Novel Property Cytotoxicity and Mechanism of Food Preservative Brevilaterins against Human Gastric Cancer Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. AMPs, Cell Lines and Materials
2.2. Effect of Brevilaterins on the Proliferation of Cells
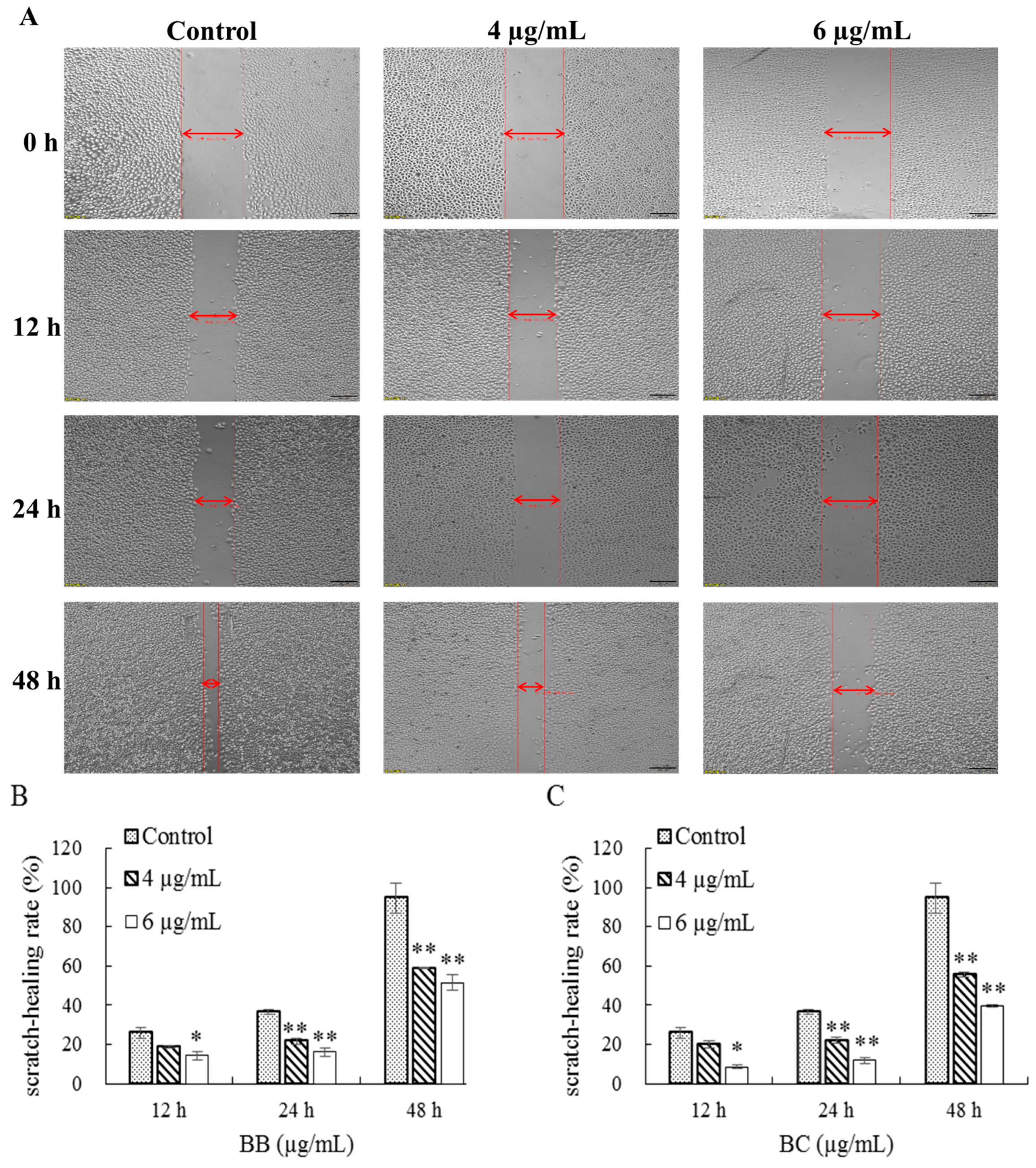
2.3. Effect of Brevilaterins on the Migration of Cells
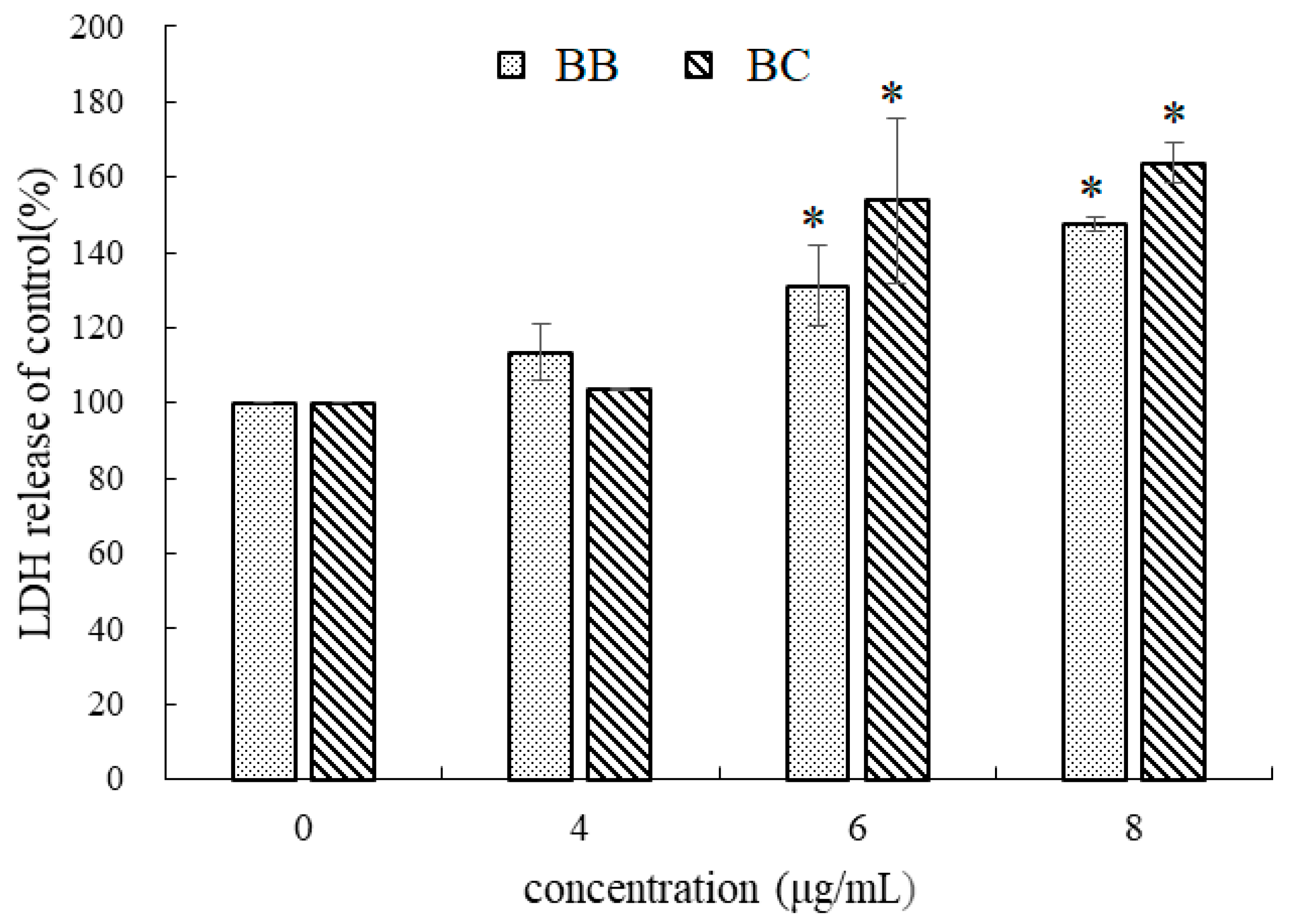
2.4. Effect of Brevilaterins on the Membrane Permeability
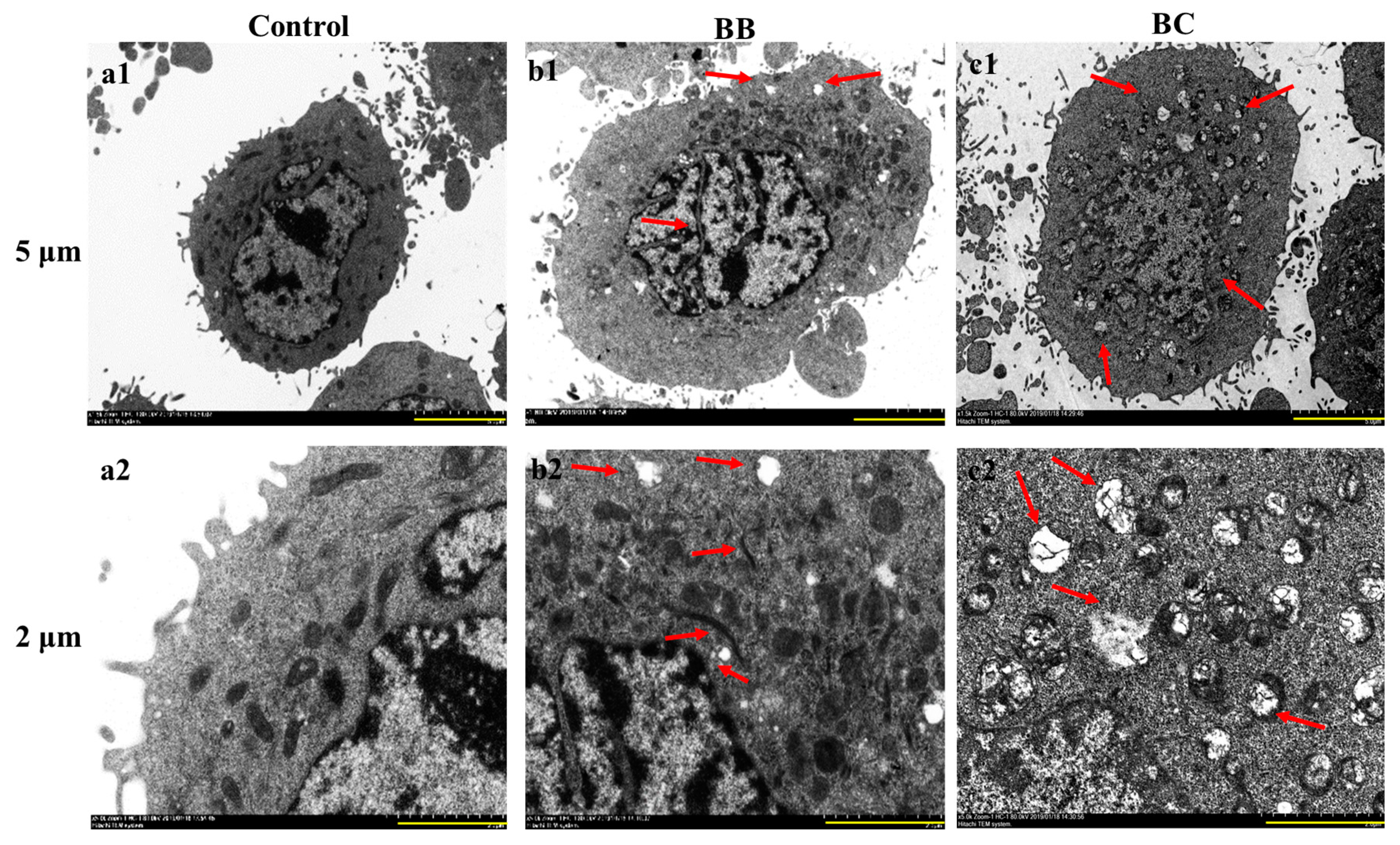
2.5. Impact of Brevilaterins on the Ultrastructural Features of Cancer Cells
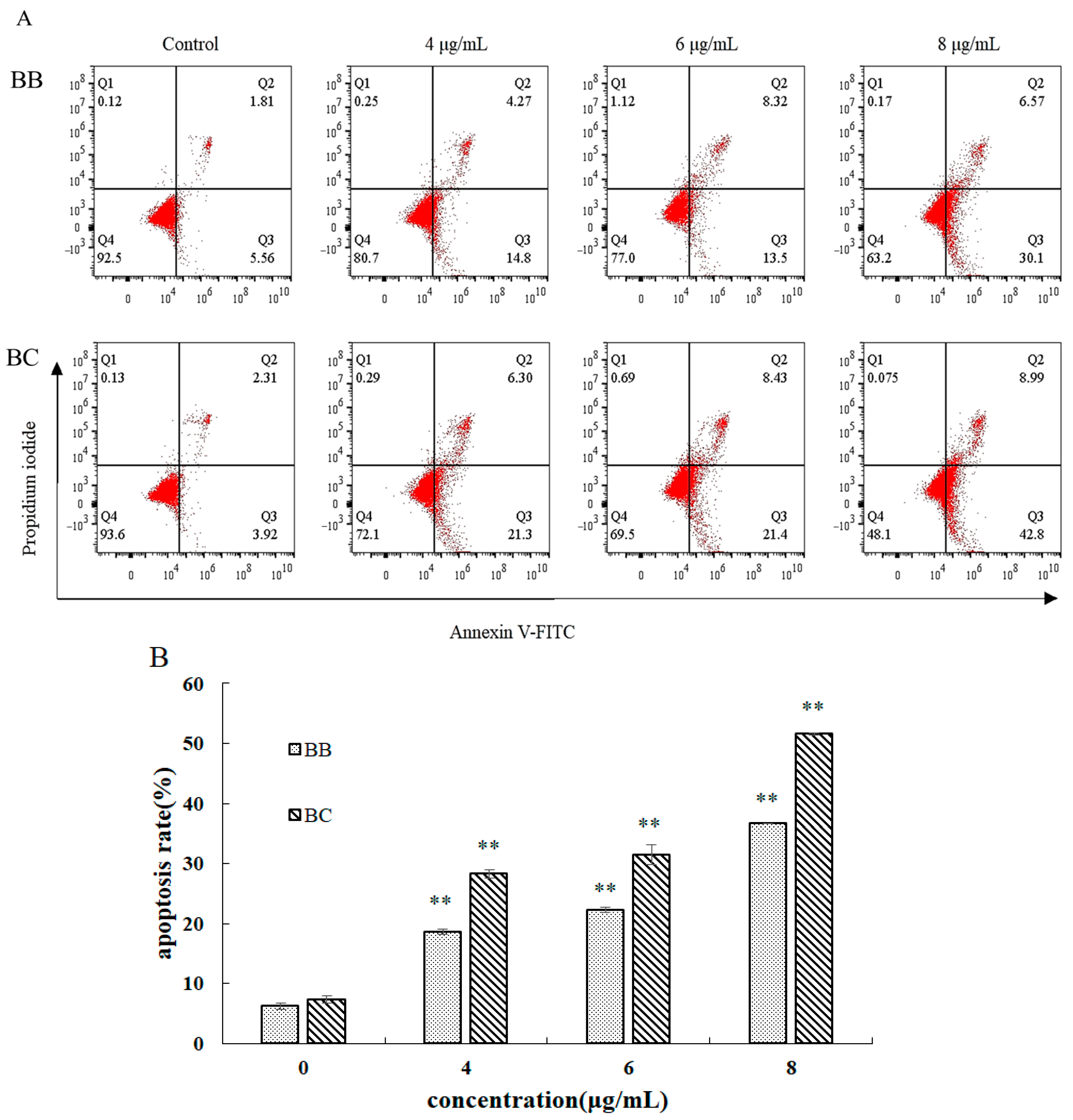
2.6. Effect of Brevilaterins on the Apoptosis of Cancer Cells
2.7. Statistical Analysis
3. Results and Discussion
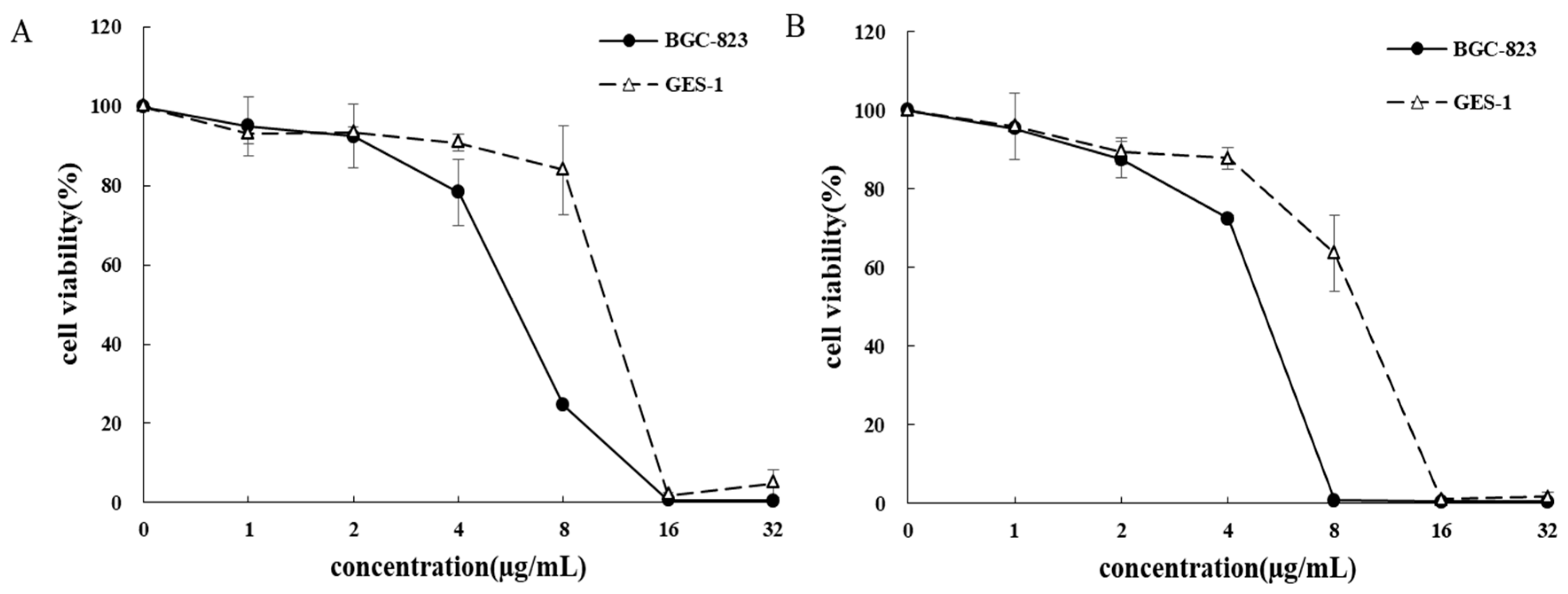
3.1. Inhibition of BB/BC on the Proliferation of Gastric Cancer Cells
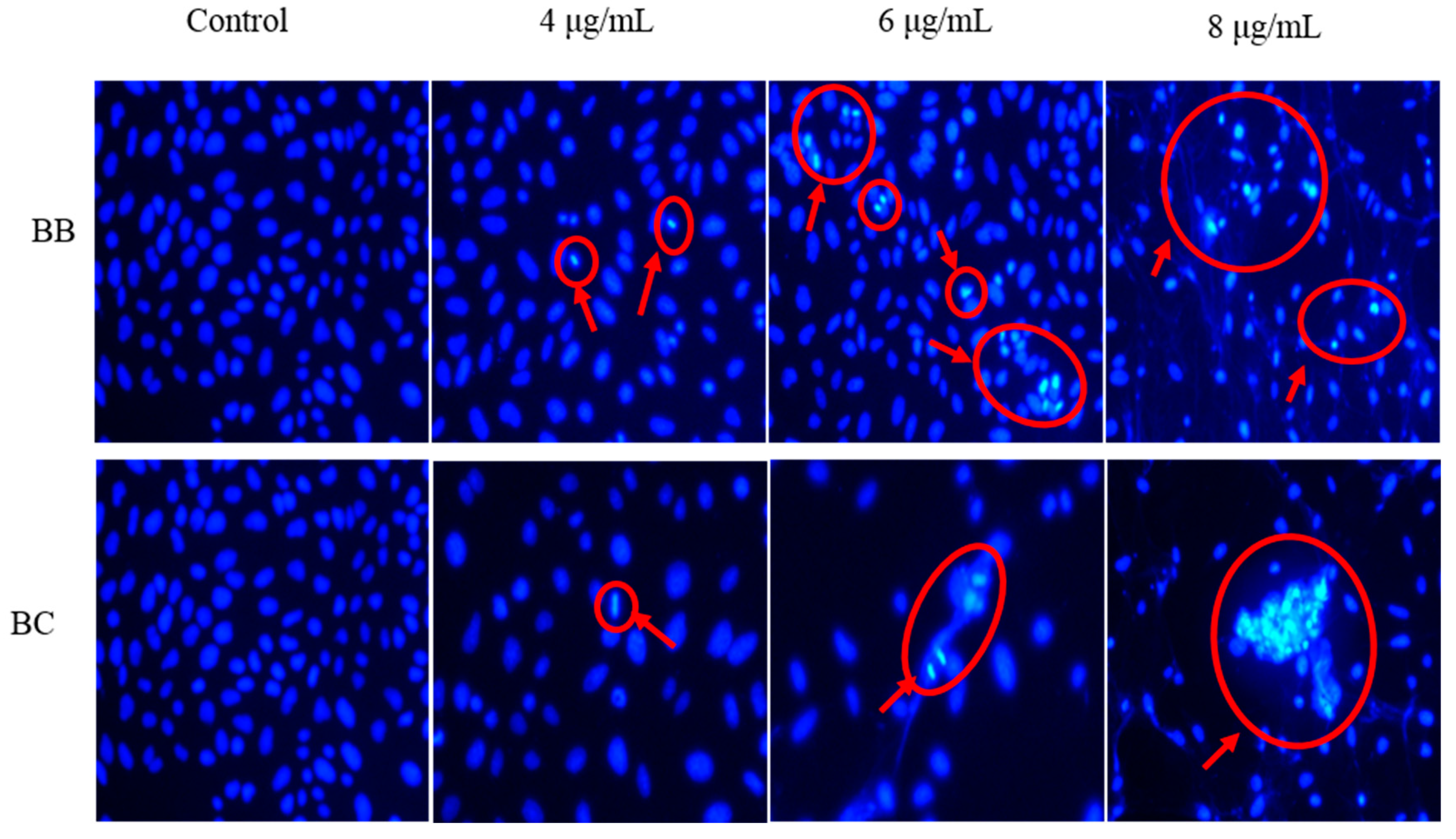
3.2. Effect on the Changes in Gastric Cancer Cell Morphology of BB/BC
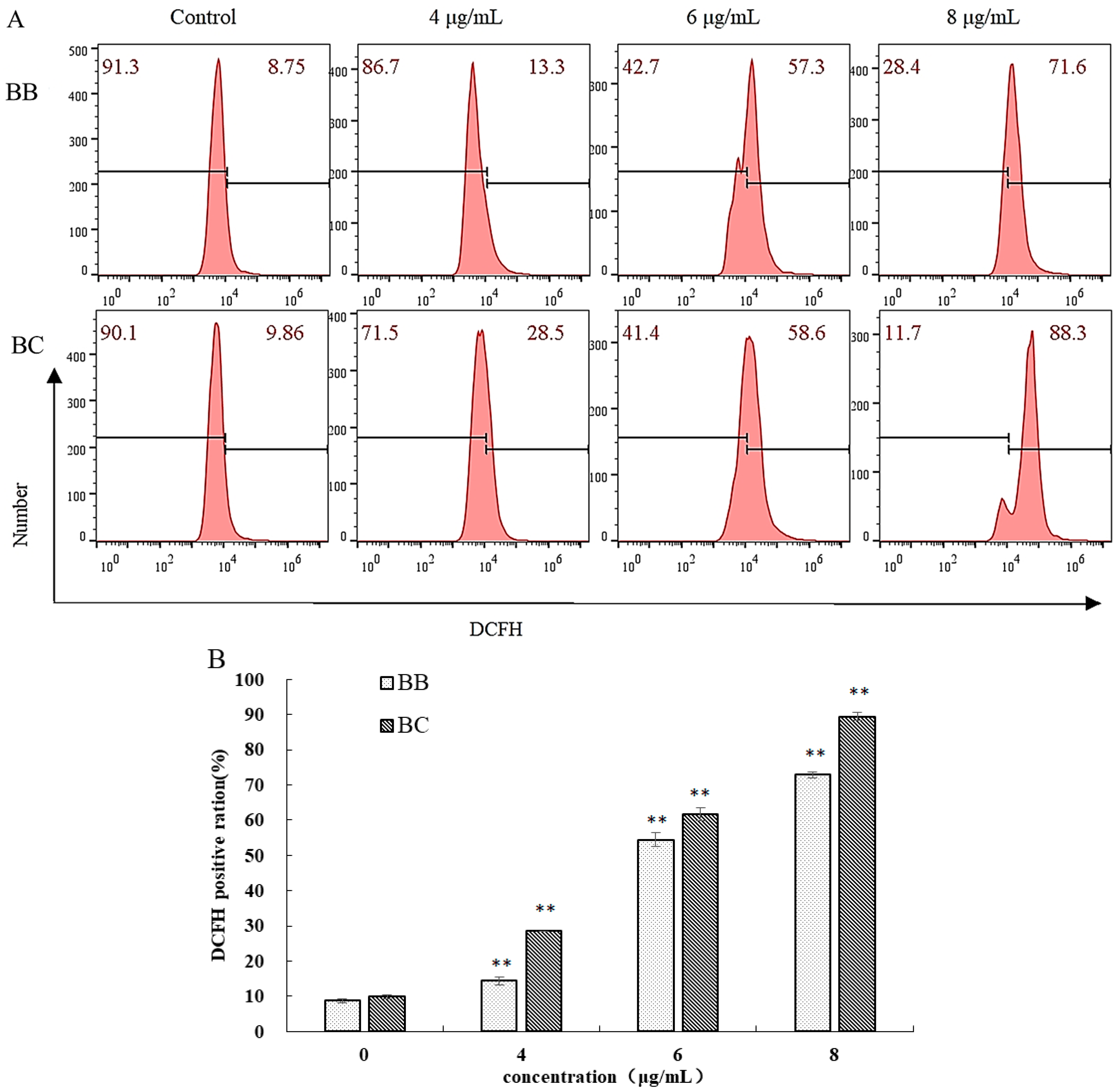
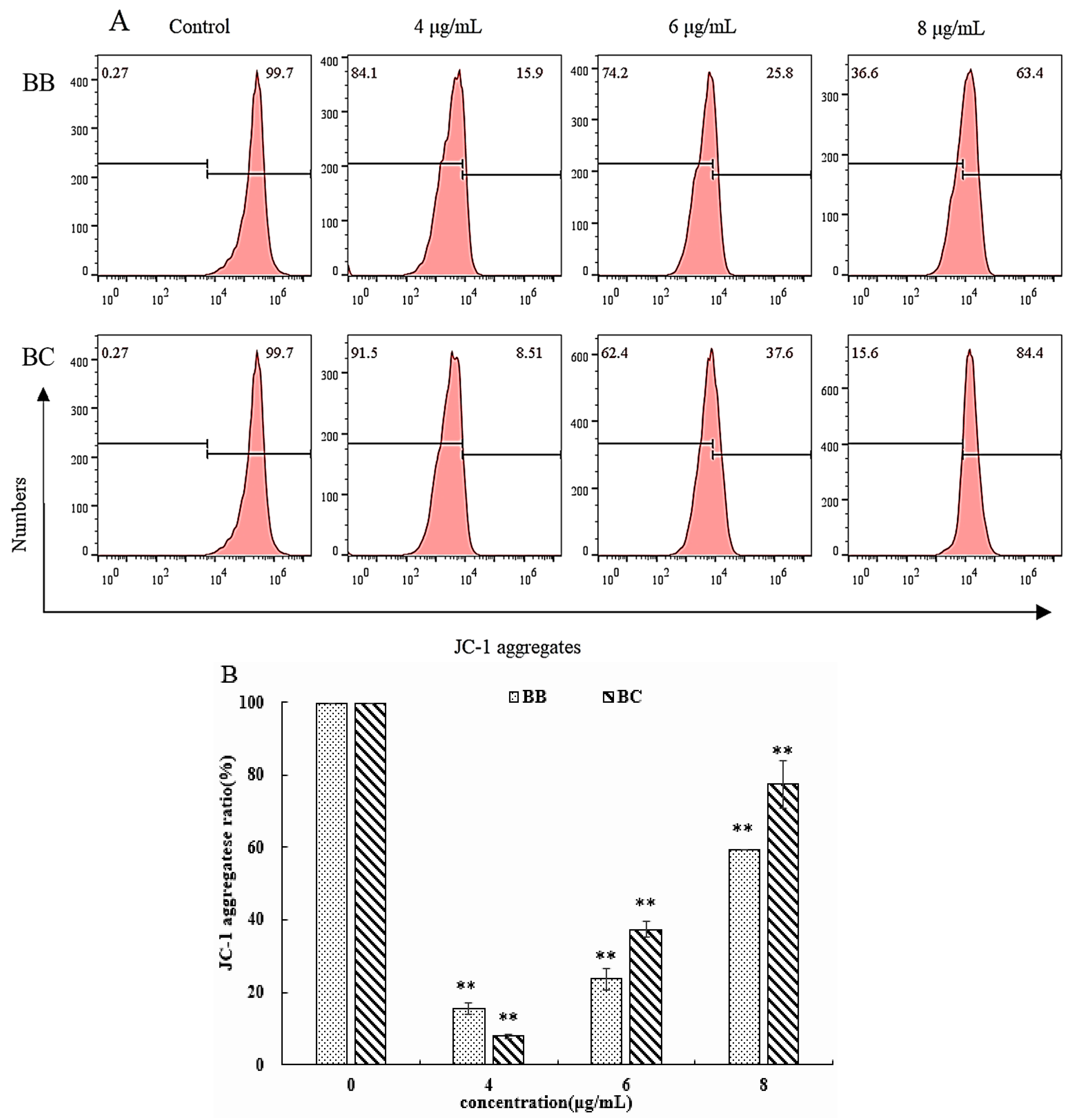
3.3. Further Study on the Apoptotic Mechanism of BB/BC towards Gastric Cancer Cells
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-D.; Zhang, P.-F.; Xi, H.-Q.; Wei, B.; Chen, L.; Tang, Y. Recent advances in the diagnosis, staging, treatment, and prognosis of advanced gastric cancer: A literature review. Front. Med. 2021, 8, 744839. [Google Scholar] [CrossRef] [PubMed]
- Proserpio, I.; Rausei, S.; Barzaghi, S.; Frattini, F.; Galli, F.; Iovino, D.; Rovera, F.; Boni, L.; Dionigi, G.; Pinotti, G. Multimodal treatment of gastric cancer. World J. Gastrointest. Surg. 2014, 6, 55–58. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [PubMed]
- Oyston, P.C.; Fox, M.A.; Richards, S.J.; Clark, G.C. Novel peptide therapeutics for treatment of infections. J. Med. Microbiol. 2009, 58 Pt 8, 977–987. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, X.; Han, P.; Liu, Y.; Hong, D.; Li, S.; Ma, A.; Jia, Y. Discovery of novel antimicrobial peptides, Brevilaterin V, from Brevibacillus laterosporus S62-9 after regulated by exogenously-added L-valine. LWT 2022, 155, 112962. [Google Scholar] [CrossRef]
- Giuliani, A.; Pirri, G.; Nicoletto, S. Antimicrobial peptides: An overview of a promising class of therapeutics. Cent. Eur. J. Biol. 2007, 2, 1–33. [Google Scholar] [CrossRef]
- Ting, C.-H.; Huang, H.-N.; Huang, T.-C.; Wu, C.-J.; Chen, J.-Y. The mechanisms by which pardaxin, a natural cationic antimicrobial peptide, targets the endoplasmic reticulum and induces c-FOS. Biomaterials 2014, 35, 3627–3640. [Google Scholar] [CrossRef]
- Kim, S.-Y.; Kim, J.Y.; Kim, S.-H.; Bae, H.J.; Yi, H.; Yoon, S.H.; Koo, B.S.; Kwon, M.; Cho, J.Y.; Lee, C.-E.; et al. Surfactin from Bacillus subtilis displays anti-proliferative effect via apoptosis induction, cell cycle arrest and survival signaling suppression. FEBS Lett. 2007, 581, 865–871. [Google Scholar] [CrossRef]
- Park, S.Y.; Kim, J.-H.; Lee, Y.J.; Lee, S.J.; Kim, Y. Surfactin suppresses TPA-induced breast cancer cell invasion through the inhibition of MMP-9 expression. Int. J. Oncol. 2012, 42, 287–296. [Google Scholar] [CrossRef]
- Jiang, H.; Ji, C.; Sui, J.; Sa, R.; Wang, X.; Liu, X.; Guo, T.L. Antibacterial and antitumor activity of Bogorol B-JX isolated from Brevibacillus laterosporus JX-5. World J. Microbiol. Biotechnol. 2017, 33, 177. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, K.; Shibasaki, C.; Takahashi, K.; Nakamura, T.; Takeuchi, T.; Umezawa, H. Antitumor activity of spergualin, a novel antitumor antibiotic. J. Antibiot. 1986, 39, 1461–1466. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Han, P.; Jia, Y.; Chen, Z.; Li, S.; Ma, A. Antibacterial regularity mining beneath the systematic activity database of lipopeptides Brevilaterins: An instructive activity handbook for its food application. Foods 2022, 11, 2991. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, L.; Hong, D.; Liu, Y.; Han, P.; Li, S.; Jia, Y. Broad-spectrum cytotoxicity to cancer cells of Brevilaterin C from Brevibacillus laterosporus and its specific mechanism on human epidermal cancer cells. J. Cell. Biochem. 2022, 123, 1237–1246. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, L.; Liu, Y.; Han, P.; Hong, D.; Li, S.; Ma, A.; Jia, Y. Brevilaterin B from Brevibacillus laterosporus has selective antitumor activity and induces apoptosis in epidermal cancer. World J. Microbiol. Biotechnol. 2022, 38, 201. [Google Scholar] [CrossRef] [PubMed]
- Deslouches, B.; Steckbeck, J.D.; Craigo, J.K.; Doi, Y.; Mietzner, T.A.; Montelaro, R.C. Rational design of engineered cationic antimicrobial peptides consisting exclusively of arginine and tryptophan, and Their Activity against multidrug-resistant pathogens. Antimicrob. Agents Chemother. 2013, 57, 2511–2521. [Google Scholar] [CrossRef] [PubMed]
- Deslouches, B.; Steckbeck, J.D.; Craigo, J.K.; Doi, Y.; Burns, J.L.; Montelaro, R.C. Engineered cationic antimicrobial peptides to overcome multidrug resistance by ESKAPE pathogens. Antimicrob. Agents Chemother. 2015, 59, 1329–1333. [Google Scholar] [CrossRef]
- Steckbeck, J.D.; Deslouches, B.; Montelaro, R.C. Antimicrobial peptides: New drugs for bad bugs? Expert. Opin. Biol. Ther. 2014, 14, 11–14. [Google Scholar] [CrossRef] [PubMed]
- Flores-Guzmán, F.; Alvarado-Sansininea, J.J.; López-Muñoz, H.; Escobar, M.L.; Espinosa-Trejo, M.; Tavera-Hernandez, R.; Jiménez-Estrada, M.; Sánchez-Sánchez, L. Antiproliferative, cytotoxic and apoptotic activity of the bentonite transformation of sesquiterpene lactone glaucolide B to 5β-hydroxy-hirsutinolide on tumor cell lines. Eur. J. Pharmacol. 2019, 856, 172406. [Google Scholar] [CrossRef]
- He, N.; Shi, X.; Zhao, Y.; Tian, L.; Wang, N.; Yang, X. Inhibitory effects and molecular mechanisms of selenium-containing tea polysaccharides on human breast cancer MCF-7 Cells. J. Agric. Food Chem. 2013, 61, 579–588. [Google Scholar] [CrossRef]
- Cerón, J.M.; Contreras-Moreno, J.; Puertollano, E.; de Cienfuegos, G.; Puertollano, M.A.; de Pablo, M.A. The antimicrobial peptide cecropin A induces caspase-independent cell death in human promyelocytic leukemia cells. Peptides 2010, 31, 1494–1503. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Fan, X.-X.; Jiang, Z.-B.; Loo, W.T.; Yao, X.-J.; Leung, E.L.-H.; Chow, L.W.; Liu, L. Shikonin inhibits gefitinib-resistant non-small cell lung cancer by inhibiting TrxR and activating the EGFR proteasomal degradation pathway. Pharmacol. Res. 2017, 115, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, L.; Ma, C.; Zhang, Y.; Xi, X.; Wang, L.; Zhou, M.; Burrows, J.F.; Chen, T. A novel antimicrobial peptide, Ranatuerin-2PLx, showing therapeutic potential in inhibiting proliferation of cancer cells. Biosci. Rep. 2018, 38, BSR20180710. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhou, Y.; Li, S.; Li, H.; Tian, L.; Shang, D. Anticancer mechanisms of temporin-1CEa, an amphipathic α-helical antimicrobial peptide, in Bcap-37 human breast cancer cells. Life Sci. 2013, 92, 1004–1014. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, D.; Veiga, A.S.; Castanho, M.A.R.B. From antimicrobial to anticancer peptides. A review. Front. Microbiol. 2013, 4, 294. [Google Scholar] [CrossRef]
- Guo, T.L.; Miller, M.A.; Shapiro, I.M.; Shenker, B.J. Mercuric chloride induces apoptosis in human T lymphocytes: Evidence of mitochondrial dysfunction. Toxicol. Appl. Pharmacol. 1998, 153, 250–257. [Google Scholar] [CrossRef]
- Stoian, I.; Oros, A.; Moldoveanu, E. Apoptosis and free radicals. Biochem. Mol. Med. 1996, 59, 93–97. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef]
- Behnisch-Cornwell, S.; Bandarub, S.S.M.; Napierkowski, M.; Wolff, L.; Zubair, M.; Urbainsky, C.; Lillig, C.; Schulzke, C.; Bednarski, P.J. Pentathiepins: A novel class of glutathione peroxidase 1 inhibitors that induce oxidative stress, loss of mitochondrial membrane potential and apoptosis in human cancer cells. ChemMedChem 2020, 15, 1515–1528. [Google Scholar] [CrossRef]
- Li, C.; Liu, H.; Yang, Y.; Xu, X.; Lv, T.; Zhang, H.; Liu, K.; Zhang, S.; Chen, Y. N-myristoylation of antimicrobial peptide CM4 enhances its anticancer activity by interacting with cell membrane and targeting mitochondria in breast cancer cells. Front. Pharmacol. 2018, 9, 1297. [Google Scholar] [CrossRef]
- Huang, T.-C.; Lee, J.-F.; Chen, J.-Y. Pardaxin, an antimicrobial peptide, triggers caspase-dependent and ROS-mediated apoptosis in HT-1080 cells. Mar. Drugs 2011, 9, 1995–2009. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Wu, Y.; Ma, J.; Yang, J.; Zhang, F. The antibacterial peptide from Bombyx mori cecropinXJ induced growth arrest and apoptosis in human hepatocellular carcinoma cells. Oncol. Lett. 2016, 12, 57–62. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Z.; Hong, D.; Li, S.; Jia, Y. Novel Property Cytotoxicity and Mechanism of Food Preservative Brevilaterins against Human Gastric Cancer Cells. Foods 2023, 12, 1732. https://doi.org/10.3390/foods12081732
Chen Z, Hong D, Li S, Jia Y. Novel Property Cytotoxicity and Mechanism of Food Preservative Brevilaterins against Human Gastric Cancer Cells. Foods. 2023; 12(8):1732. https://doi.org/10.3390/foods12081732
Chicago/Turabian StyleChen, Zhou, Dan Hong, Siting Li, and Yingmin Jia. 2023. "Novel Property Cytotoxicity and Mechanism of Food Preservative Brevilaterins against Human Gastric Cancer Cells" Foods 12, no. 8: 1732. https://doi.org/10.3390/foods12081732