Identification of the Components in a Vaccinium oldhamii Extract Showing Inhibitory Activity against Influenza Virus Adsorption



Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimens
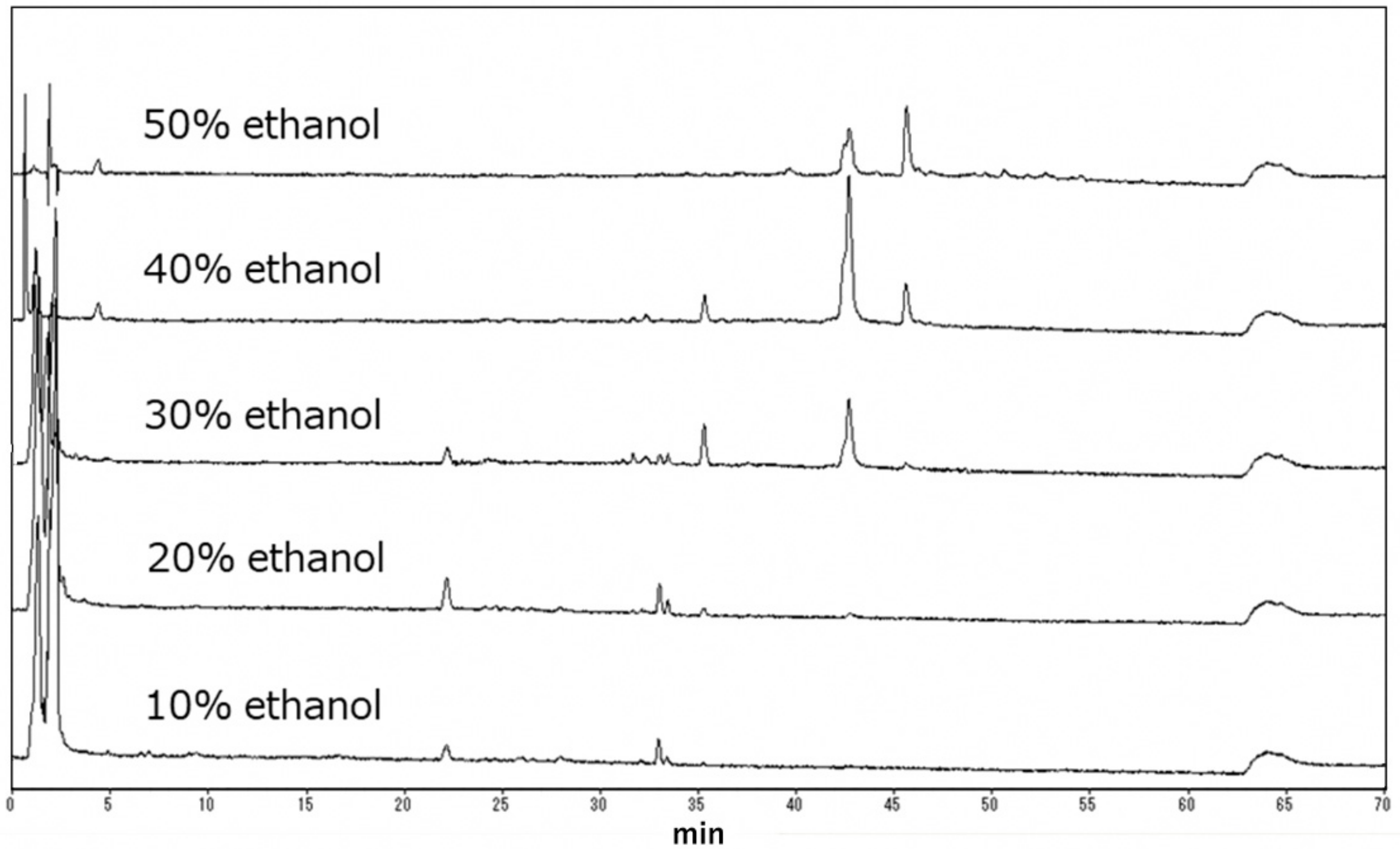
2.2. Separation of Vaccinium oldhamii by Synthetic Absorbent
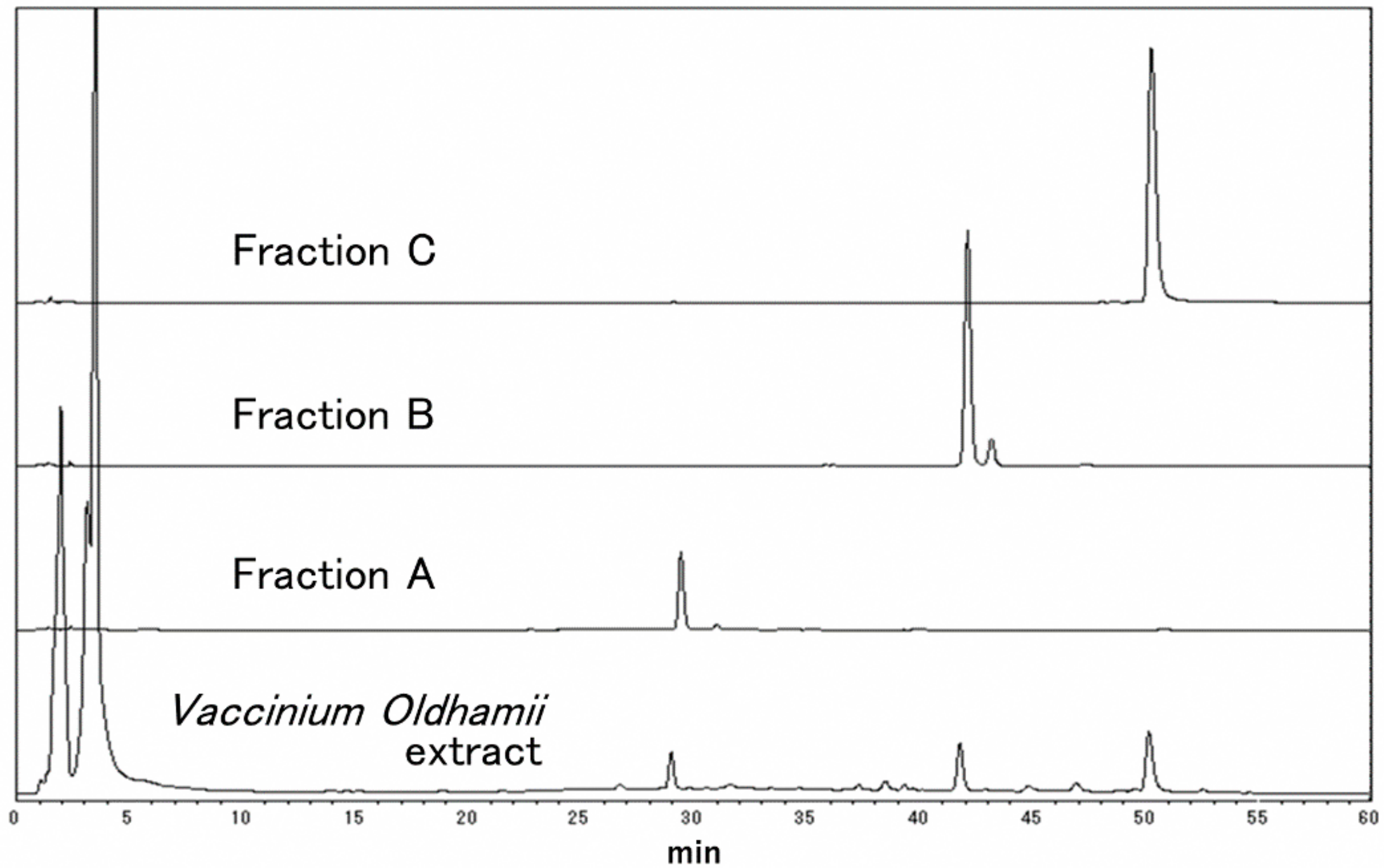
2.3. Separation of Vaccinium oldhamii by HPLC
2.4. Measurement of Total Polyphenol Content in Vaccinium oldhamii
2.5. Measurement of Total Anthocyanin Content in Vaccinium oldhamii
2.6. Cells and Viruses
2.7. IFV Adsorption Inhibition Assay
2.8. Cell Toxicity Assay
2.9. ESI–LC/MS Analysis
3. Results
3.1. Relationships among the IFV Adsorption Inhibitory Activities and Polyphenols in the Vaccinium oldhamii Fraction
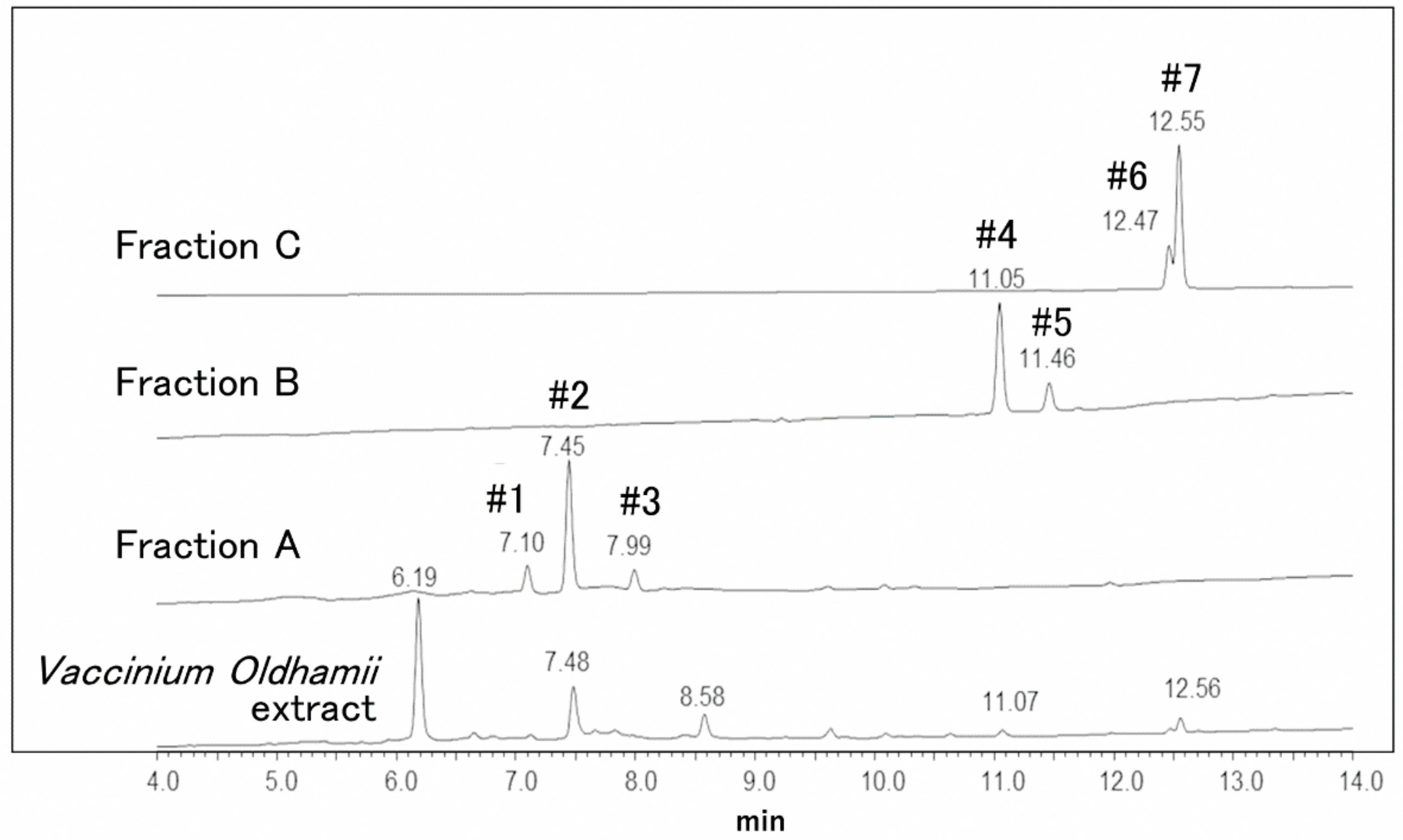
3.2. Mass Spectrometry of Vaccinium oldhamii HPLC Fractions
4. Discussion and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kalt, W.; McDonald, J.E.; Fillmore, S.A.; Tremblay, F. Blueberry Effects on Dark Vision and Recovery after Photobleaching: Placebo-Controlled Crossover Studies. J. Agric. Food Chem. 2014, 46, 11180–11189. [Google Scholar]
- Liu, Y.; Song, X.; Zhang, D.; Zhou, F.; Wang, D.; Wei, Y.; Gao, F.; Xie, L.; Jia, G.; Wu, W.; et al. Blueberry Anthocyanins: Protection against Ageing and Light-Induced Damage in Retinal Pigment Epithelial Cells. J. Nutr. 2012, 108, 16–27. [Google Scholar] [CrossRef]
- Li, J.; Deng, R.; Hua, X.; Zhang, L.; Lu, F.; Coursey, T.G.; Pflugfelder, S.C.; Li, D.-Q. Blueberry Component Pterostilbene Protects Corneal Epithelial Cells from Inflammation via Anti-Oxidative Pathway. Sci. Rep. 2016, 6, 19408. [Google Scholar] [CrossRef]
- Braga, P.C.; Antonacci, R.; Wang, Y.Y.; Lattuada, N.; Dal Sasso, M.; Marabini, L.; Fibiani, M.; Lo Scalzo, R. Comparative Antioxidant Activity of Cultivated and Wild Vaccinium Species Investigated by EPR, Human Neutrophil Burst and COMET Assay. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 1987–1999. [Google Scholar]
- Contreras, R.A.; Köhler, H.; Pizarro, M.; Zúiga, G.E. In Vitro Cultivars of Vaccinium Corymbosum L. (Ericaceae) are a Source of Antioxidant Phenolics. Antioxidants (Basel) 2015, 4, 281–292. [Google Scholar] [CrossRef]
- Bingül, İ.; Başaran-Küçükgergin, C.; Aydın, A.F.; Soluk-Tekkeşin, M.; Olgaç, V.; Doğru-Abbasoğlu, S.; Uysal, M. Blueberry Treatment Attenuated Cirrhotic and Preneoplastic Lesions and Oxidative Stress in the Liver of Diethylnitrosamine-Treated Rats. Int. J. Immunopathol. Pharmacol. 2016, 29, 426–437. [Google Scholar] [CrossRef]
- Suzutani, T.; Ogasawara, M.; Yoshida, I.; Azuma, M.; Knox, Y.M. Anti-Herpesvirus Activity of an Extract of Ribes nigrum L. Phytother. Res. 2003, 17, 609–613. [Google Scholar] [CrossRef]
- Ikuta, K.; Hashimoto, K.; Kaneko, H.; Mori, S.; Ohashi, K.; Suzutani, T. Anti-Viral and Anti-Bacterial Activities of an Extract of the Blackcurrant (Ribes nigrum L.). Microbiol. Immunol. 2012, 56, 805–809. [Google Scholar] [CrossRef]
- Knox, Y.M.; Suzutani, T.; Yoshida, I.; Azuma, M. Anti-Influenza Virus Activity of Crude Extract of Ribes nigrum L. Phytother. Res. 2003, 17, 120–122. [Google Scholar] [CrossRef]
- Knox, Y.M.; Hayashi, K.; Suzutani, T.; Ogasawara, M.; Yosida, I.; Shiina, R.; Tsukui, A.; Terahara, N.; Azuma, M. Activity of Anthocyanins from Fruit Extract of Ribes nigrum L. against Influenza A and B Viruses. Acta Virol. 2001, 45, 209–215. [Google Scholar]
- Sekizawa, H.; Ikuta, K.; Mizuta, K.; Takechi, S.; Suzutani, T. Relationship between Polyphenol Content and Anti-Influenza Viral Effects of Berries. J. Sci. Food Agric. 2013, 93, 2239–2241. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. Methods Enzimol. 1999, 299, 152–178. [Google Scholar]
- Oki, T.; Masuda, M.; Furuta, S.; Nihiba, Y.; Suda, I. Radical Scavenging Activity of Friend Chips Made from Purple-Fleshed Sweet Potato. Food Sci. Technol. Res. 2001, 48, 926–932. [Google Scholar] [CrossRef]
- Ikuta, K.; Mizuta, K.; Suzutani, T. Anti-Influenza Virus Activity of Two Extracts of the Blackcurrant (Ribes nigrum L.) from New Zealand and Poland. Fukushima J. Med. Sci. 2013, 59, 35–38. [Google Scholar] [CrossRef]
- Suzutani, T.; Machida, H.; Sakuma, T. Efficacies of Antiherpesvirus Nucleosides against Two Strains of Herpes Simplex Virus Type 1 in Vero and Human Embryo Lung Fibroblast Cells. Antimicrob. Agents Chemother. 1988, 32, 1046–1052. [Google Scholar] [CrossRef] [PubMed]
- Kammerer, D.; Carle, R.; Schieber, A. Characterization of Phenolic Acids in Black Carrots (Daucus carota ssp. sativus var. Atrorubens Alef.) by High-Performance Liquid Chromatography/Electrospray Ionization Mass Spectrometry. Rapid Commun. Mass Spectrom. 2004, 18, 1331–1340. [Google Scholar] [CrossRef] [PubMed]
- Fang, N.; Yu, S.; Prior, R.L. LC/MS/MS Characterization of Phenolic Constituents in Dried Plums. J. Agric. Food Chem. 2002, 50, 3579–3585. [Google Scholar] [CrossRef]
- Vallverdú-Queralt, A.; Regueiro, J.; Martínez-Huélamo, M.; Rinaldi Alvarenga, J.F.; Leal, L.N.; Lamuela-Raventos, R.M. A Comprehensive Study on the Phenolic Profile of Widely Used Culinary Herbs and Spices: Rosemary, Thyme, Oregano, Cinnamon, Cumin and Bay. Food Chem. 2014, 154, 299–307. [Google Scholar] [CrossRef]
- Jabeur, I.; Martins, N.; Barros, L.; Calhelha, R.C.; Vaz, J.; Achour, L.; Santos-Buelga, C.; Ferreira, I.C. Contribution of the Phenolic Composition to the Antioxidant, Anti-Inflammatory and Antitumor Potential of Equisetum giganteum L. and Tilia Platyphyllos Scop. Food Funct. 2017, 8, 975–984. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, J.I.; Lee, I.; Lee, S.; Hwang, M.W.; Bae, J.Y.; Heo, J.; Kim, D.; Han, S.Z.; Park, M.S. Aronia Melanocarpa and Its Components Demonstrate Antiviral Activity against Influenza Viruses. Biochem. Biophys. Res. Commun. 2013, 440, 14–19. [Google Scholar] [CrossRef]
- Kinoshita, E.; Hayashi, K.; Katayama, H.; Hayashi, T.; Obata, A. Anti-Influenza Virus Effects of Elderberry Juice and Its Fractions. Biosci. Biotechnol. Biochem. 2012, 76, 1633–1638. [Google Scholar] [CrossRef] [Green Version]
- Weiss, E.I.; Houri-Haddad, Y.; Greenbaum, E.; Hochman, N.; Ofek, I.; Zakay-Rones, Z. Cranberry Juice Constituents Affect Influenza Virus Adhesion and Infectivity. Antivir. Res. 2005, 66, 9–12. [Google Scholar] [CrossRef]
- Hariono, M.; Abdullah, N.; Damodaran, K.V.; Kamarulzaman, E.E.; Mohamed, N.; Hassan, S.S.; Shamsuddin, S.; Wahab, H.A. Potential New H1N1 Neuraminidase Inhibitors from Ferulic Acid and Vanillin: Molecular Modelling, Synthesis and in Vitro Assay. Sci. Rep. 2016, 6, 38692. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.F.; Bai, L.P.; Huang, W.B.; Li, X.Z.; Zhao, S.S.; Zhong, N.S.; Jiang, Z.H. Cstructure-Activity Relationship Analysis. Fitoterapia 2015, 93, 47–53. [Google Scholar] [CrossRef]
- Derksen, A.; Hensel, A.; Hafezi, W.; Herrmann, F.; Schmidt, T.J.; Ehrhardt, C.; Ludwig, S.; Kühn, J. 3-O-Galloylated Procyanidins from Rumex acetosa L. Inhibit the Attachment of Influenza A Virus. PLoS ONE 2014, 9, e110089. [Google Scholar] [CrossRef]
- Choi, H.J.; Song, J.H.; Park, K.S.; Kwon, D.H. Inhibitory Effects of Quercetin 3-Rhamnoside on Influenza A Virus Replication. Eur. J. Pharm. Sci. 2009, 37, 329–333. [Google Scholar] [CrossRef]
- Song, J.M.; Kwang, H.L.; Seong, B.L. Antiviral Effect of Catechins in Green Tea on Influenza Virus. Antivir. Res. 2005, 68, 66–74. [Google Scholar] [CrossRef]
- Martynenko, A.; Astatkie, T.; Satanina, V. Novel Hydrothermodynamic Food Processing Technology. J. Food Eng. 2014, 152, 8–16. [Google Scholar] [CrossRef]
- Chen, Y.; Martynenko, A. Storage Stability of Cranberry Puree Products Processed with Hydrothermodynamic (HTD) Technology. LWT Food Sci. Technol. 2017, 79, 543–553. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Fraction No. | Elute Solvent | Volume of Solvent | Weight of Fraction | Total Polyphenol Content | Total Anthocyanin Content | 50% Adsorption Inhibitory Concentration (IC50) | 50% Cytotoxic Concentration (CC50) | Selectivity Index |
|---|---|---|---|---|---|---|---|---|
| (mL) | (mg) | (µg/mL) | (µg/mL) | (µg/mL) | (µg/mL) | (CC50/IC50) | ||
| Fruit (dry) | - | 5000 | - | - | - | - | - | |
| Extract | - | 3077 | 423 | 297 | 72 | 159 | 2.2 | |
| 0 | Water | 200 | 2867 | 28 | 2 | ND | >400 | ND |
| 1 | 10% ethanol | 100 | 76 | 415 | 439 | ND | 236 | ND |
| 2 | 20% ethanol | 100 | 114 | 445 | 377 | 159 | >400 | >2.5 |
| 3 | 30% ethanol | 100 | 47 | 400 | 149 | 38 | 251 | 6.6 |
| 4 | 40% ethanol | 100 | 20 | 327 | 80 | 22 | 160 | 7.3 |
| 5 | 50% ethanol | 100 | 3 | 330 | 90 | 65 | 78 | 1.2 |
| 6 | 80% ethanol | 200 | 4 | 191 | 91 | 85 | 140 | 1.6 |
| Fraction | Fraction Yield | Total Polyphenol Content | IFV Adsorption Inhibitory Activity (IC50) |
|---|---|---|---|
| (mg) | (µg/mL) | (µg/mL) | |
| A | 7.6 | 228 | 38 |
| B | 3.1 | 278 | 40 |
| C | 4.2 | 371 | 238 |
| Fraction | Peak | Retention Time | UV (λmax) | ESI Mode | Precursor Ion (Relative Intensity, %) | Product Ion | Putative Compound |
|---|---|---|---|---|---|---|---|
| No. | (min) | (nm) | (m/z) | (m/z) | |||
| A | #1 | 7.10 | 279 | (−) | 451(32), 425(100), 407(41), 289(16) | 451, 425, 407, 289 | procyanidin B2 |
| (+) | 427(100), 409(63), 291(33) | 427, 429, 291 | |||||
| #2 | 7.45 | 329 | (−) | 295(8), 235(12), 217(55), 193(100), 175(47) | 295, 235, 217, 193, 175 | ferulic acid derivatives | |
| (+) | no information | no information | |||||
| #3 | 7.99 | 325 | (−) | 295(7), 235(12), 217(55), 193(100), 175(45) | 295, 235, 217, 193, 175 | ferulic acid derivatives | |
| (+) | no information | no information | |||||
| B | #4 | 11.05 | 315 | (−) | 193(100) | 193 | ferulic acid O-hexosides |
| (+) | 195(100), 163(45) | 195, 163 | |||||
| #5 | 11.46 | 307 | (−) | 193(100) | 193 | ferulic acid O-hexosides | |
| (+) | 195(100), 163(50) | 195, 163 | |||||
| C | #6 | 12.47 | 349 | (−) | 301(100) | 301 | quercetin O-rhamnoside |
| (+) | 303(100) | 303 | |||||
| #7 | 12.55 | 349 | (−) | 300(100) | 300 | Quercetin O-pentoside O-rhamnoside | |
| (+) | 449(100) | 449 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sekizawa, H.; Ikuta, K.; Ohnishi-Kameyama, M.; Nishiyama, K.; Suzutani, T. Identification of the Components in a Vaccinium oldhamii Extract Showing Inhibitory Activity against Influenza Virus Adsorption. Foods 2019, 8, 172. https://doi.org/10.3390/foods8050172
Sekizawa H, Ikuta K, Ohnishi-Kameyama M, Nishiyama K, Suzutani T. Identification of the Components in a Vaccinium oldhamii Extract Showing Inhibitory Activity against Influenza Virus Adsorption. Foods. 2019; 8(5):172. https://doi.org/10.3390/foods8050172
Chicago/Turabian StyleSekizawa, Haruhito, Kazufumi Ikuta, Mayumi Ohnishi-Kameyama, Kyoko Nishiyama, and Tatsuo Suzutani. 2019. "Identification of the Components in a Vaccinium oldhamii Extract Showing Inhibitory Activity against Influenza Virus Adsorption" Foods 8, no. 5: 172. https://doi.org/10.3390/foods8050172