5 M solution of Methyl-α-d-Mannopyranoside (Sigma-Aldrich, St. Louis, MO, USA) was used. Both fractions, MP and NMP, were subsequently dialyzed against water for 12 h at 4 °C with 3.5 kDa membranes and finally freeze-dried.
- Chardonnay white wine (500 mL) was subjected to precipitation with 60% (w/v) ammonium sulfate. The sample was then centrifuged at 14,000× g for 30 min, and the supernatant containing the ammonium sulfate-soluble protein fraction was dialyzed and lyophilized (FSA).
2.2. Protein Quantification
2.3. SDS-PAGE
2.4. Foaming Properties
2.5. Statistical Analysis
3. Result and Discussion
3.1. Protein Concentrations
3.2. SDS-PAGE Profile
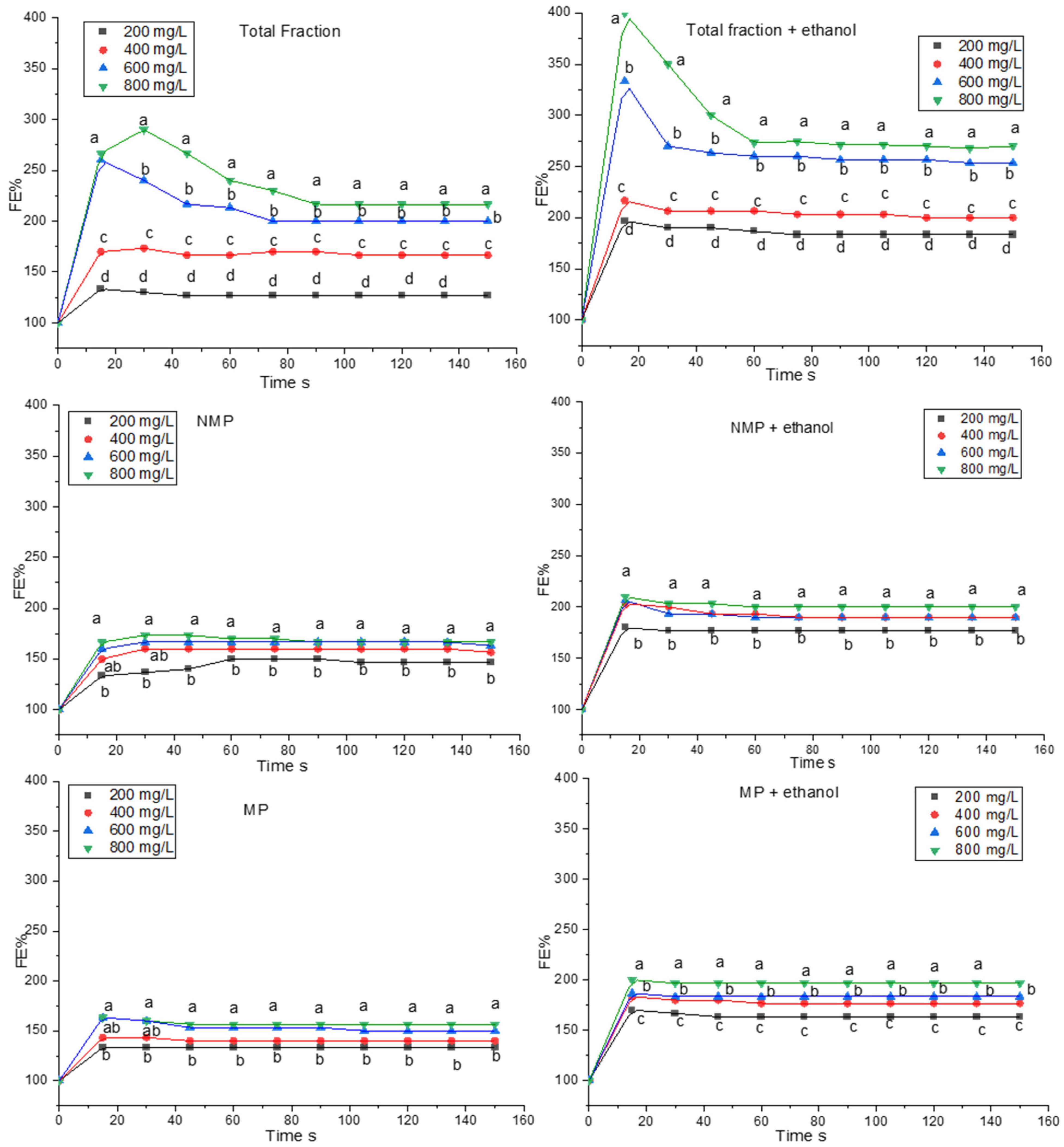
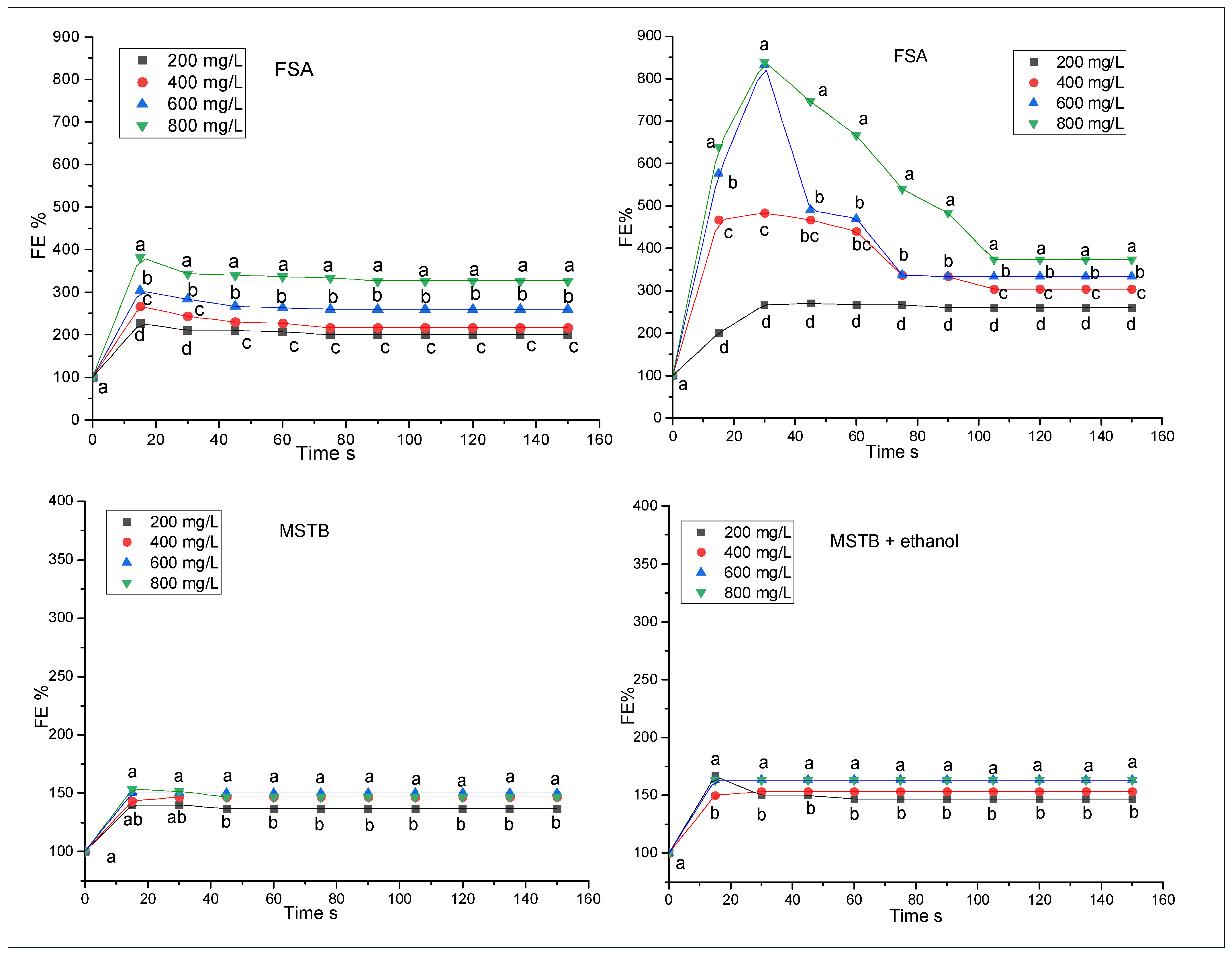
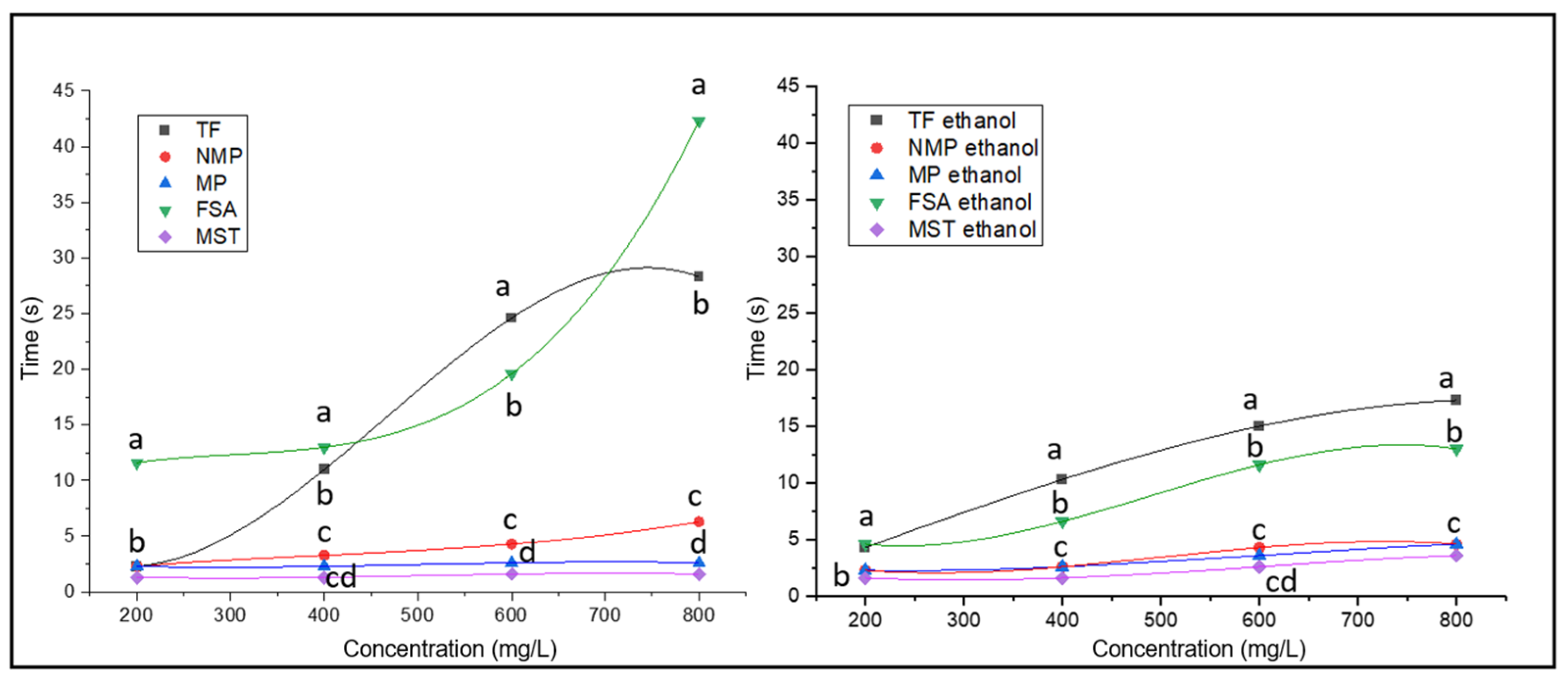
3.3. Foaming Properties
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liger-Belair, G.; Cilindre, C. Recent Progress in the Analytical Chemistry of Champagne and Sparkling Wines. Annu. Rev. Anal. Chem. 2021, 14, 21–46. [Google Scholar] [CrossRef]
- Kemp, B.; Condé, B.; Jégou, S.; Howell, K.; Vasserot, Y.; Marchal, R. Chemical Compounds and Mechanisms Involved in the Formation and Stabilization of Foam in Sparkling Wines. Crit. Rev. Food Sci. Nutr. 2019, 59, 2072–2094. [Google Scholar] [CrossRef] [PubMed]
- Schramm, L.L. Emulsions, Foams, Suspensions, and Aerosols: Microscience and Applications; John Wiley & Sons: Hoboken, NJ, USA, 2014; ISBN 978-3-527-33706-4. [Google Scholar]
- Huang, Y.; Li, J.; Liu, Y.; Gantumur, M.-A.; Sukhbaatar, N.; Zhao, P.; Oh, K.C.; Jiang, Z.; Hou, J. Improving Gas-Water Interface Properties and Bioactivities of α-Lactalbumin Induced by Three Structurally Different Saponins. Food Hydrocoll. 2023, 138, 108463. [Google Scholar] [CrossRef]
- Aguié-Béghin, V.; Adriaensen, Y.; Péron, N.; Valade, M.; Rouxhet, P.; Douillard, R. Structure and Chemical Composition of Layers Adsorbed at Interfaces with Champagne. J. Agric. Food Chem. 2009, 57, 10399–10407. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, Z.; Aguié-Béghin, V.; Abou-Saleh, K.; Douillard, R.; Bliard, C. Isolation and Analysis of Macromolecular Fractions Responsible for the Surface Properties in Native Champagne Wines. Food Res. Int. 2010, 43, 982–987. [Google Scholar] [CrossRef]
- Vilela, A.; Cosme, F.; Pinto, T. Emulsions, Foams, and Suspensions: The Microscience of the Beverage Industry. Beverages 2018, 4, 25. [Google Scholar] [CrossRef]
- Dupin, I.V.S.; Stockdale, V.J.; Williams, P.J.; Jones, G.P.; Markides, A.J.; Waters, E.J. Saccharomyces Cerevisiae Mannoproteins That Protect Wine from Protein Haze: Evaluation of Extraction Methods and Immunolocalization. J. Agric. Food Chem. 2000, 48, 1086–1095. [Google Scholar] [CrossRef] [PubMed]
- Cosme, F.; Fernandes, C.; Ribeiro, T.; Filipe-Ribeiro, L.; Nunes, F.M. White Wine Protein Instability: Mechanism, Quality Control and Technological Alternatives for Wine Stabilisation—An Overview. Beverages 2020, 6, 19. [Google Scholar] [CrossRef]
- Nunez, Y.P.; Carrascosa, A.V.; González, R.; Polo, M.C.; Martínez-Rodríguez, A.J. Effect of Accelerated Autolysis of Yeast on the Composition and Foaming Properties of Sparkling Wines Elaborated by a Champenoise Method. J. Agric. Food Chem. 2005, 53, 7232–7237. [Google Scholar] [CrossRef]
- Moreno-Arribas, M.V.; Polo, C. Wine Chemistry and Biochemistry; Springer Science & Business Media: Berlin, Germany, 2008; ISBN 978-0-387-74118-5. [Google Scholar]
- Tao, Y.; García, J.F.; Sun, D.-W. Advances in Wine Aging Technologies for Enhancing Wine Quality and Accelerating Wine Aging Process. Crit. Rev. Food Sci. Nutr. 2014, 54, 817–835. [Google Scholar] [CrossRef]
- Franceschi, D.; Lomolino, G.; Sato, R.; Vincenzi, S.; De Iseppi, A. Umami in Wine: Impact of Glutamate Concentration and Contact with Lees on the Sensory Profile of Italian White Wines. Beverages 2023, 9, 52. [Google Scholar] [CrossRef]
- Escot, S.; Feuillat, M.; Dulau, L.; Charpentier, C. Release of Polysaccharides by Yeasts and the Influence of Released Polysaccharides on Colour Stability and Wine Astringency. Aust. J. Grape Wine Res. 2001, 7, 153–159. [Google Scholar] [CrossRef]
- Péron, N.; Cagna, A.; Valade, M.; Marchal, R.; Maujean, A.; Robillard, B.; Aguié-Béghin, V.; Douillard, R. Characterisation by Drop Tensiometry and by Ellipsometry of the Adsorption Layer Formed at the Air/Champagne Wine Interface. Adv. Colloid. Interface Sci. 2000, 88, 19–36. [Google Scholar] [CrossRef]
- Puff, N.; Marchal, R.; Aguié-Béghin, V.; Douillard, R. Is Grape Invertase a Major Component of the Adsorption Layer Formed at the Air/Champagne Wine Interface? Langmuir 2001, 17, 2206–2212. [Google Scholar] [CrossRef]
- De Iseppi, A.; Marangon, M.; Vincenzi, S.; Lomolino, G.; Curioni, A.; Divol, B. A Novel Approach for the Valorization of Wine Lees as a Source of Compounds Able to Modify Wine Properties. LWT 2021, 136, 110274. [Google Scholar] [CrossRef]
- Vincenzi, S.; Crapisi, A.; Curioni, A. Foamability of Prosecco Wine: Cooperative Effects of High Molecular Weight Glycocompounds and Wine PR-Proteins. Food Hydrocoll. 2014, 34, 202–207. [Google Scholar] [CrossRef]
- Vanrell, G.; Canals, R.; Esteruelas, M.; Fort, F.; Canals, J.M.; Zamora, F. Influence of the Use of Bentonite as a Riddling Agent on Foam Quality and Protein Fraction of Sparkling Wines (Cava). Food Chem. 2007, 104, 148–155. [Google Scholar] [CrossRef]
- Mangindaan, D.; Khoiruddin, K.; Wenten, I.G. Beverage Dealcoholization Processes: Past, Present, and Future. Trends Food Sci. Technol. 2018, 71, 36–45. [Google Scholar] [CrossRef]
- Moine, V.L.; Dubourdieu, D. Biological Product for the Physico-Chemical Stabilization of a Wine. Patent Application No. FR2726284B1, 27 December 1996. [Google Scholar]
- Partsia, Z.; Kiosseoglou, V. Foaming Properties of Potato Proteins Recovered by Complexation with Carboxymethylcellulose. Colloids Surf. B Biointerfaces. 2001, 21, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Vincenzi, S.; Mosconi, S.; Zoccatelli, G.; Pellegrina, C.D.; Veneri, G.; Chignola, R.; Peruffo, A.; Curioni, A.; Rizzi, C. Development of a New Procedure for Protein Recovery and Quantification in Wine. Am. J. Enol. Vitic. 2005, 56, 182–187. [Google Scholar] [CrossRef]
- Wang, S.; Wang, X.; Zhao, P.; Ma, Z.; Zhao, Q.; Cao, X.; Cheng, C.; Liu, H.; Du, G. Mannoproteins Interfering Wine Astringency by Modulating the Reaction between Phenolic Fractions and Protein in a Model Wine System. LWT 2021, 152, 112217. [Google Scholar] [CrossRef]
- Li, S.; Zhai, H.; Ma, W.; Duan, C.; Yi, L. Yeast Mannoproteins: Organoleptic Modulating Functions, Mechanisms, and Product Development Trends in Winemaking. Food Front. 2023, 4, 1091–1126. [Google Scholar] [CrossRef]
- Vincenzi, S.; Marangon, M.; Tolin, S.; Curioni, A. Protein Evolution during the Early Stages of White Winemaking and Its Relations with Wine Stability. Aust. J. Grape Wine Res. 2011, 17, 20–27. [Google Scholar] [CrossRef]
- De Iseppi, A.; Curioni, A.; Marangon, M.; Vincenzi, S.; Kantureeva, G.; Lomolino, G. Characterization and Emulsifying Properties of Extracts Obtained by Physical and Enzymatic Methods from an Oenological Yeast Strain. J. Sci. Food Agric. 2019, 99, 5702–5710. [Google Scholar] [CrossRef] [PubMed]
- Jégou, S.; Conreux, A.; Villaume, S.; Hovasse, A.; Schaeffer, C.; Cilindre, C.; Van Dorsselaer, A.; Jeandet, P. One Step Purification of the Grape Vacuolar Invertase. Anal. Chim. Acta 2009, 638, 75–78. [Google Scholar] [CrossRef]
- Moine-Ledoux, V.; Dubourdieu, D. An Invertase Fragment Responsible for Improving the Protein Stability of Dry White Wines. J. Sci. Food Agric. 1999, 79, 537–543. [Google Scholar] [CrossRef]
- Snyman, C.; Mekoue Nguela, J.; Sieczkowski, N.; Divol, B.; Marangon, M. Characterization of Mannoprotein Structural Diversity in Wine Yeast Species. J. Agric. Food Chem. 2023, 71, 19727–19738. [Google Scholar] [CrossRef]
- Dussaud, A.; Robillard, B.; Carles, B.; Duteurtre, B.; Vignes-Adler, M. Exogenous Lipids and Ethanol Influences on the Foam Behavior of Sparkling Base Wines. J. Food Sci. 1994, 59, 148–151. [Google Scholar] [CrossRef]
- Brissonnet, F.; Maujean, A. Characterization of Foaming Proteins in a Champagne Base Wine. Am. J. Enol. Vitic. 1993, 44, 297–301. [Google Scholar] [CrossRef]
- De Iseppi, A.; Marangon, M.; Lomolino, G.; Crapisi, A.; Curioni, A. Red and White Wine Lees as a Novel Source of Emulsifiers and Foaming Agents. LWT 2021, 152, 112273. [Google Scholar] [CrossRef]
- Brierley, E.R.; Wilde, P.J.; Onishi, A.; Hughes, P.S.; Simpson, W.J.; Clark, D.C. The Influence of Ethanol on the Foaming Properties of Beer Protein Fractions: A Comparison of Rudin and Microconductivity Methods of Foam Assessment. J. Sci. Food Agric. 1996, 70, 531–537. [Google Scholar] [CrossRef]
- Ferreira, I.M.P.L.V.O.; Jorge, K.; Nogueira, L.C.; Silva, F.; Trugo, L.C. Effects of the Combination of Hydrophobic Polypeptides, Iso-α Acids, and Malto-Oligosaccharides on Beer Foam Stability. J. Agric. Food Chem. 2005, 53, 4976–4981. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fractions Extracted Protein | Weight Yield (Dry Weight, d.w.) mg/L | Total Protein mg/L | µg Protein/mg d.w. |
|---|---|---|---|
| Total Fraction (FT) | 152.3 ± 8.3 | 36.5 ± 2.3 | 239.6 ± 1.3 |
| Not mannosilated protein (NMP) | 114.3 ± 4.2 | 19.2 ± 0.9 | 167.9 ± 2.3 |
| Mannoprotein extracted by ConA (MP) | 29.4 ± 1.5 | 1.1 ± 0.15 | 37.4 ± 0.9 |
| soluble fraction in Sulfate ammonium (FSA) | 84.4 ± 1.7 | 24.2 ± 2.3 | 286.7 ± 3.4 |
| y = Intercept + B1 × x1 + B2 × x2 + B3 × x3 | |||||
|---|---|---|---|---|---|
| FSA | TF | MP | MST | NMP | |
| R2 | 1 | 1 | 1 | 1 | 1 |
| intercept | 4.5 ± 0.3 b | 13.03 ± 0.2 a | 3.2 ± 0.4 c | 2.2 ± 0.08 d | 0.3 ± 0.002 e |
| B1 | 0.06792 a | −0.128 d | −0.0077 c | −0.0077 c | 0.0141 b |
| B2 | −2.08 × 10−4 c | 4.31 × 10−4 a | 1.88 × 10−5 b | 1.88 × 10−5 b | −2.50 × 10−5 d |
| B3 | 2.27 × 10−7 a | −3.08 × 10−7 d | −1.25 × 10−8 c | −1.25 × 10−8 c | 2.08 × 10−8 b |
| FSA ethanol | FT ethanol | MP ethanol | MST ethanol | NMP ethanol | |
| R2 | 1 | 1 | 1 | 1 | 1 |
| intercept | 12.2 ± 1.1 a | −1.9 ± 0.3 d | 3.4 ± 0.4 c | 3.6 ± 0.3 c | 6.2 ± 0.5 b |
| B1 | −0.073 d | 2.90 × 10−2 a | −0.01 b | −0.01 b | −0.034 c |
| B2 | 2.02 × 10−4 a | 1.13 × 10−5 d | 2.63 × 10−5 c | 3.75 × 10−5 c | 8.75 × 10−5 b |
| B3 | −1.37 × 10−7 a | −2.29 × 10−8 b | −1.40 × 10−8 b | −2.08 × 10−8 b | −5.83 × 10−8 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lomolino, G.; Vincenzi, S.; Zannoni, S.; Vegro, M.; De Iseppi, A. White Sparkling Wine Proteins and Glycoproteins and Their Behavior in Foam Expansion and Stability. Beverages 2024, 10, 21. https://doi.org/10.3390/beverages10010021
Lomolino G, Vincenzi S, Zannoni S, Vegro M, De Iseppi A. White Sparkling Wine Proteins and Glycoproteins and Their Behavior in Foam Expansion and Stability. Beverages. 2024; 10(1):21. https://doi.org/10.3390/beverages10010021
Chicago/Turabian StyleLomolino, Giovanna, Simone Vincenzi, Stefania Zannoni, Mara Vegro, and Alberto De Iseppi. 2024. "White Sparkling Wine Proteins and Glycoproteins and Their Behavior in Foam Expansion and Stability" Beverages 10, no. 1: 21. https://doi.org/10.3390/beverages10010021