
Growth Optimization and Rearing of Mealworm (Tenebrio molitor L.) as a Sustainable Food Source

 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Establishment of the Stock Culture of Mealworm
2.2. Diet Preparation
2.3. Parameters Studied
2.4. Nutritional Profile
2.5. Percent Larval Mortality
2.6. Percent Pupal Mortality
3. Statistical Analysis
4. Results
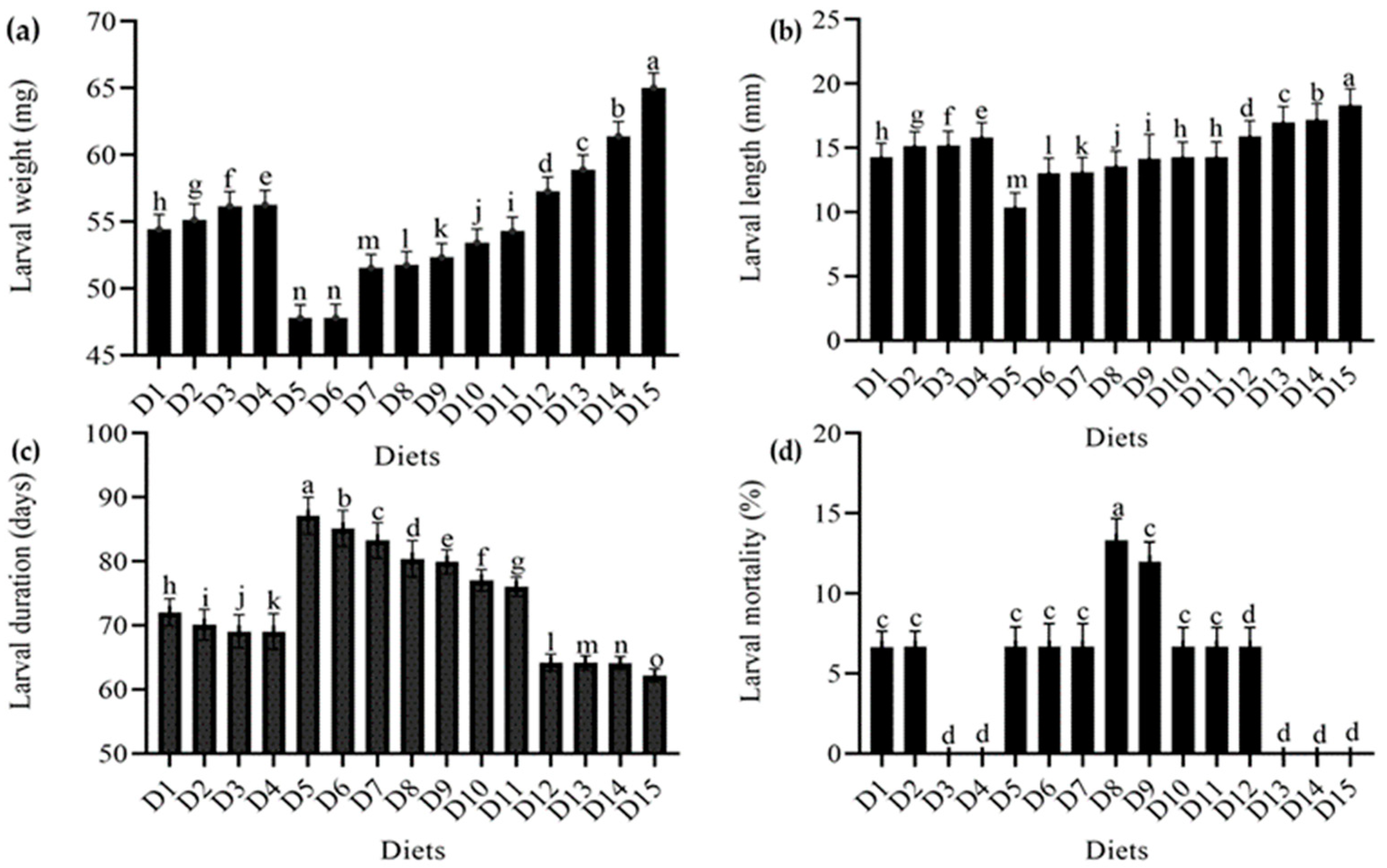
4.1. Effect of Feeding Different Diets on Various Larval Parameters of T. molitor
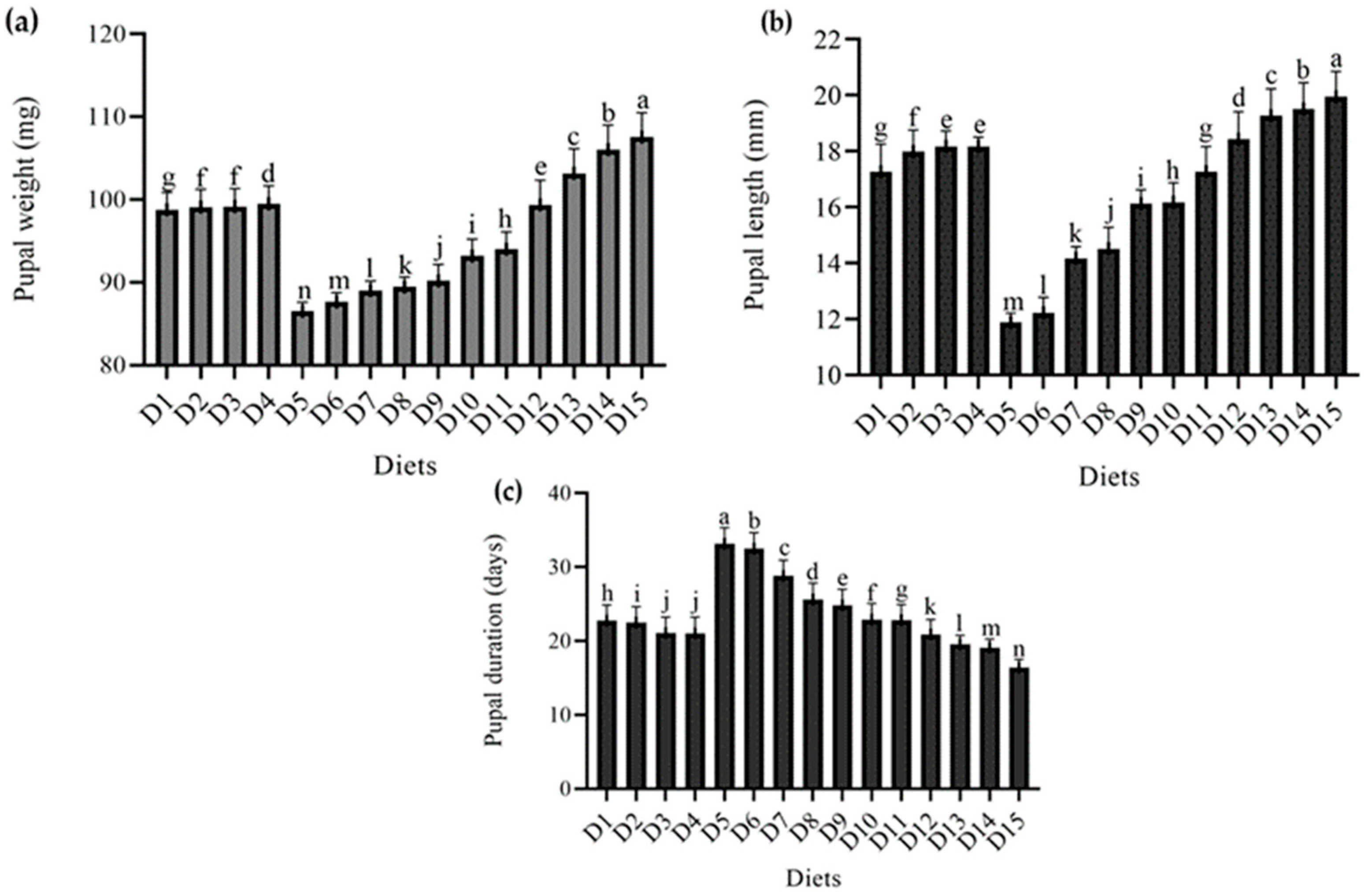
4.2. Effect of Different Diets on Different Pupal Parameters of T. molitor
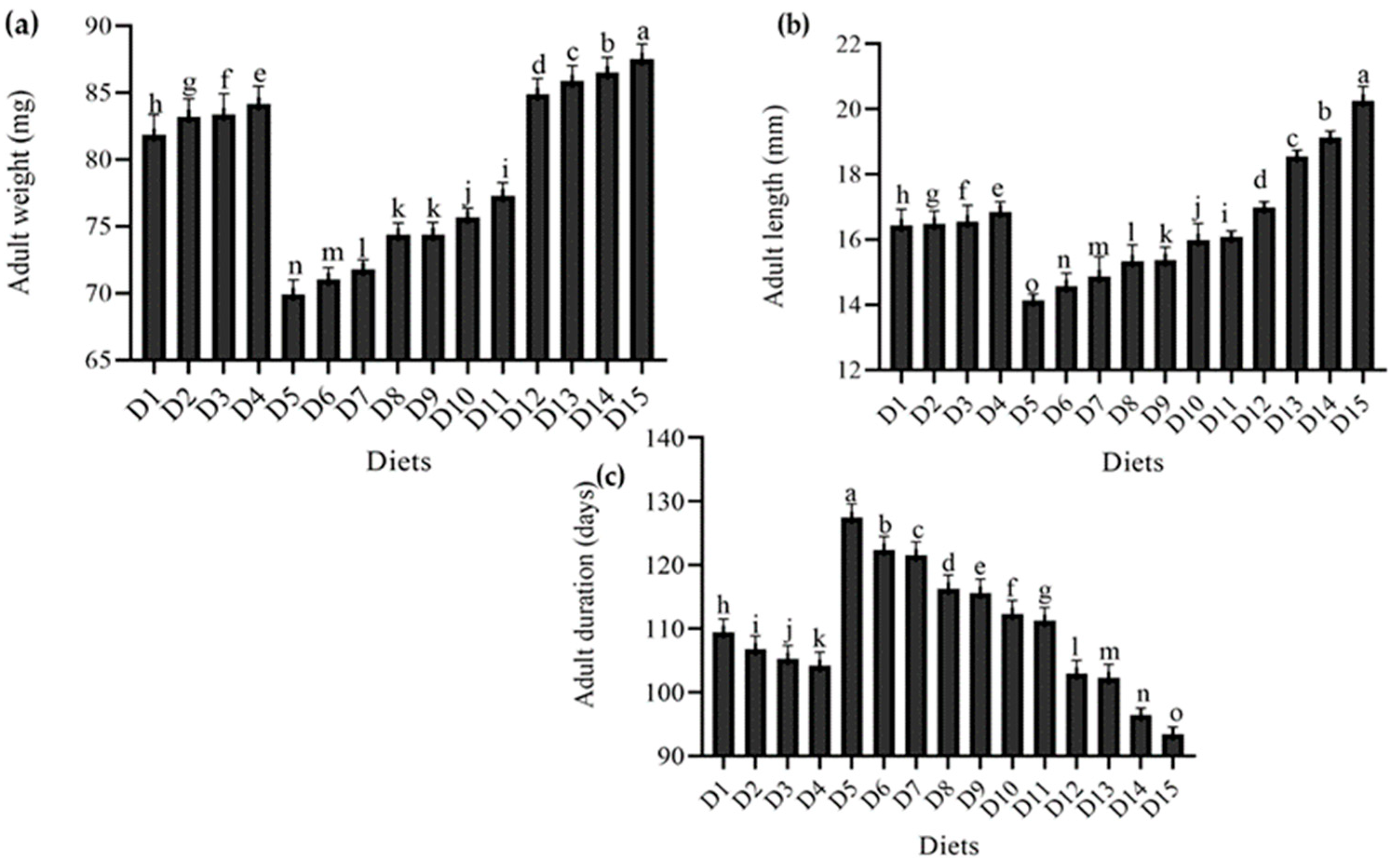
4.3. Effect of Feeding Different Diets on Various Adult Parameters of T. molitor
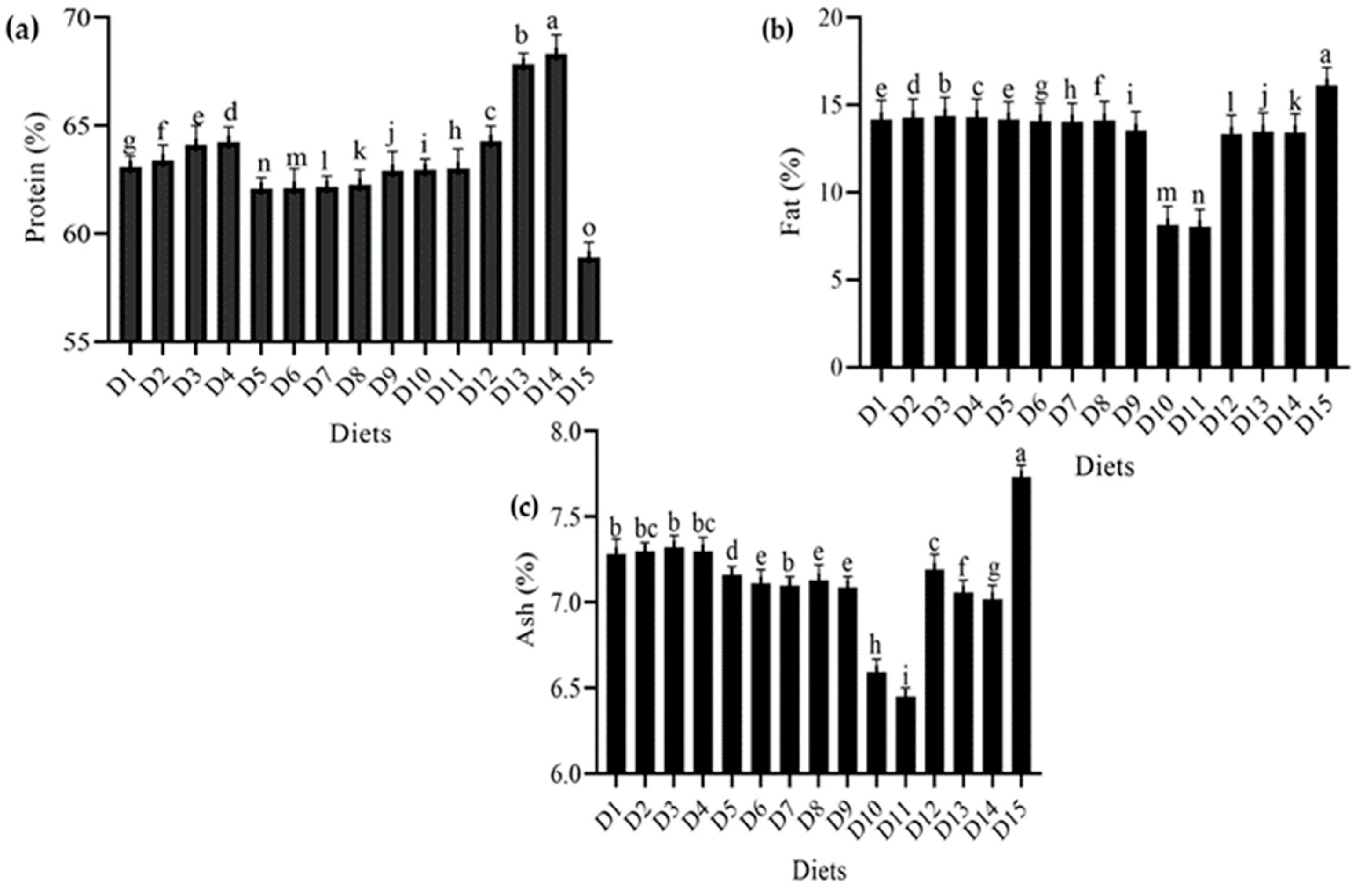
4.4. Effect of Different Diets on Proximate Composition of T. molitor Larvae
5. Discussion
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- DeFoliart, G.R. Insects as human food: Gene DeFoliart discusses some nutritional and economic aspects. Crop Prot. 1992, 11, 395–399. [Google Scholar] [CrossRef]
- Liu, C.; Zhao, J. Insects as a Novel Food. In Encyclopedia of Food Chemistry; Melton, L., Shahidi, F., Varelis, P., Eds.; Academic Press: Oxford, UK, 2019; pp. 428–436. [Google Scholar]
- Hurd, K.J.; Shertukde, S.; Toia, T.; Trujillo, A.; Pérez, R.L.; Larom, D.L.; Love, J.J.; Liu, C. The cultural importance of edible insects in Oaxaca, Mexico. Ann. Entomol. Soc. Am. 2019, 112, 552–559. [Google Scholar] [CrossRef]
- Woolf, E.; Zhu, Y.; Emory, K.; Zhao, J.; Liu, C. Willingness to consume insect-containing foods: A survey in the United States. LWT 2019, 102, 100–105. [Google Scholar] [CrossRef]
- Van Huis, A.; Van Itterbeeck, J.; Klunder, H.; Mertens, E.; Halloran, A.; Muir, G.; Vantomme, P. Edible Insects: Future Prospects for Food and Feed Security; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013. [Google Scholar]
- Ng, W.K.; Liew, F.L.; Ang, L.P.; Wong, K.W. Potential of mealworm (Tenebrio molitor) as an alternative protein source in practical diets for African catfish, Clarias gariepinus. Aquac. Res. 2001, 32, 273–280. [Google Scholar] [CrossRef]
- Rumpold, B.A.; Schlüter, O.K. Nutritional composition and safety aspects of edible insects. Mol. Nutr. Food Res. 2013, 57, 802–823. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Dai, Y.; Zhou, B. Regulatory effects of Tenebrio molitor Linnaeus on immunological function in mice. Afr. J. Biotechnol. 2012, 11, 8348–8352. [Google Scholar]
- Makkar, H.P.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed. Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Melis, R.; Braca, A.; Mulas, G.; Sanna, R.; Spada, S.; Serra, G.; Fadda, M.L.; Roggio, T.; Uzzau, S.; Anedda, R. Effect of freezing and drying processes on the molecular traits of edible yellow mealworm. Innov. Food Sci. Emerg. Technol. 2018, 48, 138–149. [Google Scholar] [CrossRef]
- Ding, M.; Yang, S.; Ding, J.; Zhang, Z.; Zhao, Y.; Dai, W.; Wu, W. Gut Microbiome Associating with Carbon and Nitrogen Metabolism during Biodegradation of Polyethene in Tenebrio larvae with Crop Residues as Co-Diets. Environ. Sci. Technol. 2023, 57, 3031–3041. [Google Scholar] [CrossRef]
- Caparros Megido, R.; Sablon, L.; Geuens, M.; Brostaux, Y.; Alabi, T.; Blecker, C.; Drugmand, D.; Haubruge, É.; Francis, F. Edible insects acceptance by B elgian consumers: Promising attitude for entomophagy development. J. Sens. Stud. 2014, 29, 14–20. [Google Scholar] [CrossRef]
- Ghaly, A.E.; Alkoaik, F. The yellow mealworm as a novel source of protein. Am. J. Agric. Biol. Sci. 2009, 4, 319–331. [Google Scholar] [CrossRef]
- Siemianowska, E.; Kosewska, A.; Aljewicz, M.; Skibniewska, K.A.; Juszczak, L.P.; Jarocki, A.; Jedras, M. Larvae of mealworm (Tenebrio molitor L.) as European novel food. Agri Sci. 2013, 4, 287–291. [Google Scholar]
- Choi, H.-S.; Kim, S.-A.; Shin, H.-J. Present and perspective on insect biotechnology. KSBB J. 2015, 30, 257–267. [Google Scholar] [CrossRef]
- Hu, B.; Das, P.; Lv, X.; Shi, M.; Aa, J.; Wang, K.; Wu, X. Effects of ‘Healthy’ Fecal Microbiota Transplantation against the Deterioration of Depression in Fawn-Hooded Rats. mSystems 2022, 7, e21822. [Google Scholar] [CrossRef]
- Weaver, D.K.; McFarlane, J. The effect of larval density on growth and development of Tenebrio molitor. J. Insect Physiol. 1990, 36, 531–536. [Google Scholar] [CrossRef]
- Damborsky, M.; Sandrigo-Ybran, T.; Oscherov, E. Ciclo de Vida de Tenebrio molitor (Coleoptera, Tenebrionidae) en Condiciones Experimentales. 2000. Available online: http://www.unne.edu.ar/unnevieja/Web/cyt/cyt/biologia/b-011.pdf (accessed on 14 February 2023).
- Hill, D.S. Pests of Stored Foodstuffs and Their Control; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2002. [Google Scholar]
- Fanatico, A.C.; Arsi, K.; Upadhyaya, I.; Morales-Ramos, J.; Donoghue, D.; Donoghue, A.M. Sustainable fish and invertebrate meals for methionine and protein feeds in organic poultry production. J. Appl. Poult. Res. 2018, 27, 437–448. [Google Scholar] [CrossRef]
- Kröncke, N.; Grebenteuch, S.; Keil, C.; Demtröder, S.; Kroh, L.; Thünemann, A.F.; Benning, R.; Haase, H. Effect of different drying methods on nutrient quality of the yellow mealworm (Tenebrio molitor L.). Insects 2019, 10, 84. [Google Scholar] [CrossRef]
- Finke, M.D. Complete nutrient composition of commercially raised invertebrates used as food for insectivores. Zoo. Biol. 2002, 21, 269–285. [Google Scholar] [CrossRef]
- Sun, J.; Jia, Q.; Li, Y.; Zhang, T.; Chen, J.; Ren, Y.; Fu, S. Effects of Arbuscular Mycorrhizal Fungi and Biochar on Growth, Nutrient Absorption, and Physiological Properties of Maize (Zea mays L.). J. Fungi 2022, 8, 1275. [Google Scholar] [CrossRef]
- Yang, K.; Geng, Q.; Luo, Y.; Xie, R.; Sun, T.; Wang, Z.; Tian, J. Dysfunction of FadA-cAMP signalling decreases Aspergillus flavus resistance to antimicrobial natural preservative Perillaldehyde and AFB1 biosynthesis. Environ. Microbiol. 2022, 24, 1590–1607. [Google Scholar] [CrossRef]
- Pan, C.; Yang, K.; Erhunmwunsee, F.; Li, Y.; Liu, M.; Pan, S.; Tian, J. Inhibitory effect of cinnamaldehyde on Fusarium solani and its application in postharvest preservation of sweet potato. Food Chem. 2023, 408, 135213. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, S.; Yang, X.; Wang, W.; Liu, X.; Wang, H.; Zhang, H. Enhancing the fermentation performance of frozen dough by ultrasonication: Effect of starch hierarchical structures. J. Cereal Sci. 2022, 106, 103500. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, S.; Yang, X.; Zhang, J.; Zhang, Y.; Liu, X.; Wang, H. Effect of germination on nutritional properties and quality attributes of glutinous rice flour and dumplings. J. Food. Compos. Anal. 2022, 108, 104440. [Google Scholar] [CrossRef]
- Li, L.; Xie, B.; Dong, C.; Hu, D.; Wang, M.; Liu, G.; Liu, H. Rearing Tenebrio molitor L. (Coleptera: Tenebrionidae) in the “Lunar Palace 1” during a 105-day multi-crew closed integrative BLSS experiment. Life Sci. Space Res. 2015, 7, 9–14. [Google Scholar] [CrossRef]
- Veldkamp, T.; Van Duinkerken, G.; van Huis, A.; Lakemond CM, M.; Ottevanger, E.; Bosch, G.; Van Boekel, T. Insects as a Sustainable Feed Ingredient in Pig and Poultry Diets—A Feasibility Study; Rapport 638; Wageningen Livestock Research: Wageningen, The Netherlands, 2012. [Google Scholar]
- Bovera, F.; Loponte, R.; Marono, S.; Piccolo, G.; Parisi, G.; Iaconisi, V.; Gasco, L.; Nizza, A. Use of larvae meal as protein source in broiler diet: Effect on growth performance, nutrient digestibility, and carcass and meat traits. J. Anim. Sci. 2016, 94, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.D.; Rivers, J.P.W.; Cowgill, U.M. Culturing mealworms as food for animals in captivity. Int. Zoo Yearb. 1976, 16, 63–70. [Google Scholar] [CrossRef]
- Durst, P.D.; Shono, K. Edible forest insects: Exploring new horizons and traditional practices. In Forest Insects as Food: Humans Bite Back; Durst, P.B., Johnson, D.V., Leslie, R.N., Shono, K., Eds.; Food and Agricultural Organization of the United Nations, Regional Office for Asia and Pacific: Bangkok, Thailand, 2010; pp. 1–4. [Google Scholar]
- Vantomme, P.; Mertens, E.; van Huis, A.; Klunder, H. Assessing the Potential of Insects as Food and Feed in Assuring Food Security: Summary Report; Food and Agricultural Organization of the United Nations, Forestry Department: Rome, Italy, 2012. [Google Scholar]
- Codex Alimentarius Commission (Ed.) Food Labelling, 5th ed.; World Health Organization: Rome, Italy, 2007. [Google Scholar]
- Nowak, V.; Persijn, D.; Rittenschober, D.; Charrondiere, U. Review of food composition data for edible insects. Food Chem. 2016, 193, 39–46. [Google Scholar] [CrossRef]
- Zhang, F.; Pant, D.; Logan, B.E. Long-term performance of activated carbon air cathodes with different diffusion layer porosities in microbial fuel cells. Biosens. Bioelectron. 2011, 30, 49–55. [Google Scholar] [CrossRef]
- Feng, Q.; Feng, Z.; Su, X.; Bai, Y.D.; Ding, B.Y. Design and Simulation of Human Resource Allocation Model Based on Double-Cycle Neural Network. Comput. Intel. Neurosci. 2021, 2021, 7149631. [Google Scholar] [CrossRef]
- Gao, J.; Sun, H.; Han, J.; Sun, Q.; Zhong, T. Research on Recognition Method of Electrical Components Based on FEYOLOv4-tiny. J. Elect. Engineer. Technol. 2022, 17, 3541–3551. [Google Scholar] [CrossRef]
- Bordiean, A.; Krzyżaniak, M.; Stolarski, M.J.; Czachorowski, S.; Peni, D. Will yellow mealworm become a source of safe proteins for Europe? Agriculture 2020, 10, 233. [Google Scholar] [CrossRef]
- Mellanby, K. The effect of atmospheric humidity on the metabolism of the fasting mealworm (Tenebrio molitor L., Coleoptera). Proc. R. Soc. Lond. 1932, 111, 376–390. [Google Scholar]
- Lyons, W. Rearing Mealworms Fact Sheet; Entomology Center, Ohio State University: Columbia, OH, USA, 1991. [Google Scholar]
- Gibson, R. The Multipurpose Mealworm Leaping from the Box.com. 2009. Available online: http://www.leapingfromthebox.com/art/rlg/mealworms.html (accessed on 14 February 2023).
- Morales-Ramos, J.A.; Rojas, M.G.; Shapiro-llan, D.I.; Tedders, W.L. Use of nutrient self-selection as a diet refining tool in Tenebrio molitor (Coleoptera: Tenebrionidae). J. Entomol. Sci. 2013, 48, 206–221. [Google Scholar] [CrossRef]
- Hardouin, J.; Mahoux, G. Zootechnie D’insects—Elevage et Utilization au Benefice de I’homme et de Certains Animaux; Bureau Pour I’ Echange et la Distribution de I’ Information sur le Mini-Elevage (BEDIM): Gembloux, Belgium, 2003; 164p. [Google Scholar]
- Greenberg, S.; Ar, A. Effects of chronic hypoxia, normoxia, and hyperoxia on larval development in the beetle Tenebrio molitor. J. Insect Physiol. 1996, 42, 991–996. [Google Scholar] [CrossRef]
- Fraenkel, G. The nutrition of the mealworm, Tenebrio molitor L. (Tenebrionidae, Coleoptera). Physiol. Zool. 1950, 23, 92–108. [Google Scholar] [CrossRef]
- Cheng, M.; Cui, Y.; Yan, X.; Zhang, R.; Wang, J.; Wang, X. Effect of dual-modified cassava starches on intelligent packaging films containing red cabbage extracts. Food Hydrocoll. 2022, 124, 107225. [Google Scholar] [CrossRef]
- Yuan, Q.; Kato, B.; Fan, K.; Wang, Y. Phased array guided wave propagation in curved plates. Mech. Syst. Signal Process. 2023, 185, 109821. [Google Scholar] [CrossRef]
- Kim, S.Y.; Park, J.B.; Lee, Y.B.; Yoon, H.J.; Lee, K.Y.; Kim, N.J. Growth characteristics of mealworm (Tenebrio molitor). J. Sericultural Entomol. Sci. 2015, 53, 1–5. [Google Scholar] [CrossRef]
- Cotton, R.T. Notes on the biology of the meal worms, Tenebrio molitor Linne and T. obscurus Fab. Ann. Entomol. Soc. Am. 1927, 20, 81–86. [Google Scholar] [CrossRef]
- Aguilar-Miranda, E.D.; Lopez, M.G.; Escamilla-Santana, C.; Rosa, A.P. Characteristics of maize flour tortilla supplemented with ground Tenebrio molitor larvae. J. Agr. Food Chem. 2002, 50, 192–195. [Google Scholar] [CrossRef]
- Barker, D.; Fitzpatrick, M.P.; Dierenfeld, E.S. Nutrient compotion of selected whole invertebrates. Zoo Biol. 1998, 17, 123–134. [Google Scholar] [CrossRef]
- Ravzanaadii, N.; Kim, S.H.; Choi, W.H.; Hong, S.J.; Kim, N.J. Nutritional value of mealworm, Tenebrio molitor as food source. Int. J. Ind. Entomol. 2012, 25, 93–98. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riaz, K.; Iqbal, T.; Khan, S.; Usman, A.; Al-Ghamdi, M.S.; Shami, A.; El Hadi Mohamed, R.A.; Almadiy, A.A.; Al Galil, F.M.A.; Alfuhaid, N.A.; et al. Growth Optimization and Rearing of Mealworm (Tenebrio molitor L.) as a Sustainable Food Source. Foods 2023, 12, 1891. https://doi.org/10.3390/foods12091891
Riaz K, Iqbal T, Khan S, Usman A, Al-Ghamdi MS, Shami A, El Hadi Mohamed RA, Almadiy AA, Al Galil FMA, Alfuhaid NA, et al. Growth Optimization and Rearing of Mealworm (Tenebrio molitor L.) as a Sustainable Food Source. Foods. 2023; 12(9):1891. https://doi.org/10.3390/foods12091891
Chicago/Turabian StyleRiaz, Kanwal, Toheed Iqbal, Sarzamin Khan, Amjad Usman, Mariam S. Al-Ghamdi, Ashwag Shami, Rania Ali El Hadi Mohamed, Abdulrahman A. Almadiy, Fahd Mohammed Abd Al Galil, Nawal Abdulaziz Alfuhaid, and et al. 2023. "Growth Optimization and Rearing of Mealworm (Tenebrio molitor L.) as a Sustainable Food Source" Foods 12, no. 9: 1891. https://doi.org/10.3390/foods12091891