

Spiking Neuron Mathematical Models: A Compact Overview


Abstract
:1. Introduction
2. Modeling Spiking Neurons and Their Dynamical Behaviors
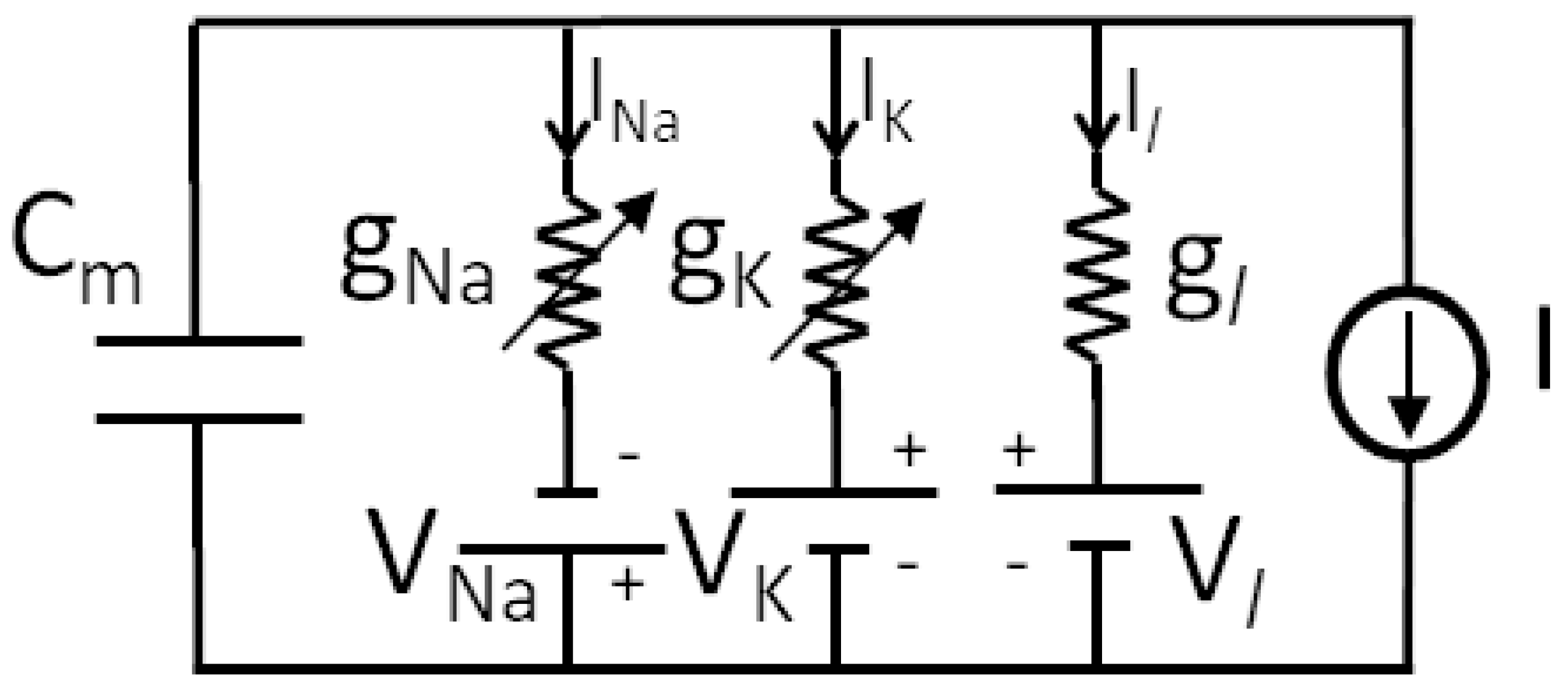
2.1. Genesis of Neuron Modeling
2.2. Mathematical Models and Bifurcations
3. Compact Overview of Recent Literature
4. Focus on the Recent Bibliography on Spiking Systems
4.1. Spiking Neuron Parameter Estimation
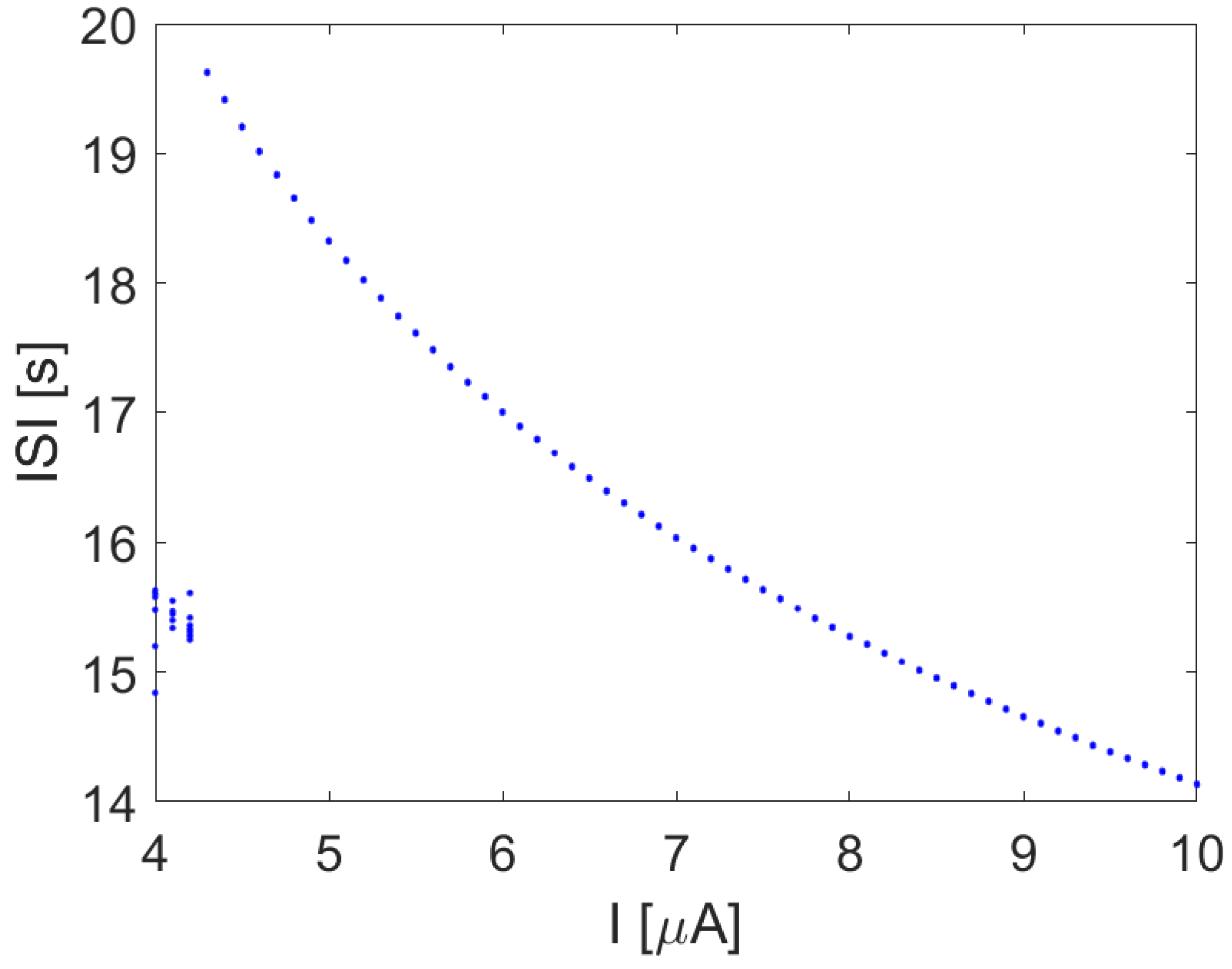
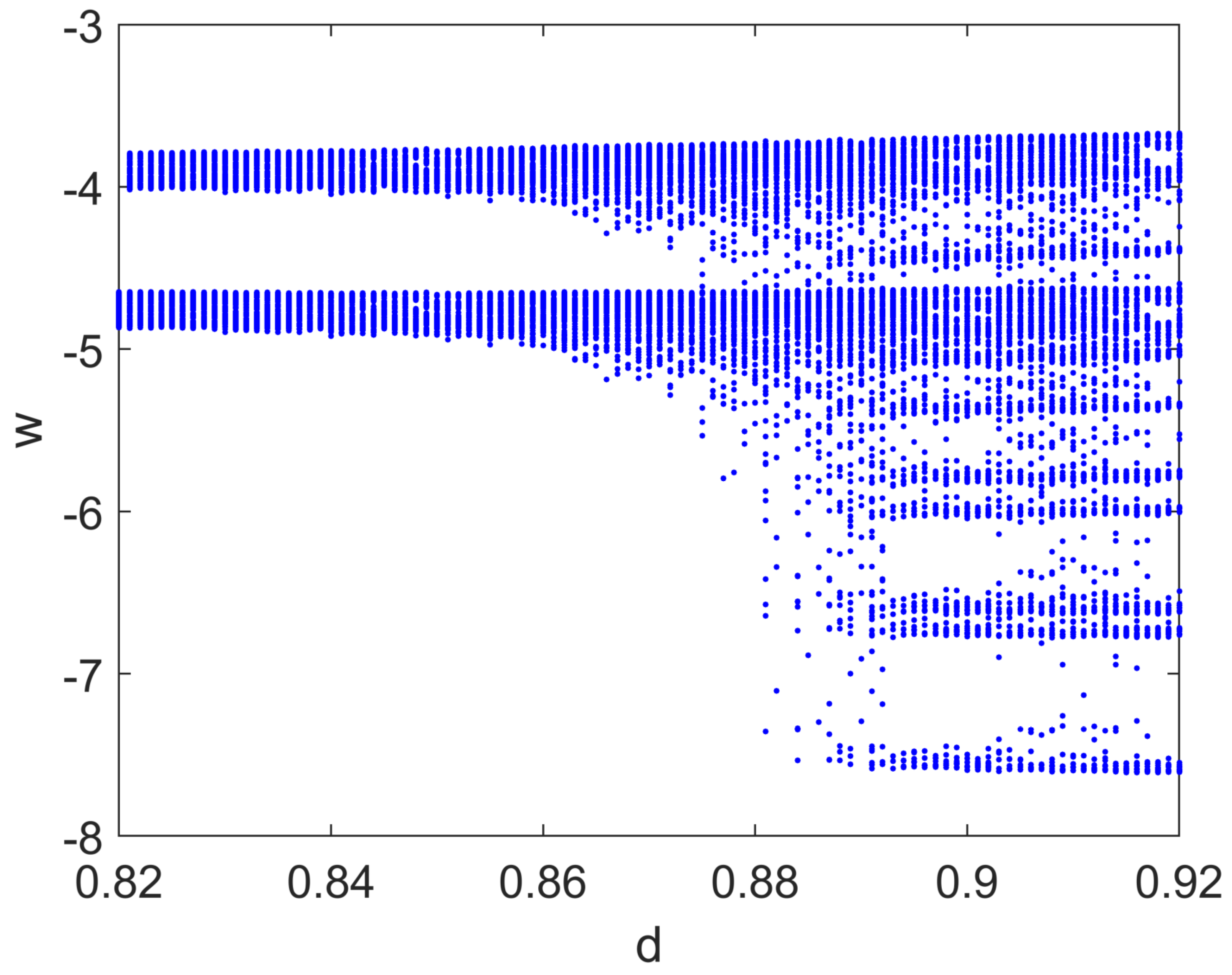
4.2. Focus on Bifurcation and Chaos in Spiking Neurons
4.3. Synchronization of Spiking Neurons
4.4. Focus on Stochasticity and Noise in Spiking Neurons
4.5. Focus on Non-Integer Order Spiking Neurons
4.6. Focus on Spiking Neurons and Memristors
- The state equations are nonlinear , ;
- They have bipole memories, called ReRAM;
- They are prone to creating neural synaptic memories, as shown by Crupi et al. [67]
4.7. Focus on the Implementation of Spiking Neurons
5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Fortuna, L.; Buscarino, A.; Frasca, M.; Famoso, C. Control of Imperfect Nonlinear Electromechanical Large Scale Systems: From Dynamics to Hardware Implementation; World Scientific: Singapore, 2017; Volume 91. [Google Scholar]
- Rössler, O.E. An equation for continuous chaos. Phys. Lett. A 1976, 57, 397–398. [Google Scholar] [CrossRef]
- Grzesiak, L.M.; Meganck, V. Spiking signal processing: Principle and applications in control system. Neurocomputing 2018, 308, 31–48. [Google Scholar] [CrossRef]
- Lapique, L. Recherches quantitatives sur l’excitation electrique des nerfs traitee comme une polarization. J. Physiol. Pathol. 1907, 9, 620–635. [Google Scholar]
- Abbott, L.F. Lapicque’s introduction of the integrate-and-fire model neuron (1907). Brain Res. Bull. 1999, 50, 303–304. [Google Scholar] [CrossRef] [PubMed]
- Brunel, N.; Van Rossum, M.C. Lapicque’s 1907 paper: From frogs to integrate-and-fire. Biol. Cybern. 2007, 97, 337–339. [Google Scholar] [CrossRef] [PubMed]
- Nernst, W. Reasoning of Theoretical Chemistry: Nine Papers (1889–1921); Verlag Harri Deutsch: Frankfurt am Main, Germany, 2003. [Google Scholar]
- Hodgkin, A.L.; Huxley, A.F. A quantitative description of membrane current and its application to conduction and excitation in nerve. J. Physiol. 1952, 117, 500. [Google Scholar] [CrossRef] [PubMed]
- FitzHugh, R. Impulses and physiological states in theoretical models of nerve membrane. Biophys. J. 1961, 1, 445–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hindmarsh, J.L.; Rose, R. A model of neuronal bursting using three coupled first order differential equations. Proc. R. Soc. London. Ser. B. Biol. Sci. 1984, 221, 87–102. [Google Scholar]
- Joshi, S.K. Synchronization of Coupled Hindmarsh-Rose Neuronal Dynamics: Analysis and Experiments. IEEE Trans. Circuits Syst. II: Express Briefs 2021, 69, 1737–1741. [Google Scholar] [CrossRef]
- La Rosa, M.; Rabinovich, M.; Huerta, R.; Abarbanel, H.; Fortuna, L. Slow regularization through chaotic oscillation transfer in an unidirectional chain of Hindmarsh–Rose models. Phys. Lett. A 2000, 266, 88–93. [Google Scholar] [CrossRef]
- Izhikevich, E.M. Simple model of spiking neurons. IEEE Trans. Neural Networks 2003, 14, 1569–1572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, C.; Lecar, H. Voltage oscillations in the barnacle giant muscle fiber. Biophys. J. 1981, 35, 193–213. [Google Scholar] [CrossRef]
- Tsumoto, K.; Kitajima, H.; Yoshinaga, T.; Aihara, K.; Kawakami, H. Bifurcations in Morris–Lecar neuron model. Neurocomputing 2006, 69, 293–316. [Google Scholar] [CrossRef]
- Buscarino, A.; Fortuna, L.; Frasca, M. Essentials of Nonlinear Circuit Dynamics with MATLAB® and Laboratory Experiments; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Buscarino, A.; Fortuna, L.; Frasca, M.; Gambuzza, L.V.; Sciuto, G. Memristive chaotic circuits based on cellular nonlinear networks. Int. J. Bifurc. Chaos 2012, 22, 1250070. [Google Scholar] [CrossRef]
- Rodríguez-Collado, A.; Rueda, C. A simple parametric representation of the Hodgkin-Huxley model. PLoS ONE 2021, 16, e0254152. [Google Scholar] [CrossRef] [PubMed]
- Doruk, R.O.; Abosharb, L. Estimating the parameters of FitzHugh–nagumo neurons from neural spiking data. Brain Sci. 2019, 9, 364. [Google Scholar] [CrossRef] [PubMed]
- Fradkov, A.L.; Kovalchukov, A.; Andrievsky, B. Parameter Estimation for Hindmarsh–Rose Neurons. Electronics 2022, 11, 885. [Google Scholar] [CrossRef]
- Jauberthie, C.; Verdière, N. Bounded-Error Parameter Estimation Using Integro-Differential Equations for Hindmarsh–Rose Model. Algorithms 2022, 15, 179. [Google Scholar] [CrossRef]
- Baysal, V.; Yılmaz, E. Chaotic signal induced delay decay in Hodgkin-Huxley Neuron. Appl. Math. Comput. 2021, 411, 126540. [Google Scholar] [CrossRef]
- Li, L.; Zhao, Z. White-noise-induced double coherence resonances in reduced Hodgkin-Huxley neuron model near subcritical Hopf bifurcation. Phys. Rev. E 2022, 105, 034408. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Feng, R.; Rui, Z.; Zhang, A. Effects of time delay on chaotic neuronal discharges. Math. Comput. Appl. 2010, 15, 840–845. [Google Scholar] [CrossRef] [Green Version]
- Tlelo-Cuautle, E.; Díaz-Mu noz, J.D.; González-Zapata, A.M.; Li, R.; León-Salas, W.D.; Fernández, F.V.; Guillén-Fernández, O.; Cruz-Vega, I. Chaotic image encryption using hopfield and hindmarsh–rose neurons implemented on FPGA. Sensors 2020, 20, 1326. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, Y.; Tsushima, H.; Ikeguchi, T. Non-periodic responses of the Izhikevich neuron model to periodic inputs. Nonlinear Theory Its Appl. IEICE 2022, 13, 367–372. [Google Scholar] [CrossRef]
- Xue, Y.; Jiang, J.; Hong, L. A LSTM based prediction model for nonlinear dynamical systems with chaotic itinerancy. Int. J. Dyn. Control 2020, 8, 1117–1128. [Google Scholar] [CrossRef]
- Pal, K.; Ghosh, D.; Gangopadhyay, G. Synchronization and metabolic energy consumption in stochastic Hodgkin-Huxley neurons: Patch size and drug blockers. Neurocomputing 2021, 422, 222–234. [Google Scholar] [CrossRef]
- Adomaitienė, E.; Bumelienė, S.; Tamaševičius, A. Controlling synchrony in an array of the globally coupled FitzHugh–Nagumo type oscillators. Phys. Lett. A 2022, 431, 127989. [Google Scholar] [CrossRef]
- Hussain, I.; Jafari, S.; Ghosh, D.; Perc, M. Synchronization and chimeras in a network of photosensitive FitzHugh–Nagumo neurons. Nonlinear Dyn. 2021, 104, 2711–2721. [Google Scholar] [CrossRef]
- Plotnikov, S.A.; Fradkov, A.L. On synchronization in heterogeneous FitzHugh–Nagumo networks. Chaos Solitons Fractals 2019, 121, 85–91. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, G.; Li, X.; Wang, D. The synchronization behaviors of coupled fractional-order neuronal networks under electromagnetic radiation. Symmetry 2021, 13, 2204. [Google Scholar] [CrossRef]
- Rehák, B.; Lynnyk, V. Synchronization of a network composed of stochastic Hindmarsh–Rose neurons. Mathematics 2021, 9, 2625. [Google Scholar] [CrossRef]
- Vivekanandhan, G.; Hamarash, I.I.; Ali Ali, A.M.; He, S.; Sun, K. Firing patterns of Izhikevich neuron model under electric field and its synchronization patterns. Eur. Phys. J. Spec. Top. 2022, 231, 1–7. [Google Scholar] [CrossRef]
- Margarit, D.H.; Reale, M.V.; Scagliotti, A.F. Analysis of a signal transmission in a pair of Izhikevich coupled neurons. Biophys. Rev. Lett. 2020, 15, 195–206. [Google Scholar] [CrossRef]
- Li, T.; Wang, G.; Yu, D.; Ding, Q.; Jia, Y. Synchronization mode transitions induced by chaos in modified Morris–Lecar neural systems with weak coupling. Nonlinear Dyn. 2022, 108, 2611–2625. [Google Scholar] [CrossRef]
- Camps, O.; Stavrinides, S.G.; De Benito, C.; Picos, R. Implementation of the Hindmarsh–Rose Model Using Stochastic Computing. Mathematics 2022, 10, 4628. [Google Scholar] [CrossRef]
- Cai, R.; Liu, Y.; Duan, J.; Abebe, A.T. State transitions in the Morris-Lecar model under stable Lévy noise. Eur. Phys. J. B 2020, 93, 1–9. [Google Scholar] [CrossRef]
- Beaubois, R.; Khoyratee, F.; Branchereau, P.; Ikeuchi, Y.; Levi, T. From real-time single to multicompartmental Hodgkin-Huxley neurons on FPGA for bio-hybrid systems. In Proceedings of the 2022 44th Annual International Conference of the IEEE Engineering in Medicine & Biology Society (EMBC), Glasgow, Scotland, 11–15 July 2022; pp. 1602–1606. [Google Scholar]
- Stoliar, P.; Schneegans, O.; Rozenberg, M.J. Biologically relevant dynamical behaviors realized in an ultra-compact neuron model. Front. Neurosci. 2020, 14, 421. [Google Scholar] [CrossRef]
- Leigh, A.J.; Mirhassani, M.; Muscedere, R. An efficient spiking neuron hardware system based on the hardware-oriented modified Izhikevich neuron (HOMIN) model. IEEE Trans. Circuits Syst. II: Express Briefs 2020, 67, 3377–3381. [Google Scholar] [CrossRef]
- Karaca, Z.; Korkmaz, N.; Altuncu, Y.; Kılıç, R. An extensive FPGA-based realization study about the Izhikevich neurons and their bio-inspired applications. Nonlinear Dyn. 2021, 105, 3529–3549. [Google Scholar] [CrossRef]
- Alkabaa, A.S.; Taylan, O.; Yilmaz, M.T.; Nazemi, E.; Kalmoun, E.M. An Investigation on Spiking Neural Networks Based on the Izhikevich Neuronal Model: Spiking Processing and Hardware Approach. Mathematics 2022, 10, 612. [Google Scholar] [CrossRef]
- Ghiasi, A.; Zahedi, A. Field-programmable gate arrays-based Morris-Lecar implementation using multiplierless digital approach and new divider-exponential modules. Comput. Electr. Eng. 2022, 99, 107771. [Google Scholar] [CrossRef]
- Takaloo, H.; Ahmadi, A.; Ahmadi, M. Design and Analysis of the Morris-Lecar Spiking Neuron in Efficient Analog Implementation. IEEE Trans. Circuits Syst. II Express Briefs 2022, 70, 6–10. [Google Scholar] [CrossRef]
- Alfaqeih, S.; Mısırlı, E. On convergence analysis and analytical solutions of the conformable fractional FitzHugh–nagumo model using the conformable sumudu decomposition method. Symmetry 2021, 13, 243. [Google Scholar] [CrossRef]
- Angstmann, C.N.; Henry, B.I. Time Fractional Fisher–KPP and FitzHugh–Nagumo Equations. Entropy 2020, 22, 1035. [Google Scholar] [CrossRef]
- Azizi, T. Analysis of Neuronal Oscillations of Fractional-Order Morris-Lecar Model. Eur. J. Math. Anal 2023, 3, 2. [Google Scholar] [CrossRef]
- Chua, L. Memristor, Hodgkin–Huxley, and edge of chaos. Nanotechnology 2013, 24, 383001. [Google Scholar] [CrossRef]
- Huang, H.M.; Yang, R.; Tan, Z.H.; He, H.K.; Zhou, W.; Xiong, J.; Guo, X. Quasi-Hodgkin–Huxley Neurons with Leaky Integrate-and-Fire Functions Physically Realized with Memristive Devices. Adv. Mater. 2019, 31, 1803849. [Google Scholar] [CrossRef]
- Bao, H.; Liu, W.; Chen, M. Hidden extreme multistability and dimensionality reduction analysis for an improved non-autonomous memristive FitzHugh–Nagumo circuit. Nonlinear Dyn. 2019, 96, 1879–1894. [Google Scholar] [CrossRef]
- Xu, W.; Wang, J.; Yan, X. Advances in memristor-based neural networks. Front. Nanotechnol. 2021, 3, 645995. [Google Scholar] [CrossRef]
- Qi, G.; Wu, Y.; Hu, J. Abundant Firing Patterns in a Memristive Morris–Lecar Neuron Model. Int. J. Bifurc. Chaos 2021, 31, 2150170. [Google Scholar] [CrossRef]
- Zheng, C.; Peng, L.; Eshraghian, J.K.; Wang, X.; Cen, J.; Iu, H.H.C. Spiking Neuron Implementation Using a Novel Floating Memcapacitor Emulator. Int. J. Bifurc. Chaos 2022, 32, 2250224. [Google Scholar] [CrossRef]
- Chen, S.; Billings, S.A. Neural networks for nonlinear dynamic system modeling and identification. Int. J. Control 1992, 56, 319–346. [Google Scholar] [CrossRef]
- Arena, P.; Fazzino, S.; Fortuna, L.; Maniscalco, P. Game theory and non-linear dynamics: The Parrondo Paradox case study. Chaos Solitons Fractals 2003, 17, 545. [Google Scholar] [CrossRef]
- Hayashi, H.; Ishizuka, S.; Ohta, M.; Hirakawa, K. Chaotic behavior in the onchidium giant neuron under sinusoidal stimulation. Phys. Lett. A 1982, 88, 435–438. [Google Scholar] [CrossRef]
- Bucolo, M.; Buscarino, A.; Famoso, C.; Fortuna, L.; Frasca, M. Control of imperfect dynamical systems. Nonlinear Dyn. 2019, 98, 2989–2999. [Google Scholar] [CrossRef]
- Camps, O.; Stavrinides, S.G.; Picos, R. Stochastic computing implementation of chaotic systems. Mathematics 2021, 9, 375. [Google Scholar] [CrossRef]
- Gaines, B.R. Stochastic computing systems. In Advances in Information Systems Science; Springer: Berlin/Heidelberg, Germany, 1969; pp. 37–172. [Google Scholar]
- Alaghi, A.; Hayes, J.P. Survey of stochastic computing. ACM Trans. Embed. Comput. Syst. (TECS) 2013, 12, 1–19. [Google Scholar] [CrossRef]
- Chua, L. Memristor-the missing circuit element. IEEE Trans. Circuit Theory 1971, 18, 507–519. [Google Scholar] [CrossRef]
- Williams, R.S. How we found the missing memristor. IEEE Spectr. 2008, 45, 28–35. [Google Scholar] [CrossRef]
- Mehonic, A.; Shluger, A.L.; Gao, D.; Valov, I.; Miranda, E.; Ielmini, D.; Bricalli, A.; Ambrosi, E.; Li, C.; Yang, J.J.; et al. Silicon oxide (SiOx): A promising material for resistance switching? Adv. Mater. 2018, 30, 1801187. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Liu, G.; Wang, C.; Zhang, W.; Li, R.W.; Wang, L. Polymer memristor for information storage and neuromorphic applications. Mater. Horizons 2014, 1, 489–506. [Google Scholar] [CrossRef]
- Campbell, K.A. The Self-directed Channel Memristor: Operational Dependence on the Metal-Chalcogenide Layer. In Handbook of Memristor Networks; Springer: Berlin/Heidelberg, Germany, 2019; pp. 815–842. [Google Scholar]
- Crupi, M.; Pradhan, L.; Tozer, S. modeling neural plasticity with memristors. IEEE Can. Rev. 2012, 68, 10–14. [Google Scholar]
- Besrour, M.; Zitoun, S.; Lavoie, J.; Omrani, T.; Koua, K.; Benhouria, M.; Boukadoum, M.; Fontaine, R. Analog Spiking Neuron in 28 nm CMOS. In Proceedings of the 2022 20th IEEE Interregional NEWCAS Conference (NEWCAS), Quebec City, QC, Canada, 19–22 June 2022; pp. 148–152. [Google Scholar]
- Torti, E.; Florimbi, G.; Dorici, A.; Danese, G.; Leporati, F. Towards the Simulation of a Realistic Large-Scale Spiking Network on a Desktop Multi-GPU System. Bioengineering 2022, 9, 543. [Google Scholar] [CrossRef] [PubMed]
- Baden, T.; James, B.; Zimmermann, M.J.; Bartel, P.; Grijseels, D.; Euler, T.; Lagnado, L.; Maravall, M. Spikeling: A low-cost hardware implementation of a spiking neuron for neuroscience teaching and outreach. PLoS Biol. 2018, 16, e2006760. [Google Scholar] [CrossRef] [PubMed]
- Branciforte, M.; Buscarino, A.; Fortuna, L. A Hyperneuron Model Towards in Silico Implementation. Int. J. Bifurc. Chaos 2022, 32, 2250202. [Google Scholar] [CrossRef]
- Arena, P.; Bucolo, M.; Buscarino, A.; Fortuna, L.; Frasca, M. Reviewing bioinspired technologies for future trends: A complex systems point of view. Front. Phys. 2021, 9, 750090. [Google Scholar] [CrossRef]
- Sepulchre, R. Spiking control systems. Proc. IEEE 2022, 110, 577–589. [Google Scholar] [CrossRef]
- Mead, C. Neuromorphic electronic systems. Proc. IEEE 1990, 78, 1629–1636. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Kumar, A.; Parkash, C.; Vashishtha, G.; Tang, H.; Xiang, J. A novel entropy-based sparsity measure for prognosis of bearing defects and development of a sparsogram to select sensitive filtering band of an axial piston pump. Measurement 2022, 203, 111997. [Google Scholar] [CrossRef]
- Mead, C. How we created neuromorphic engineering. Nat. Electron. 2020, 3, 434–435. [Google Scholar] [CrossRef]





















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HH | FHN | HR | Izhikevich | ML | |
|---|---|---|---|---|---|
| Parameter Estimation | [18] | [19] | [20,21] | ||
| Bifurcation and Chaos | [22,23] | [24,25] | [26] | [27] | |
| Synchronization | [28] | [29,30,31] | [32,33] | [34,35] | [36] |
| Stochasticity and Noise | [23,28] | [33,37] | [38] | ||
| Implementations | [39] | [25,37] | [40,41,42,43] | [44,45] | |
| Non-integer order | [46,47] | [32] | [48] | ||
| Memristors | [49,50] | [51,52] | [53,54] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fortuna, L.; Buscarino, A. Spiking Neuron Mathematical Models: A Compact Overview. Bioengineering 2023, 10, 174. https://doi.org/10.3390/bioengineering10020174
Fortuna L, Buscarino A. Spiking Neuron Mathematical Models: A Compact Overview. Bioengineering. 2023; 10(2):174. https://doi.org/10.3390/bioengineering10020174
Chicago/Turabian StyleFortuna, Luigi, and Arturo Buscarino. 2023. "Spiking Neuron Mathematical Models: A Compact Overview" Bioengineering 10, no. 2: 174. https://doi.org/10.3390/bioengineering10020174