Mechanics and Microstructure of the Atrioventricular Heart Valve Chordae Tendineae: A Review


Abstract
:1. Introduction
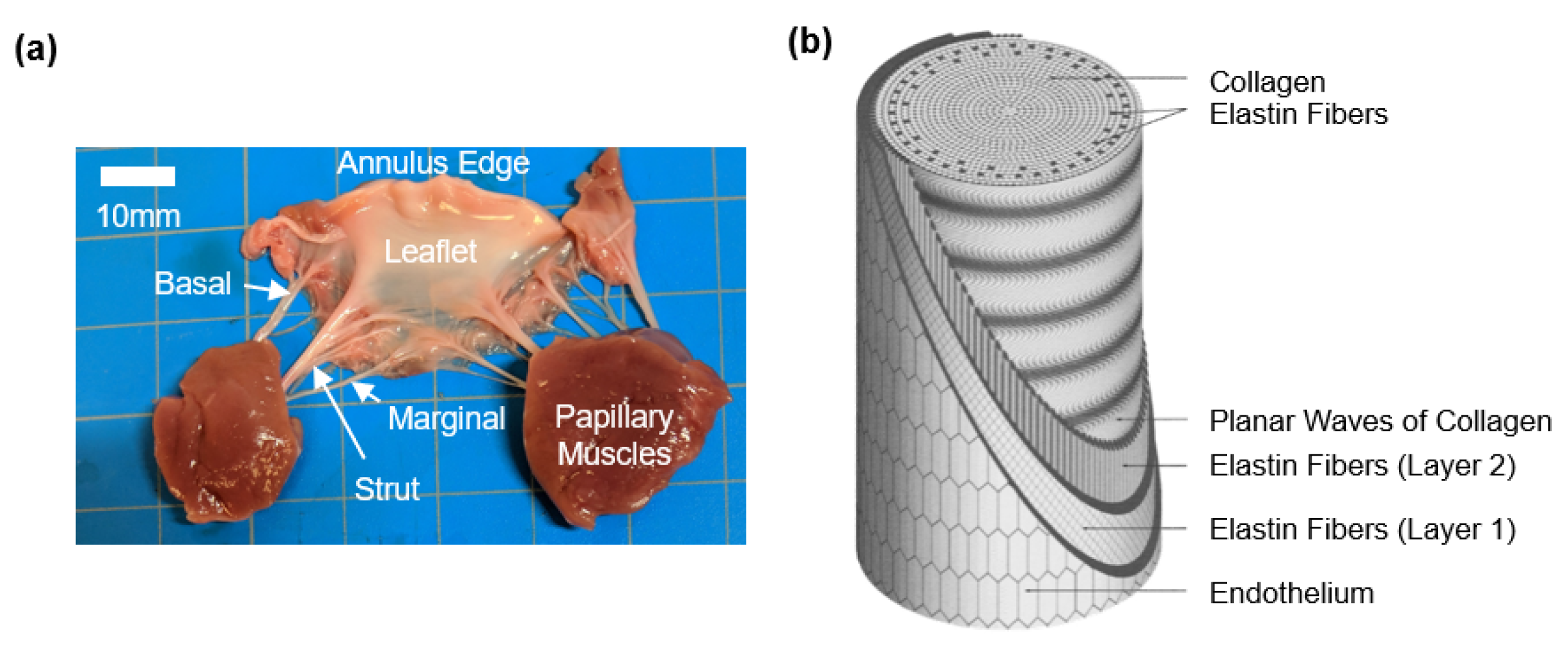
2. Overview on the Anatomy and Morphology of the AHV Chordae Tendineae
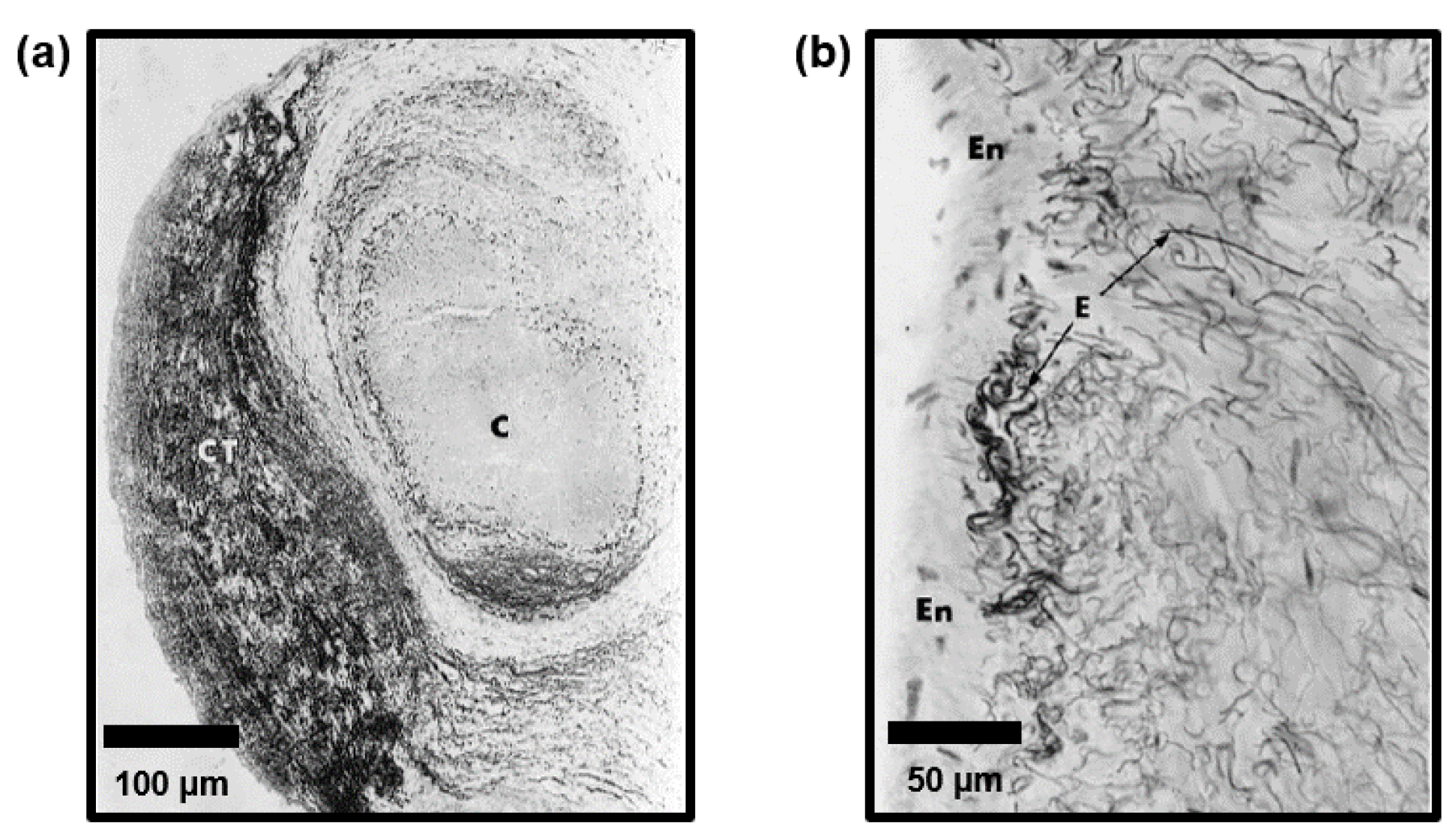
3. Chordae Tendineae Microstructure
3.1. Microstructures of Human AHV Chordae Tendineae
3.2. Effects of Disease on Human AHV Chordae Tendineae Microstructure
3.3. Comparisons of the Chordae Microstructures Between Different Species
3.4. Comparisons of the Microstructures Between Chordae Subsets
3.5. Microstructures of the Chordae Insertion Regions
3.6. Microstructures of the Artificial Chordae
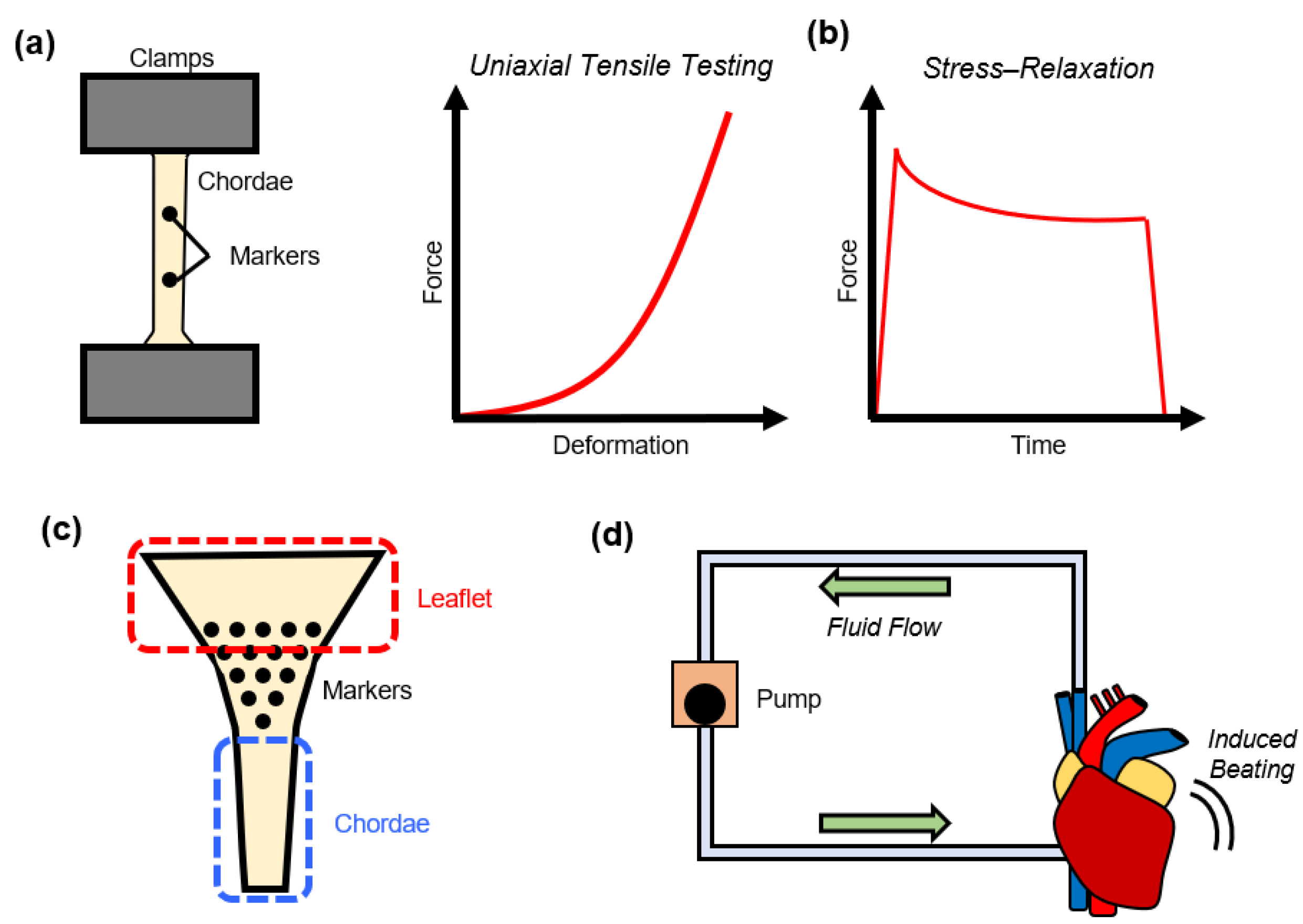
4. Tissue Mechanics of the Chordae Tendineae
4.1. Uniaxial Mechanical Testing of the Chordae Tendineae
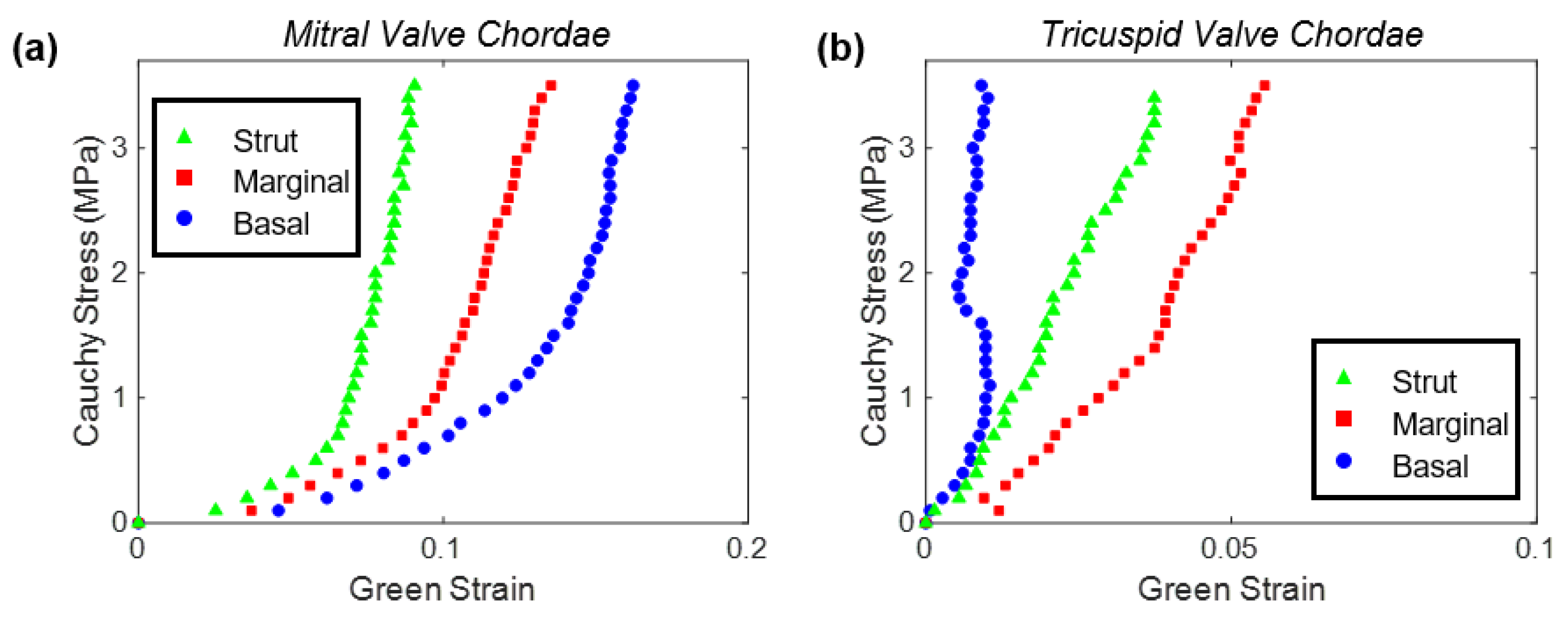
4.1.1. Uniaxial Tensile Characterizations of Human Chordae Tendineae
4.1.2. Effects of Disease on the Tensile Characteristics of Human Chordae Tendineae
4.1.3. Mechanical Characterizations of Porcine Chordae Tendineae
4.2. Stress-Relaxation Testing of the Chordae Tendineae
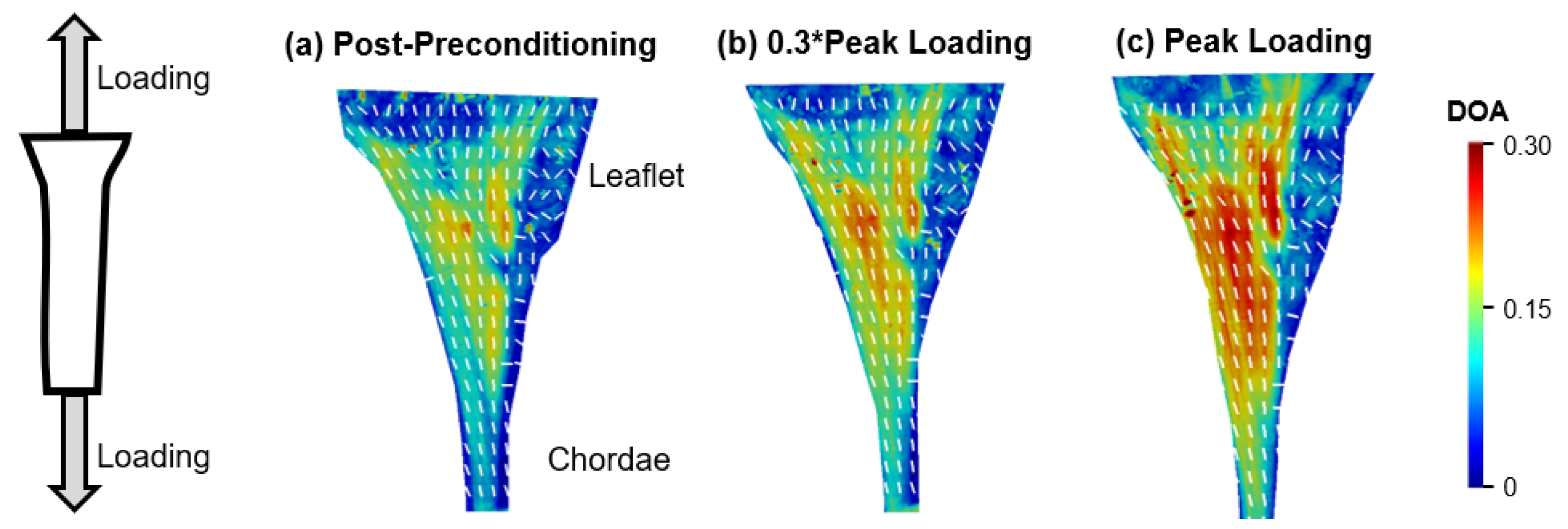
4.3. Load-Dependent Collagen Fiber Architecture of the Chordae-Leaflet Insertion Region
4.4. In Vitro Flow Loop Testing of Chordae Tendineae
4.5. Mechanics of Artificial Chordae
5. Closing Remarks and Future Prospects
Author Contributions
Funding
Conflicts of Interest
References
- Waller, B.F.; Howard, J.; Fess, S. Pathology of mitral valve stenosis and pure mitral regurgitation—Part I. Clin. Cardiol. 1994, 17, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Waller, B.F.; Howard, J.; Fess, S. Pathology of tricuspid valve stenosis and pure tricuspid regurgitation—Part I. Clin. Cardiol. 1995, 18, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Sanders, C.A.; Austen, W.G.; Harthorne, J.W.; Dinsmore, R.E.; Scannell, J.G. Diagnosis and surgical treatment of mitral regurgitation secondary to ruptured chordae tendineae. N. Engl. J. Med. 1967, 276, 943–949. [Google Scholar] [CrossRef] [PubMed]
- Orszulak, T.A.; Schaff, H.V.; Danielson, G.K.; Piehler, J.M.; Pluth, J.R.; Frye, R.L.; McGoon, D.C.; Elveback, L.R. Mitral regurgitation due to ruptured chordae tendineae: Early and late results of valve repair. J. Thorac. Cardiovasc. Surg. 1985, 89, 491–498. [Google Scholar] [CrossRef]
- Jeresaty, R.M.; Edwards, J.E.; Chawla, S.K. Mitral valve prolapse and ruptured chordae tendineae. Am. J. Cardiol. 1985, 55, 138–142. [Google Scholar] [CrossRef]
- Khoiy, K.A.; Asgarian, K.T.; Loth, F.; Amini, R. Dilation of tricuspid valve annulus immediately after rupture of chordae tendineae in ex-vivo porcine hearts. PLoS ONE 2018, 13, e0206744. [Google Scholar] [CrossRef]
- Espino, D.M.; Hukins, D.W.; Shepherd, D.E.; Buchan, K.G. Mitral valve repair: An in-vitro comparison of the effect of surgical repair on the pressure required to cause mitral valve regurgitation. J. Heart. Valve Dis. 2006, 15, 375. [Google Scholar]
- Carpentier, A.; Adams, D.H.; Filsoufi, F. Carpentier’s Reconstructive Valve Surgery From Valve Analysis to Valve Reconstruction; Saunders Elsevier Health Sciences: Philadelphia, PA, USA, 2011. [Google Scholar]
- Smedira, N.G.; Selman, R.; Cosgrove, D.M.; McCarthy, P.M.; Lytle, B.W.; Taylor, P.C.; Apperson-Hansen, C.; Stewart, R.W.; Loop, F.D. Repair of anterior leaflet prolapse: Chordal transfer is superior to chordal shortening. J. Thorac. Cardiovasc. Surg. 1996, 112, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Phillips, M.R.; Daly, R.C.; Schaff, H.V.; Dearani, J.A.; Mullany, C.J.; Orszulak, T.A. Repair of anterior leaflet mitral valve prolapse: Chordal replacement versus chordal shortening. Ann. Thorac. Surg. 2000, 69, 25–29. [Google Scholar] [CrossRef]
- Sousa, U.M.; Grare, P.; Jebara, V.; Fuzelier, J.F.; Portoghese, M.; Acar, C.; Relland, J.; Mihaileanu, S.; Fabiani, J.N.; Carpentier, A. Transposition of chordae in mitral valve repair. Mid-term results. Circulation 1993, 88, II35–II38. [Google Scholar]
- Revuelta, J.M.; Garcia-Rinaldi, R.; Gaite, L.; Val, F.; Garijo, F. Generation of chordae tendineae with polytetrafluoroethylene stents: Results of mitral valve chordal replacement in sheep. J. Thorac. Cardiovasc. Surg. 1989, 97, 98–103. [Google Scholar] [CrossRef]
- Tabata, M.; Kasegawa, H.; Fukui, T.; Shimizu, A.; Sato, Y.; Takanashi, S. Long-term outcomes of artificial chordal replacement with tourniquet technique in mitral valve repair: A single-center experience of 700 cases. J. Thorac. Cardiovasc. Surg. 2014, 148, 2033–2038.e2031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cochran, R.P.; Kuzelman, K.S. Comparison of viscoelastic properties of suture versus porcine mitral valve chordae tendineae. J. Card. Surg. 1991, 6, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Colli, A.; Manzan, E.; Rucinskas, K.; Janusauskas, V.; Zucchetta, F.; Zakarkaitė, D.; Aidietis, A.; Gerosa, G. Acute safety and efficacy of the NeoChord procedure. Interact. Cardiovasc. Thorac. Surg. 2015, 20, 575–581. [Google Scholar] [CrossRef] [Green Version]
- Butany, J.; Collins, M.J.; David, T.E. Ruptured synthetic expanded polytetrafluoroethylene chordae tendinae. Cardiovasc. Pathol. 2004, 13, 182–184. [Google Scholar] [CrossRef]
- Gunnal, S.A.; Wabale, R.N.; Farooqui, M.S. Morphological study of chordae tendinae in human cadaveric hearts. Heart Views 2015, 16, 1–12. [Google Scholar] [CrossRef]
- Lam, J.H.C.; Ranganathan, N.; Wigle, E.D.; Silver, M.D. Morphology of the human mitral valve: I. Chordae tendineae: A new classification. Circulation 1970, 41, 449–458. [Google Scholar] [CrossRef] [Green Version]
- Silver, M.D.; Lam, J.H.C.; Ranganathan, N.; Wigle, E.D. Morphology of the human tricuspid valve. Circulation 1971, 43, 333–348. [Google Scholar] [CrossRef] [Green Version]
- Lomholt, M.; Nielsen, S.L.; Hansen, S.B.; Andersen, N.T.; Hasenkam, J.M. Differential tension between secondary and primary mitral chordae in an acute in-vivo porcine model. J. Heart. Valve Dis. 2002, 11, 337–345. [Google Scholar]
- Zuo, K.; Pham, T.; Li, K.; Martin, C.; He, Z.; Sun, W. Characterization of biomechanical properties of aged human and ovine mitral valve chordae tendineae. J. Mech. Behav. Biomed. Mater. 2016, 62, 607–618. [Google Scholar] [CrossRef] [Green Version]
- Millington-Sanders, C.; Meir, A.; Lawrence, L.; Stolinski, C. Structure of chordae tendineae in the left ventricle of the human heart. J. Anat. 1998, 192, 573–581. [Google Scholar] [CrossRef] [PubMed]
- De Almeida, M.T.B.; Aragao, I.C.S.A.; Aragao, F.M.S.A.; Reis, F.P.; Aragao, J.A. Morphological study on mitral valve chordae tendineae in the hearts of human fetuses. Int. J. Anat. Var. 2019, 12, 17–20. [Google Scholar]
- Jett, S.V.; Laurence, D.W.; Kunkel, R.P.; Babu, A.R.; Kramer, K.E.; Baumwart, R.; Towner, R.A.; Wu, Y.; Lee, C.-H. An investigation of the anisotropic mechanical properties and anatomical structure of porcine atrioventricular heart valves. J. Mech. Behav. Biomed. Mater. 2018, 87, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Fenoglio, J.J., Jr.; Pham, T.D.; Wit, A.L.; Bassett, A.L.; Wagner, B.M. Canine mitral complex: Ultrastructure and electromechanical properties. Circul. Res. 1972, 31, 417–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, K.O.; Boughner, D.R. Scanning electron microscopical study of human mitral valve chordae tendineae. Arch. Pathol. Lab. Med. 1977, 101, 236–238. [Google Scholar]
- Lim, K.O. Mechanical properties and ultrastructure of normal human tricuspid valve chordae tendineae. Jpn. J. Physiol. 1980, 30, 455–464. [Google Scholar] [CrossRef] [Green Version]
- Grande-Allen, K.J.; Griffin, B.P.; Ratliff, N.B.; Cosgrove, D.M.; Vesely, I. Glycosaminoglycan profiles of myxomatous mitral leaflets and chordae parallel the severity of mechanical alterations. J. Am. Coll. Cardiol. 2003, 42, 271–277. [Google Scholar] [CrossRef] [Green Version]
- Baker, P.B.; Bansal, G.; Boudoulas, H.; Kolibash, A.J.; Kilman, J.; Wooley, C.F. Floppy mitral valve chordae tendineae: Histopathologic alterations. Hum. Pathol. 1988, 19, 507–512. [Google Scholar] [CrossRef]
- Hollander, W. Unified concept on the role of acid mucopolysaccharides and connective tissue proteins in the accumulation of lipids, lipoproteins, and calcium in the atherosclerotic plaque. Exp. Mol. Pathol. 1976, 25, 106–120. [Google Scholar] [CrossRef]
- Fornes, P.; Heudes, D.; Fuzellier, J.-F.; Tixier, D.; Bruneval, P.; Carpentier, A. Correlation between clinical and histologic patterns of degenerative mitral valve insufficiency: A histomorphometric study of 130 excised segments. Cardiovasc. Pathol. 1999, 8, 81–92. [Google Scholar] [CrossRef]
- Tamura, K.; Fukuda, Y.; Ishizaki, M.; Masuda, Y.; Yamanaka, N.; Ferrans, V.J. Abnormalities in elastic fibers and other connective-tissue components of floppy mitral valve. Am. Heart J. 1995, 129, 1149–1158. [Google Scholar] [CrossRef]
- Lis, Y.; Burleigh, M.C.; Parker, D.J.; Child, A.H.; Hogg, J.; Davies, M.J. Biochemical characterization of individual normal, floppy and rheumatic human mitral valves. Biochem. J. 1987, 244, 597–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Icardo, J.M.; Colvee, E.; Revuelta, J.M. Structural analysis of chordae tendineae in degenerative disease of the mitral valve. Int. J. Cardiol. 2013, 167, 1603–1609. [Google Scholar] [CrossRef]
- Thein, E.; Hammer, C. Physiologic barriers to xenotransplantation. Curr. Opin. Organ. Transplant. 2004, 9, 186–189. [Google Scholar] [CrossRef]
- Crick, S.J.; Sheppard, M.N.; Ho, S.Y.; Gebstein, L.; Anderson, R.H. Anatomy of the pig heart: Comparisons with normal human cardiac structure. J. Anat. 1998, 193, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, J.; Warnock, J.N.; Yoganathan, A.P. Structural characterization of the chordae tendineae in native porcine mitral valves. Ann. Thorac. Surg. 2005, 80, 189–197. [Google Scholar] [CrossRef]
- Duran, C.M.G.; Gunning, A.J. The vascularization of the heart valves: A comparative study. Cardiovasc. Res. 1968, 2, 290–296. [Google Scholar] [CrossRef]
- Liao, J.; Priddy, L.B.; Wang, B.; Chen, J.; Vesely, I. Ultrastructure of porcine mitral valve chordae tendineae. J. Heart. Valve Dis. 2009, 18, 292. [Google Scholar]
- Liao, J.; Vesely, I. A structural basis for the size-related mechanical properties of mitral valve chordae tendineae. J. Biomech. 2003, 36, 1125–1133. [Google Scholar] [CrossRef]
- Liao, J.; Vesely, I. Relationship between collagen fibrils, glycosaminoglycans, and stress relaxation in mitral valve chordae tendineae. Ann. Biomed. Eng. 2004, 32, 977–983. [Google Scholar] [CrossRef]
- Chen, L.; Yin, F.C.P.; May-Newman, K. The structure and mechanical properties of the mitral valve leaflet-strut chordae transition zone. J. Biomech. Eng. 2004, 126, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Gusukuma, L.W.; Prates, J.C.; Smith, R.L.; Gusukuma, W.L.; Prates, J.C.; Smith, R.L. Chordae tendineae architecture in the papillary muscle insertion. Int. J. Morphol. 2004, 22, 267–272. [Google Scholar] [CrossRef] [Green Version]
- Salvador, L.; Mirone, S.; Bianchini, R.; Regesta, T.; Patelli, F.; Minniti, G.; Masat, M.; Cavarretta, E.; Valfrè, C. A 20-year experience with mitral valve repair with artificial chordae in 608 patients. J. Thorac. Cardiovasc. Surg. 2008, 135, 1280–1287.e1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minatoya, K.; Kobayashi, J.; Sasako, Y.; Ishibashi-Ueda, H.; Yutani, C.; Kitamura, S. Long-term pathological changes of expanded polytetrafluoroethylene (ePTFE) suture in the human heart. J. Heart. Valve Dis. 2001, 10, 139–142. [Google Scholar]
- Coutinho, G.F.; Carvalho, L.; Antunes, M.J. Acute mitral regurgitation due to ruptured ePTFE neo-chordae. J. Heart Valve Dis. 2007, 16, 278–281. [Google Scholar] [PubMed]
- Lim, K.O.; Boughner, D.R. Mechanical properties of human mitral valve chordae tendineae: Variation with size and strain rate. Can. J. Physiol. Pharmacol. 1975, 53, 330–339. [Google Scholar] [CrossRef]
- Yamada, H.; Evans, F.G. Strength of Biological Materials; Williams & Wilkins: Baltimore, MD, USA, 1970. [Google Scholar]
- Lim, K.O.; Boughner, D.R. Morphology and relationship to extensibility curves of human mitral valve chordae tendineae. Circul. Res. 1976, 39, 580–585. [Google Scholar] [CrossRef] [Green Version]
- Clark, R.E. Stress-strain characteristics of fresh and frozen human aortic and mitral leaflets and chordae tendineae. Implications for clinical use. J. Thorac. Cardiovasc. Surg. 1973, 66, 202–208. [Google Scholar] [CrossRef]
- Barber, J.E.; Ratliff, N.B.; Cosgrove, D.M., 3rd; Griffin, B.P.; Vesely, I. Myxomatous mitral valve chordae. I: Mechanical properties. J. Heart. Valve Dis. 2001, 10, 320–324. [Google Scholar]
- Lim, K.O.; Boughner, D.R.; Perkins, D.G. Ultrastructure and mechanical properties of chordae tendineae from a myxomatous tricuspid valve. Jpn. Heart J. 1983, 24, 539–548. [Google Scholar] [CrossRef] [Green Version]
- Casado, J.A.; Diego, S.; Ferreño, D.; Ruiz, E.; Carrascal, I.; Méndez, D.; Revuelta, J.M.; Pontón, A.; Icardo, J.M.; Gutiérrez-Solana, F. Determination of the mechanical properties of normal and calcified human mitral chordae tendineae. J. Mech. Behav. Biomed. Mater. 2012, 13, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritchie, J.; Jimenez, J.; He, Z.; Sacks, M.S.; Yoganathan, A.P. The material properties of the native porcine mitral valve chordae tendineae: An in vitro investigation. J. Biomech. 2006, 39, 1129–1135. [Google Scholar] [CrossRef] [PubMed]
- Pokutta-Paskaleva, A.; Sulejmani, F.; DelRocini, M.; Sun, W. Comparative mechanical, morphological, and microstructural characterization of porcine mitral and tricuspid leaflets and chordae tendineae. Acta Biomater. 2019, 85, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Sedransk, K.L.; Grande-Allen, K.J.; Vesely, I. Failure mechanics of mitral valve chordae tendineae. J. Heart. Valve Dis. 2002, 11, 644–650. [Google Scholar] [PubMed]
- Gunning, G.M.; Murphy, B.P. The effects of decellularization and cross-linking techniques on the fatigue life and calcification of mitral valve chordae tendineae. J. Mech. Behav. Biomed. Mater. 2016, 57, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Gunning, G.M.; Murphy, B.P. Characterisation of the fatigue life, dynamic creep and modes of damage accumulation within mitral valve chordae tendineae. Acta Biomater. 2015, 24, 193–200. [Google Scholar] [CrossRef]
- Constable, M.; Burton, H.E.; Lawless, B.M.; Gramigna, V.; Buchan, K.G.; Espino, D.M. Effect of glutaraldehyde based cross-linking on the viscoelasticity of mitral valve basal chordae tendineae. Biomed. Eng. 2018, 17, 93. [Google Scholar] [CrossRef] [Green Version]
- Ross, C.J.; Laurence, D.W.; Hsu, M.-C.; Baumwart, R.; Zhao, D.Y.; Mir, A.; Burkhart, H.M.; Holzapfel, G.A.; Wu, Y.; Lee, C.-H. Mechanics of porcine heart valves’ strut chordae tendineae investigated as a leaflet-chordae-papillary muscle entity. Ann. Biomed. Eng. 2020. [Google Scholar] [CrossRef]
- Madhurapantula, R.S.; Krell, G.; Morfin, B.; Roy, R.; Lister, K.; Orgel, J.P. Advanced methodology and preliminary measurements of molecular and mechanical properties of heart valves under dynamic strain. Int. J. Mol. Sci. 2020, 21, 763. [Google Scholar] [CrossRef] [Green Version]
- Kunzelman, K.S.; Cochran, K.P. Mechanical properties of basal and marginal mitral valve chordae tendineae. ASAIO J. 1990, 36, M405–M407. [Google Scholar]
- Ross, C.J.; Laurence, D.W.; Richardson, J.; Babu, A.R.; Evans, L.E.; Beyer, E.G.; Wu, Y.; Towner, R.A.; Fung, K.-M.; Mir, A.; et al. An investigation of the glycosaminoglycan contribution to biaxial mechanical behaviors of porcine atrioventricular heart valve leaflets. J. R. Soc. Interface 2019, 16, 20190069. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, V.K.; Grande-Allen, K.J. The role of proteoglycans and glycosaminoglycans in heart valve biomechanics. In Advances in Heart Valve Biomechanics; Springer: Cham, Switzerland, 2018; pp. 59–79. [Google Scholar]
- Padala, M.; Sacks, M.S.; Liou, S.W.; Balachandran, K.; He, Z.; Yoganathan, A.P. Mechanics of the mitral valve strut chordae insertion region. J. Biomech. Eng. 2010, 132, 081004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jett, S.V.; Hudson, L.T.; Baumwart, R.; Bohnstedt, B.N.; Mir, A.; Burkhart, H.M.; Holzapfel, G.A.; Wu, Y.; Lee, C.-H. Integration of polarized spatial frequency domain imaging (pSFDI) with a biaxial mechanical testing system for quantification of load-dependent collagen architecture in soft collagenous tissues. Acta Biomater. 2020, 102, 149–168. [Google Scholar] [CrossRef] [PubMed]
- Goth, W.; Potter, S.; Allen, A.C.B.; Zoldan, J.; Sacks, M.S.; Tunnell, J.W. Non-destructive reflectance mapping of collagen fiber alignment in heart valve leaflets. Ann. Biomed. Eng. 2019, 47, 1250–1264. [Google Scholar] [CrossRef] [PubMed]
- Caimmi, P.P.; Sabbatini, M.; Fusaro, L.; Borrone, A.; Cannas, M. A study of the mechanical properties of ePTFE suture used as artificial mitral chordae. J. Card. Surg. 2016, 31, 498–502. [Google Scholar] [CrossRef]
- Goth, W.; Yang, B.; Lesicko, J.; Allen, A.; Sacks, M.S.; Tunnell, J.W. Polarized spatial frequency domain imaging of heart valve fiber structure. In Proceedings of SPIE, Optical Elastography and Tissue Biomechanics III, San Francisco, CA, USA, 24 June 2016; Volume 9710, p. 971019. [Google Scholar]
- Arts, T.; Meerbaum, S.; Reneman, R.; Corday, E. Stresses in the closed mitral valve: A model study. J. Biomech. 1983, 16, 539–547. [Google Scholar] [CrossRef]
- McQueen, D.M.; Peskin, C.S.; Yellin, E.L. Fluid dynamics of the mitral valve: Physiological aspects of a mathematical model. Am. J. Physiol. Heart Circ. Physiol. 1982, 242, H1095–H1110. [Google Scholar] [CrossRef]
- Lim, K.H.; Yeo, J.H.; Duran, C. Three-dimensional asymmetrical modeling of the mitral valve: A finite element study with dynamic boundaries. J. Heart. Valve Dis. 2005, 14, 386–392. [Google Scholar]
- Wang, Q.; Sun, W. Finite element modeling of mitral valve dynamic deformation using patient-specific multi-slices computed tomography scans. Ann. Biomed. Eng. 2013, 41, 142–153. [Google Scholar] [CrossRef]
- Kunzelman, K.; Reimink, M.S.; Verrier, E.D.; Cochran, R.P. Replacement of mitral valve posterior chordae tendineae with expanded polytetrafluoroethylene suture: A finite element study. J. Card. Surg. 1996, 11, 136–145. [Google Scholar] [CrossRef]
- Wenk, J.F.; Ratcliffe, M.B.; Guccione, J.M. Finite element modeling of mitral leaflet tissue using a layered shell approximation. Med. Biol Eng. Comput 2012, 50, 1071–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalighi, A.H.; Drach, A.; Bloodworth, C.H.; Pierce, E.L.; Yoganathan, A.P.; Gorman, R.C.; Gorman, J.H., III; Sacks, M.S. Mitral valve chordae tendineae: Topological and geometrical characterization. Ann. Biomed. Eng. 2017, 45, 378–393. [Google Scholar] [CrossRef] [PubMed]
- Meador, W.D.; Mathur, M.; Sugerman, G.P.; Jazwiec, T.; Malinowski, M.; Bersi, M.R.; Timek, T.A.; Rausch, M.K. A detailed mechanical and microstructural analysis of ovine tricuspid valve leaflets. Acta Biomater. 2020, 102, 100–113. [Google Scholar] [CrossRef] [PubMed]
- Khalighi, A.H.; Rego, B.V.; Drach, A.; Gorman, R.C.; Gorman, J.H., III; Sacks, M.S. Development of a functionally equivalent model of the mitral valve chordae tendineae through topology optimization. Ann. Biomed. Eng. 2019, 47, 60–74. [Google Scholar] [CrossRef]
- Rim, Y.; Laing, S.T.; McPherson, D.D.; Kim, H. Mitral valve repair using ePTFE sutures for ruptured mitral chordae tendineae: A computational simulation study. Ann. Biomed. Eng. 2014, 42, 139–148. [Google Scholar] [CrossRef]
- Aggarwal, A.; Aguilar, V.S.; Lee, C.-H.; Ferrari, G.; Gorman, J.H., III; Gorman, R.C.; Sacks, M.S. Patient-specific modeling of heart valves: From image to simulation. In Proceedings of the International Conference on Functional Imaging and Modeling of the Heart, London, UK, 20–22 June 2013; Sébastien, O., Rueckert, D., Smith, N., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 141–149. [Google Scholar]
- Ionasec, R.I. Patient-Specific Modeling and Quantification of the Heart Valves from Multimodal Cardiac Images; Technische Universität München: Munich, Germany, 2010. [Google Scholar]
- Siefert, A.W.; Rabbah, J.-P.M.; Pierce, E.L.; Kunzelman, K.S.; Yoganathan, A.P. Quantitative evaluation of annuloplasty on mitral valve chordae tendineae forces to supplement surgical planning model development. Cardiovasc. Eng. Technol. 2014, 5, 35–43. [Google Scholar] [CrossRef]
- Stevanella, M.; Votta, E.; Lemma, M.; Antona, C.; Redaelli, A. Finite element modelling of the tricuspid valve: A preliminary study. Med. Eng. Phys. 2010, 32, 1213–1223. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MVAL Strut | TVAL Strut | ||||
| Study | Species | Tissue Stretch λ (−) | Cauchy Stress (MPa) | Tissue Stretch λ (−) | Cauchy Stress (MPa) |
| Pokutta-Paskaleva et al. (2019) [55] | porcine (n = not provided) | 1.09 | 3.5 | 1.04 | 3.5 |
| Ritchie et al. (2006) [54] | porcine (n = not provided) | 1.05 | 0.89 to 1.18 | – | – |
| Liao and Vesely (2003) [40] | porcine (n = 16) | 1.16 ± 0.03 (mean ± SD) | 0.75 ± 0.15 (mean ± SD) | – | – |
| Zuo et al. (2016) [21] | ovine (n = 18) | 1.07 ± 0.08 (mean ± SD) | 24 (mean ± SD) | – | – |
| Ross et al. (2020) [60] | porcine (n = 12) | 1.03 ± 0.01 (mean ± SEM) | 1.59 ± 0.16 (mean ± SEM) | 1.02 ± 0.01 (mean ± SEM) | 2.71 ± 0.10 (mean ± SEM) |
| MVAL Marginal | TVAL Marginal | ||||
| Pokutta-Paskaleva et al. (2019) [55] | porcine (n = not provided) | 1.13 | 3.5 | 1.05 | 3.5 |
| Kunzelman and Cochran (1990) [62] | porcine (n = 31) | 1.09 | 1.96 ± 0.20 (mean ± SEM) | – | – |
| Liao and Vesely (2003) [40] | porcine (n = 16) | 1.04 ± 0.01 (mean ± SD) | 5.22 ± 3.30 (mean ± SD) | – | – |
| MVAL Basal | TVAL Basal | ||||
| Pokutta-Paskaleva et al. (2019) [55] | porcine (n = not provided) | 1.15 | 3.5 | 1.01 | 3.5 |
| Kunzelman and Cochran (1990) [62] | porcine (n = 29) | 1.12 | 1.57 ± 0.05 (mean ± SEM) | – | – |
| Liao and Vesely (2003) [40] | porcine (n = 20) | 1.08 ± 0.03 (mean ± SD) | 2.41 ± 0.81 (mean ± SD) | – | – |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ross, C.J.; Zheng, J.; Ma, L.; Wu, Y.; Lee, C.-H. Mechanics and Microstructure of the Atrioventricular Heart Valve Chordae Tendineae: A Review. Bioengineering 2020, 7, 25. https://doi.org/10.3390/bioengineering7010025
Ross CJ, Zheng J, Ma L, Wu Y, Lee C-H. Mechanics and Microstructure of the Atrioventricular Heart Valve Chordae Tendineae: A Review. Bioengineering. 2020; 7(1):25. https://doi.org/10.3390/bioengineering7010025
Chicago/Turabian StyleRoss, Colton J., Junnan Zheng, Liang Ma, Yi Wu, and Chung-Hao Lee. 2020. "Mechanics and Microstructure of the Atrioventricular Heart Valve Chordae Tendineae: A Review" Bioengineering 7, no. 1: 25. https://doi.org/10.3390/bioengineering7010025