
Antioxidant Activity and Inhibition of Liver Cancer Cells’ Growth of Extracts from 14 Marine Macroalgae Species of the Mediterranean Sea
 , ,
, ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
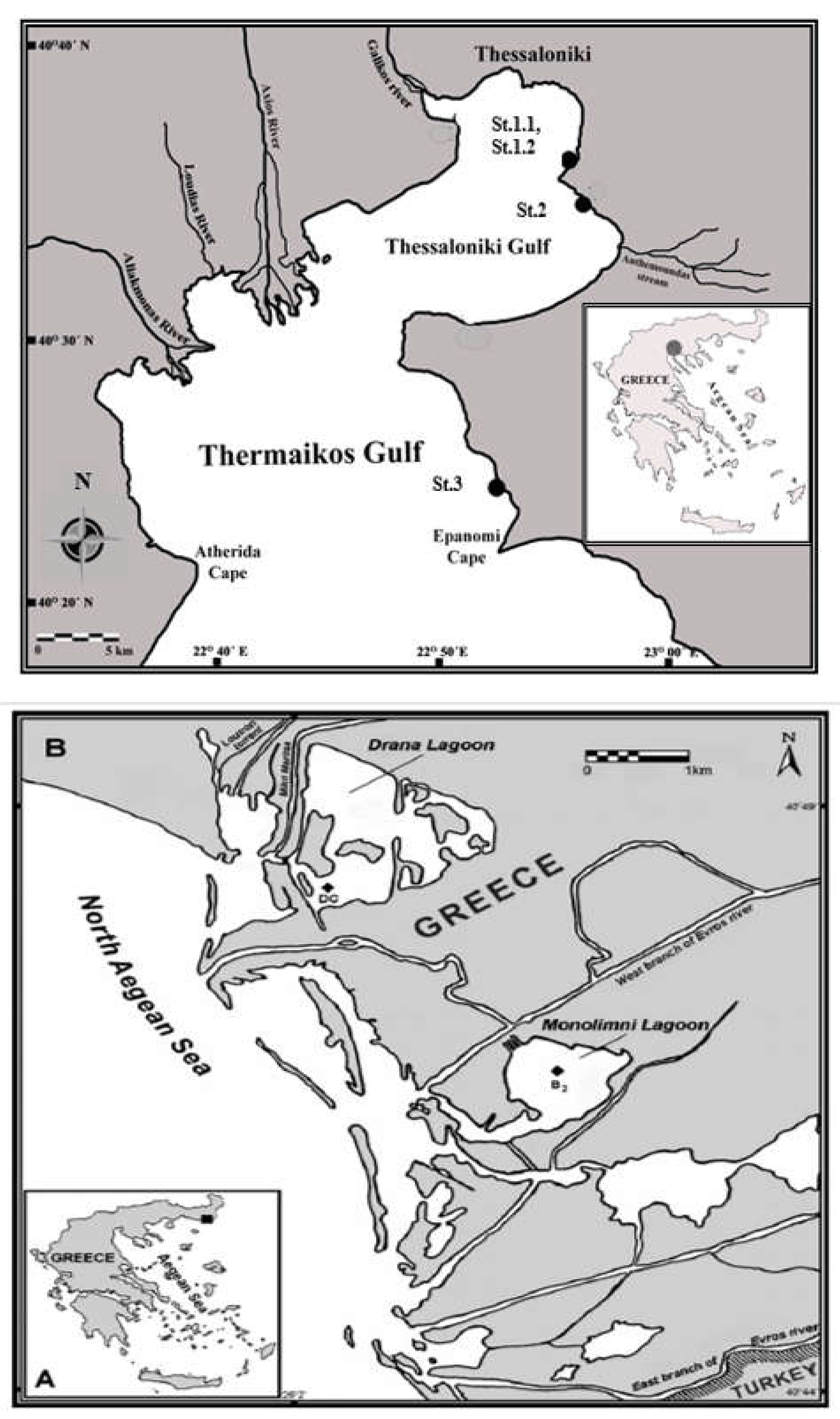
2.1. Marine Macroalgae Species Collection
2.2. Extract Preparation
2.3. Assessment of Macroalgae Extracts’ Polyphenolic Contents
2.4. DPPH Radical Scavenging Assay
2.5. ABTS•+ Radical Scavenging Assay
2.6. Hydroxyl Radical Scavenging Assay
2.7. Superoxide Radical Scavenging Assay
2.8. RP Assay
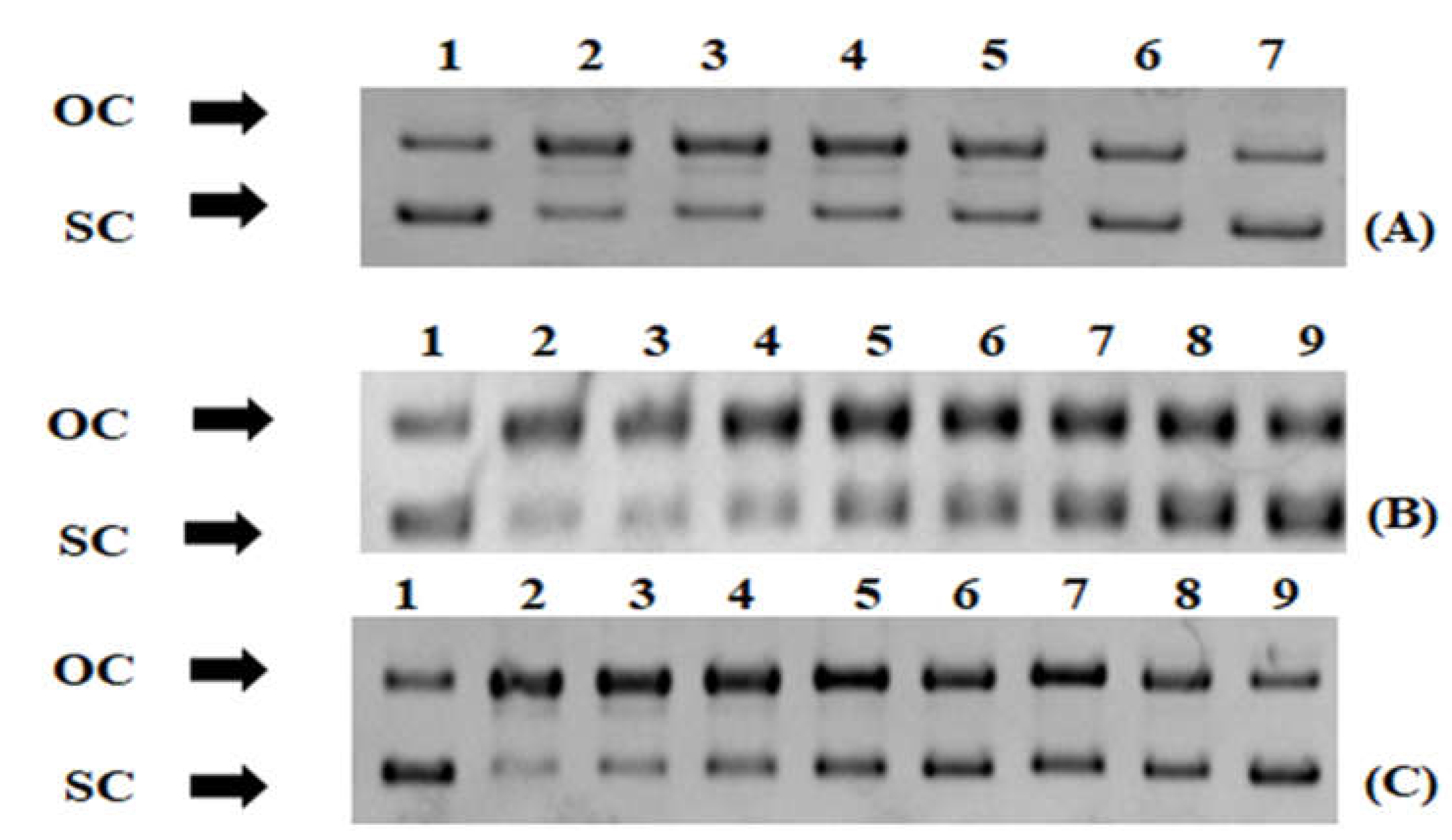
2.9. ROS-Induced DNA Plasmid Strand Cleavage Assay
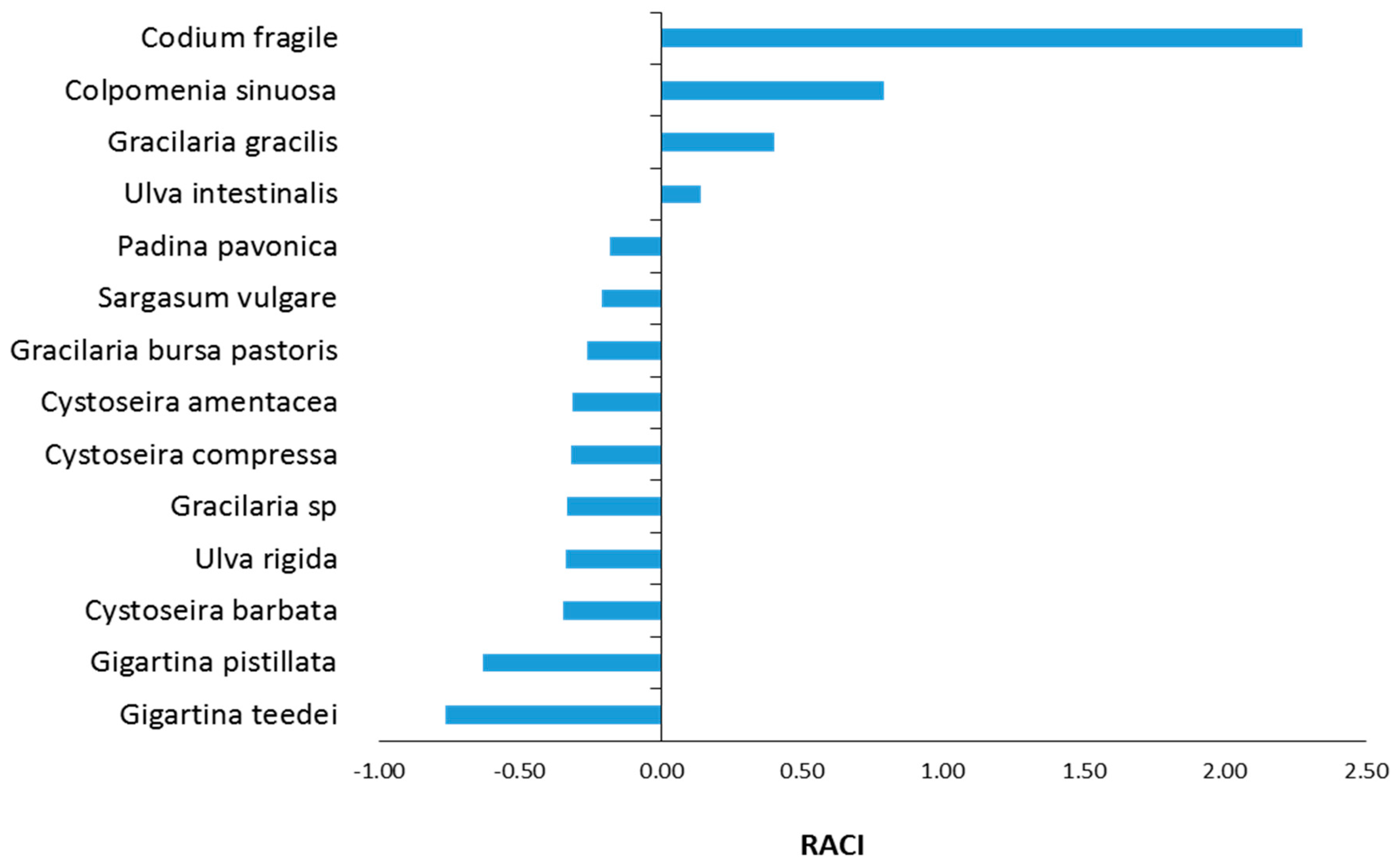
2.10. Evaluation of Relative Antioxidant Capacity Index (RACI)
2.11. Cell Culture Conditions
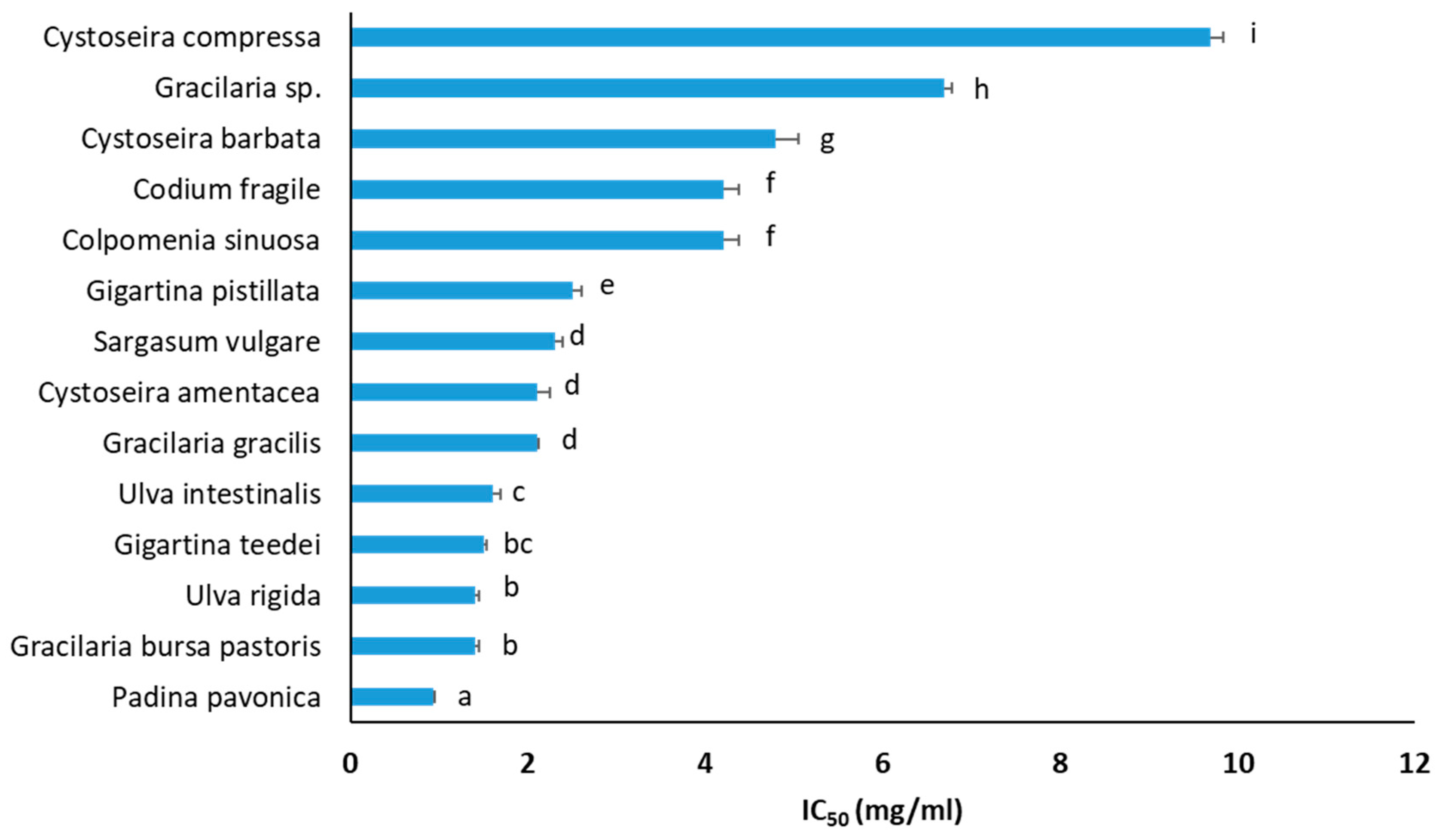
2.12. XTT Assay for Inhibition of Cell Proliferation
2.13. Statistical Analysis
3. Results and Discussion
3.1. Extraction Yield and Assessment of Polyphenolic Content
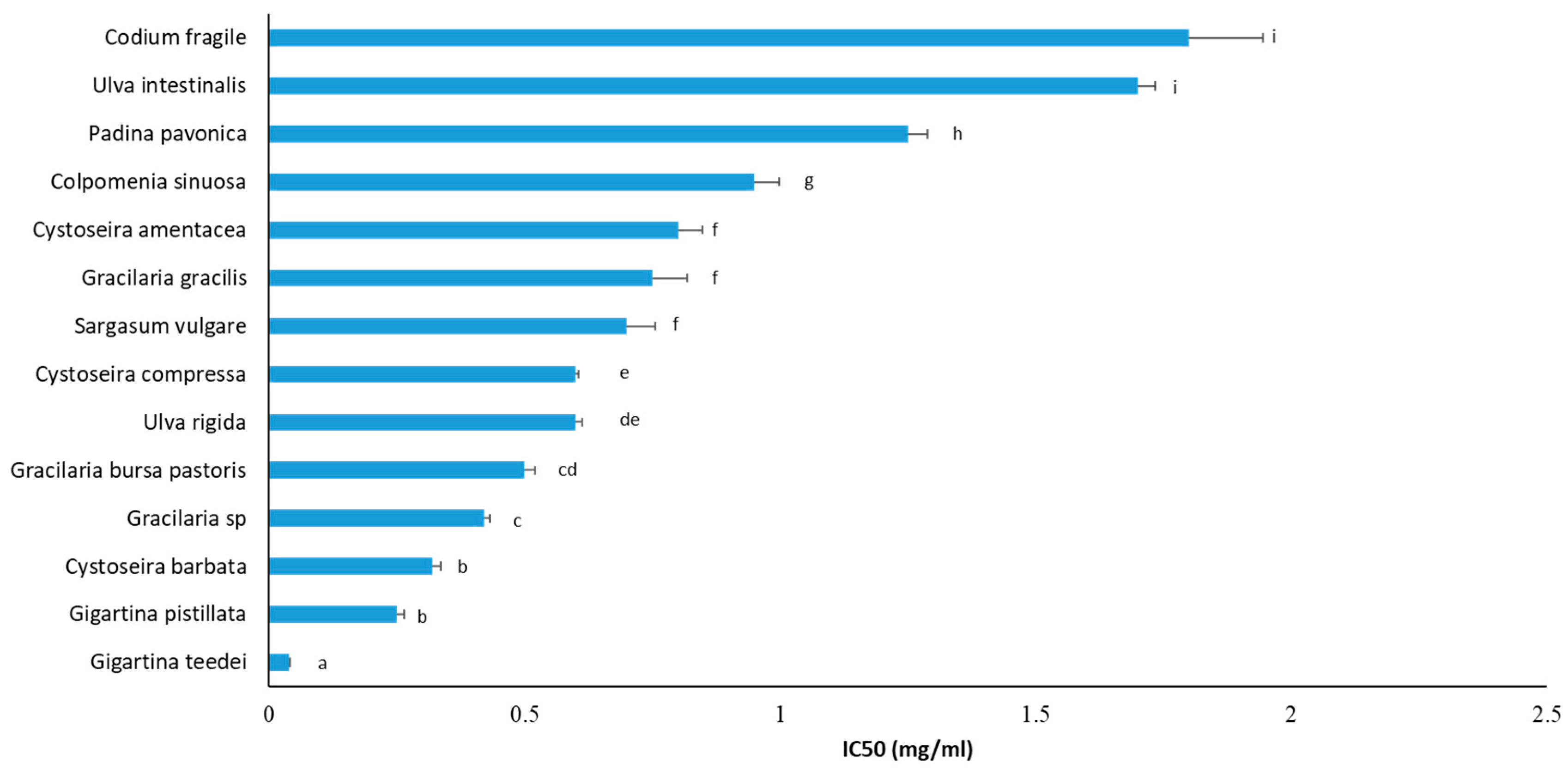
3.2. Free Radical Scavenging Activity
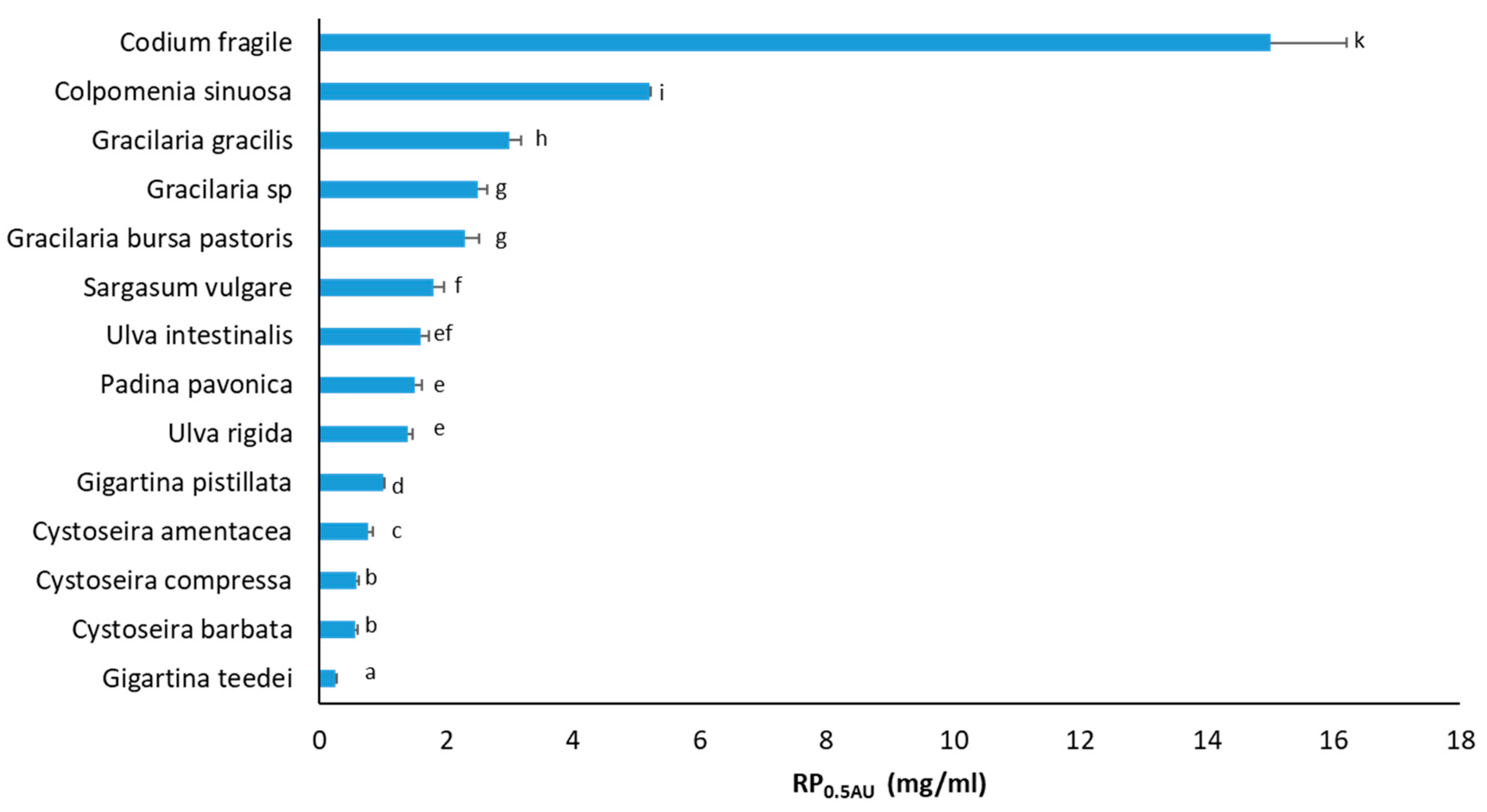
3.3. RP Activity
3.4. Protection from ROS-Induced DNA Damage
3.5. Estimation of RACI Values
3.6. Inhibition of Cancer Cell Growth
3.7. Correlation Analysis
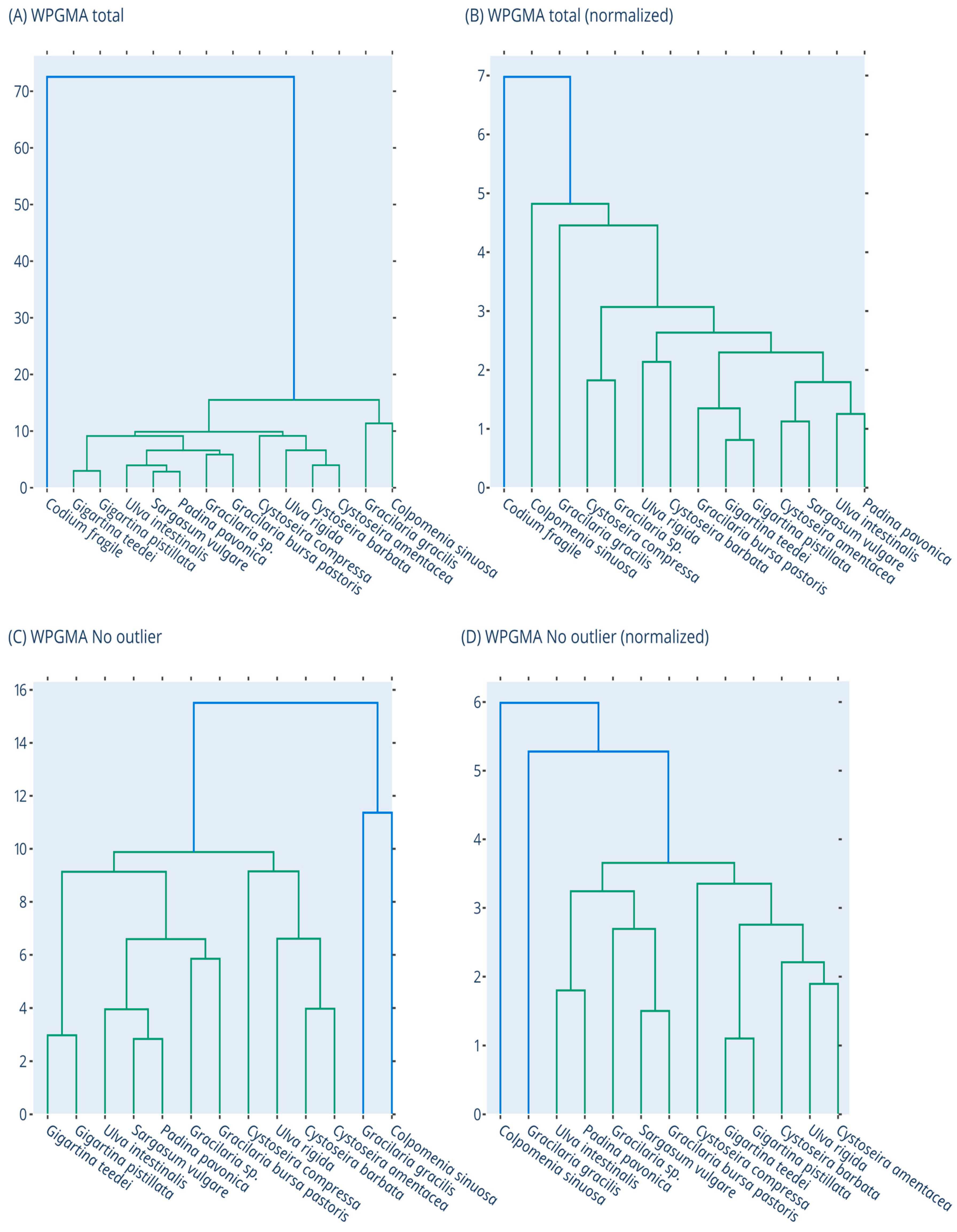
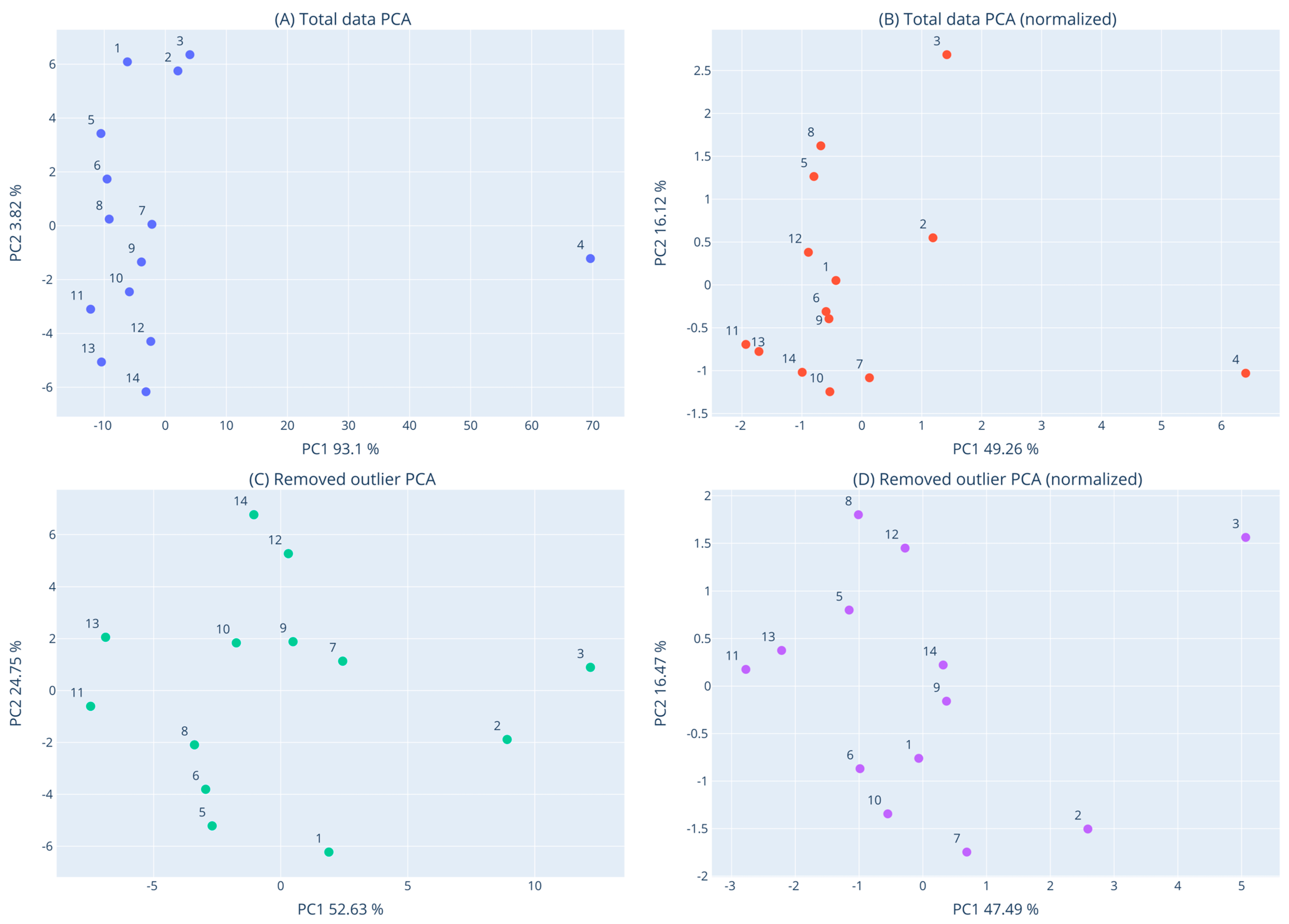
3.8. Clustering of Macroalgae Extracts Based on their Activities with Dendrogram and PCA
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Stagos, D.; Amoutzias, G.D.; Matakos, A.; Spyrou, A.; Tsatsakis, A.M.; Kouretas, D. Chemoprevention of liver cancer by plant polyphenols. Food Chem. Toxicol. 2012, 50, 2155–2170. [Google Scholar] [CrossRef]
- Haque, A.; Brazeau, D.; Amin, A.R. Perspectives on natural compounds in chemoprevention and treatment of cancer: An update with new promising compounds. Eur. J. Cancer 2021, 149, 165–183. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Dai, C. Potential Treatment Strategies for Hepatocellular Carcinoma Cell Sensitization to Sorafenib. J. Hepatocell. Carcinoma 2023, 10, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Correia-da-Silva, M.; Sousa, E.; Pinto, M.; Kijjoa, A. Anticancer and cancer preventive compounds from edible marine organisms. Semin. Cancer Biol. 2017, 46, 55–64. [Google Scholar] [CrossRef]
- Hayes, J.D.; Dinkova-Kostova, A.T.; Tew, K.D. Oxidative Stress in Cancer. Cancer Cell 2020, 38, 167–197. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Hansen, P.E.; Lin, X. Bromophenols in marine algae and their bioactivities. Mar. Drugs 2011, 9, 1273–1292. [Google Scholar] [CrossRef] [Green Version]
- Mateos, R.; Pérez-Correa, J.R.; Domínguez, H. Bioactive Properties of Marine Phenolics. Mar. Drugs 2020, 18, 501. [Google Scholar] [CrossRef]
- Nova, P.; Gomes, A.M.; Costa-Pinto, A.R. It comes from the sea: Macroalgae-derived bioactive compounds with anti-cancer potential. Crit. Rev. Biotechnol. 2023, 1–15. [Google Scholar] [CrossRef]
- Gómez-Guzmán, M.; Rodríguez-Nogales, A.; Algieri, F.; Gálvez, J. Potential Role of Seaweed Polyphenols in Cardiovascular-Associated Disorders. Mar. Drugs 2018, 16, 250. [Google Scholar] [CrossRef] [Green Version]
- Ganesan, A.R.; Tiwari, U.; Rajauria, G. Seaweed nutraceuticals and their therapeutic role in disease prevention. Food Sci. Hum. Wellness 2019, 8, 256–263. [Google Scholar] [CrossRef]
- Gonçalves, A.; Fernandes, M.; Lima, M.; Gomes, J.P.; Silva, F.; Castro, S.; Sampaio, F.; Gomes, A.C. Nanotechnology to the Rescue: Therapeutic Strategies Based on Brown Algae for Neurodegenerative. Dis. Appl. Sci. 2023, 13, 1883. [Google Scholar] [CrossRef]
- Gubelit, Y.I. Opportunistic Macroalgae as a Component in Assessment of Eutrophication. Diversity 2022, 14, 1112. [Google Scholar] [CrossRef]
- Yang, Y.; Hassan, S.H.A.; Awasthi, M.K.; Gajendran, B.; Sharma, M.; Ji, M.-K.; Salama, E.-S. The recent progress on the bioactive compounds from algal biomass for human health applications. Food Biosci. 2023, 51, 102267. [Google Scholar] [CrossRef]
- Pardilhó, S.; Cotas, J.; Pereira, L.; Oliveira, M.B.; Dias, J.M. Marine macroalgae in a circular economy context: A comprehensive analysis focused on residual biomass. Biotechnol. Adv. 2022, 60, 107987. [Google Scholar] [CrossRef] [PubMed]
- Healy, L.E.; Zhu, X.; Pojić, M.; Sullivan, C.; Tiwari, U.; Curtin, J.; Tiwari, B.K. Biomolecules from Macroalgae-Nutritional Profile and Bioactives for Novel Food Product Development. Biomolecules 2023, 13, 386. [Google Scholar] [CrossRef] [PubMed]
- Malea, P.; Chatziapostolou, A.; Kevrekidis, T. Trace element seasonality in marine macroalgae of different functional-form groups. Mar. Environ. Res. 2015, 103, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Malea, P.; Kevrekidis, T. Trace element patterns in marine macroalgae. Sci. Total Environ. 2014, 494–495, 144–157. [Google Scholar] [CrossRef]
- Guo, J.; Qi, M.; Chen, H.; Zhou, C.; Ruan, R.; Yan, X.; Cheng, P. Macroalgae-Derived Multifunctional Bioactive Substances: The Potential Applications for Food and Pharmaceuticals. Foods 2022, 11, 3455. [Google Scholar] [CrossRef]
- Armeli Minicante, S.; Bongiorni, L.; De Lazzari, A. Bio-Based Products from Mediterranean Seaweeds: Italian Opportunities and Challenges for a Sustainable Blue Economy. Sustainability 2022, 14, 5634. [Google Scholar] [CrossRef]
- Tsiamis, K.; Verlaque, M.; Panayotidis, P.; Montesanto, B. New macroalgal records for the Aegean Sea (Greece, eastern Mediterranean Sea). Bot. Mar. 2010, 53, 319–331. [Google Scholar] [CrossRef]
- Taşkln, E.; Çaklr, M. Marine macroalgal flora on the Aegean and the Levantine coasts of Turkey. Bot. Mar. 2022, 65, 231–241. [Google Scholar] [CrossRef]
- Papathanassiou, E.; Zenetos, A. (Eds.) State of the Hellenic Marine Environment; Hellenic Center for Marine Research: Athens, Greece, 2005; p. 360. [Google Scholar]
- Tsiamis, K.; Panayotidis, P.; Economou-Amilli, A.; Katsaros, C. Seaweeds of the Greek coasts. I. Phaeophyceae. Medit. Mar. Sci. 2013, 14, 141–157. [Google Scholar] [CrossRef]
- Tsiamis, K.; Panayotidis, P.; Economou-Amilli, A.; Katsaros, C. Seaweeds of the Greek coasts. II. Ulvophyceae. Medit. Mar. Sci. 2014, 15, 449–461. [Google Scholar] [CrossRef] [Green Version]
- Montalvão, S.; Demirel, Z.; Devi, P.; Lombardi, V.; Hongisto, V.; Perälä, M.; Hattara, J.; Imamoglu, E.; Tilvi, S.S.; Turan, G.; et al. Large-scale bioprospecting of cyanobacteria, micro- and macroalgae from the Aegean Sea. N. Biotechnol. 2016, 33, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Demirel, Z.; Yilmaz-Koz, F.F.; Karabay-Yavasoglu, U.N.; Ozdemir, G.; Sukatar, A. Antimicrobial and antioxidant activity of brown algae from the Aegean Sea. J. Serb. Chem. Soc. 2009, 74, 619–628. [Google Scholar] [CrossRef]
- Güner, A.; Köksal, Ç.; Erel, Ş.B.; Kayalar, H.; Nalbantsoy, A.; Sukatar, A.; Karabay Yavaşoğlu, N.Ü. Antimicrobial and antioxidant activities with acute toxicity, cytotoxicity and mutagenicity of Cystoseira compressa (Esper) Gerloff & Nizamuddin from the coast of Urla (Izmir, Turkey). Cytotechnology 2015, 67, 135–143. [Google Scholar] [PubMed] [Green Version]
- Boubonari, T.; Malea, P.; Kevrekidis. The green seaweed Ulva rigida as a bioindicator of metals (Zn, Cu, Pb and Cd) in a low-salinity coastal environment. Bot. Mar. 2008, 51, 472–484. [Google Scholar] [CrossRef]
- Pell, A.; Kokkinis, G.; Malea, P.; Pergantis, S.A.; Rubio, R.; López-Sánchez, J.F. LC-ICP-MS analysis of arsenic compounds in dominant seaweeds from the Thermaikos Gulf (Northern Aegean Sea, Greece). Chemosphere 2013, 93, 2187–2194. [Google Scholar] [CrossRef] [Green Version]
- Verlaque, M.; Ruitton, S.; Mineur, F.; Boudouresque, C.F. CIESM Atlas of Exotic Species of the Mediterranean. Volume 4: Macrophytes; Briand, F., Ed.; CIESM: Monaco City, Monaco, 2015; p. 362. [Google Scholar]
- Gómez Garreta, A.; Gallardo, T.; Ribera, M.A.; Cormaci, M.; Furnari, G.; Giaccone, G.; Boudouresque, C.F. Checklist of Mediterranean seaweeds. III. Rhodophyceae rabenh. 1. Ceramiales oltm. Bot. Mar. 2001, 44, 425–460. [Google Scholar] [CrossRef]
- Gallardo, T.; Garreta, A.G.; Ribera, M.A.; Cormaci, M.; Furnari, G.; Giaccone, G.; Boudouresque, C.F. Check-list of Mediterranean Seaweeds II. Chlorophyceae Wille s. l. Bot. Mar. 1993, 36, 399–422. [Google Scholar] [CrossRef]
- Ribera, M.A.; Garreta, A.G.; Gallardo, T.; Cormaci, M.; Furnari, G.; Giaccone, G. Check-list of Mediterranean Seaweeds: I. Fucophyceae (Warming, 1884). Bot. Mar. 1992, 35, 109–130. [Google Scholar] [CrossRef]
- Frick, H.G.; Boudouresque, C.F.; Verlaque, M. A checklist of marine algae of the Lavezzi Archipelago, with special attention to species rare or new to Corsica (Mediterranean). Nova Hedwig. 1996, 62, 119–135. [Google Scholar]
- Benhissoune, S.; Boudouresque, C.F.; Verlaque, M. A checklist of the seaweeds of the Mediterranean and Atlantic coasts of Morocco. II. Phaeophyceae. Bot. Mar. 2002, 45, 217–230. [Google Scholar] [CrossRef]
- Benhissoune, S.; Boudouresque, C.F.; Perret-Boudouresque, M.; Verlaque, M. A checklist of the seaweeds of the Mediterranean and Atlantic coasts of Morocco. III. Rhodophyceae (excluding Ceramiales). Bot. Mar. 2002, 45, 391–412. [Google Scholar]
- Benhissoune, S.; Boudouresque, C.F.; Perret-Boudouresque, M.; Verlaque, M. A checklist of the seaweeds of the Mediterranean and Atlantic coasts of Morocco. IV. Rhodophyceae—Ceramiales. Bot. Mar. 2003, 46, 55–68. [Google Scholar] [CrossRef]
- South, G.R.; Titley, I. A Checklist and Distributional Index of the Benthic Marine Algae of the North Atlantic Ocean; British Museum (Natural History): London, UK, 1986; p. 76. [Google Scholar]
- Burrows, E.M. Seaweeds of the British Isles. Volume 2. Chlorophyta; Natural History Museum Publications: London, UK, 1991. [Google Scholar]
- AlgaeBase. Available online: https://www.algaebase.org/search/species/ (accessed on 13 November 2022).
- de Los Santos, C.B.; Pérez-Lloréns, J.L.; Vergara, J.J. Photosynthesis and growth in macroalgae: Linking functional-form and power-scaling approaches. Mar. Ecol. Prog. Ser. 2009, 377, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Farasat, M.; Khavari-Nejad, R.A.; Nabavi, S.M.; Namjooyan, F. Antioxidant Properties of two Edible Green Seaweeds From Northern Coasts of the Persian Gulf. Jundishapurr J. Nat. Pharm. Prod. 2013, 8, 47–52. [Google Scholar] [CrossRef] [Green Version]
- Apostolou, A.; Stagos, D.; Galitsiou, E.; Spyrou, A.; Haroutounian, S.; Portesis, N.; Trizoglou, I.; Wallace Hayes, A.; Tsatsakis, A.M.; Kouretas, D. Assessment of polyphenolic content, antioxidant activity, protection against ROS-induced DNA damage and anticancer activity of Vitis vinifera stem extracts. Food Chem. Toxicol. 2013, 61, 60–68. [Google Scholar] [CrossRef]
- Kerasioti, E.; Stagos, D.; Priftis, A.; Aivazidis, S.; Tsatsakis, A.M.; Hayes, A.W.; Kouretas, D. Antioxidant effects of whey protein on muscle C2C12 cells. Food Chem. 2014, 155, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Priftis, A.; Mitsiou, D.; Halabalaki, M.; Ntasi, G.; Stagos, D.; Skaltsounis, L.A.; Kouretas, D. Roasting has a distinct effect on the antimutagenic activity of coffee varieties. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2018, 829–830, 33–42. [Google Scholar] [CrossRef]
- Kreatsouli, K.; Fousteri, Z.; Zampakas, K.; Kerasioti, E.; Veskoukis, A.S.; Mantas, C.; Gkoutsidis, P.; Ladas, D.; Petrotos, K.; Kouretas, D.; et al. A Polyphenolic Extract from Olive Mill Wastewaters Encapsulated in Whey Protein and Maltodextrin Exerts Antioxidant Activity in Endothelial Cells. Antioxidants 2019, 8, 280. [Google Scholar] [CrossRef] [Green Version]
- Virtanen, P.; Gommers, R.; Oliphant, T.E.; Haberland, M.; Reddy, T.; Cournapeau, D.; Burovski, E.; Peterson, P.; Weckesser, W.; Bright, J.; et al. SciPy 1.0 Contributors. SciPy 1.0: Fundamental algorithms for scientific computing in Python. Nat. Methods 2020, 17, 261–272. [Google Scholar] [CrossRef] [Green Version]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine Learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Sultana, B.; Anwar, F.; Ashraf, M. Effect of Extraction Solvent/Technique on the Antioxidant Activity of Selected Medicinal Plant Extracts. Molecules 2009, 14, 2167–2180. [Google Scholar] [CrossRef] [Green Version]
- Sapatinha, M.; Oliveira, A.; Costa, S.; Pedro, S.; Gonçalves, A.; Mendes, R.; Bandarra, N.M.; Pires, C. Red and brown seaweeds extracts: A source of biologically active compounds. Food Chem. 2022, 393, 133453. [Google Scholar] [CrossRef]
- De La Fuente, G.; Fontana, M.; Asnaghi, V.; Chiantore, M.; Mirata, S.; Salis, A.; Damonte, G.; Scarfì, S. The Remarkable Antioxidant and Anti-Inflammatory Potential of the Extracts of the Brown Alga Cystoseira amentacea var. stricta. Mar. Drugs 2020, 19, 2. [Google Scholar] [CrossRef]
- Trifan, A.; Vasincu, A.; Luca, S.V.; Neophytou, C.; Wolfram, E.; Opitz, S.; Sava, D.; Bucur, L.; Cioroiu, B.I.; Miron, A.; et al. Unravelling the potential of seaweeds from the Black Sea coast of Romania as bioactive compounds sources. Part I: Cystoseira barbata (Stackhouse) C. Agardh. Food Chem. Toxicol. 2019, 134, 110820. [Google Scholar] [CrossRef]
- Francavilla, M.; Franchi, M.; Monteleone, M.; Caroppo, C. The red seaweed Gracilaria gracilis as a multi products source. Mar. Drugs 2013, 11, 3754–3776. [Google Scholar] [CrossRef] [Green Version]
- Caf, F.; Yilmaz, Ö.; Durucan, F.; Şen Özdemir, N. 2015. Biochemical components of three marine macroalgae (Padina pavonica, Ulva lactuca and Taonia atomaria) from the Levantine Sea Coast of Antalya, Turkey. JBES 2015, 6, 401–411. [Google Scholar]
- Bernardini, G.; Minetti, M.; Polizzotto, G.; Biazzo, M.; Santucci, A. Pro-Apoptotic Activity of French Polynesian Padina pavonica Extract on Human Osteosarcoma Cells. Mar. Drugs 2018, 16, 504. [Google Scholar] [CrossRef] [Green Version]
- De La Fuente, G.; Pinteus, S.; Silva, J.; Alves, C.; Pedrosa, R. Antioxidant and antimicrobial potential of six Fucoids from the Mediterranean Sea and the Atlantic Ocean. J. Sci. Food Agric. 2022, 102, 5568–5575. [Google Scholar] [CrossRef]
- Čagalj, M.; Skroza, D.; Razola-Díaz, M.; Verardo, V.; Bassi, D.; Frleta, R.; Generalić Mekinić, I.; Tabanelli, G.; Šimat, V. Variations in the Composition, Antioxidant and Antimicrobial Activities of Cystoseira compressa during Seasonal Growth. Mar. Drugs 2022, 20, 64. [Google Scholar] [CrossRef]
- Dang, T.T.; Bowyer, M.C.; van Altena, I.A.; Scarlett, C.J. Comparison of chemical profile and antioxidant properties of the brown algae. Int. J. Food Sci. Technol. 2018, 53, 174–181. [Google Scholar] [CrossRef]
- Jormalainen, V.; Wikström, S.A.; Honkanen, T. Fouling mediates grazing: Intertwining of resistances to multiple enemies in the brown alga Fucus vesiculosus. Oecologia 2008, 155, 559–569. [Google Scholar] [CrossRef]
- Martins, A.; Alves, C.; Silva, J.; Pinteus, S.; Gaspar, H.; Pedrosa, R. Sulfated Polysaccharides from Macroalgae-A Simple Roadmap for Chemical Characterization. Polymers 2023, 15, 399. [Google Scholar] [CrossRef]
- Neto, R.T.; Marçal, C.; Queirós, A.S.; Abreu, H.; Silva, A.; Cardoso, S.M. Screening of Ulva rigida, Gracilaria sp. Fucus vesiculosus and Saccharina latissima as Functional Ingredients. Int. J. Mol. Sci. 2018, 19, 2987. [Google Scholar]
- Mezghani, S.; Bourguiba, I.; Hfaiedh, I.; Amri, M. Antioxidant potential of Ulva rigida extracts: Protection of HeLa cells against H2O2 cytotoxicity. Biol. Bull. 2013, 225, 1–7. [Google Scholar] [CrossRef]
- Tolpeznikaite, E.; Starkute, V.; Zokaityte, E.; Ruzauskas, M.; Pilkaityte, R.; Viskelis, P.; Urbonaviciene, D.; Ruibys, R.; Rocha, J.M.; Bartkiene, E. Effect of solid-state fermentation and ultrasonication processes on antimicrobial and antioxidant properties of algae extracts. Front. Nutr. 2022, 9, 990274. [Google Scholar] [CrossRef]
- Wekre, M.E.; Kåsin, K.; Underhaug, J.; Holmelid, B.; Jordheim, M. Quantification of Polyphenols in Seaweeds: A Case Study of Ulva intestinalis. Antioxidants 2019, 8, 612. [Google Scholar] [CrossRef] [Green Version]
- Heffernan, N.; Smyth, T.J.; Soler-Villa, A.; Fitzgerald, R.J.; Brunton, N.P. Phenolic content and antioxidant activity of fractions obtained from selected Irish macroalgae species (Laminaria digitata, Fucus serratus, Gracilaria gracilis and Codium fragile). J. Appl. Phycol. 2015, 27, 519–530. [Google Scholar] [CrossRef]
- Santos, S.A.; Félix, R.; Pais, A.C.; Rocha, S.M.; Silvestre, A.J. The Quest for Phenolic Compounds from Macroalgae: A Review of Extraction and Identification Methodologies. Biomolecules 2019, 9, 847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milović, S.; Stanković, I.; Nikolić, D.; Radović, J.; Kolundžić, M.; Nikolić, V.; Stanojković, T.; Petović, S.; Kundaković-Vasović, T. Chemical Analysis of Selected Seaweeds and Seagrass from the Adriatic Coast of Montenegro. Chem Biodivers. 2019, 16, e1900327. [Google Scholar] [CrossRef]
- Yuan, Y.; Zheng, Y.; Zhou, J.; Geng, Y.; Zou, P.; Li, Y.; Zhang, C. Polyphenol-Rich Extracts from Brown Macroalgae Lessonia trabeculate Attenuate Hyperglycemia and Modulate Gut Microbiota in High-Fat Diet and Streptozotocin-Induced Diabetic Rats. J. Agric. Food Chem. 2019, 67, 12472–12480. [Google Scholar] [CrossRef]
- Aminina, N.M.; Karaulova, E.P.; Vishnevskaya, T.I.; Yakush, E.V.; Kim, Y.K.; Nam, K.H.; Son, K.T. Characteristics of Polyphenolic Content in Brown Algae of the Pacific Coast of Russia. Molecules 2020, 25, 3909. [Google Scholar] [CrossRef] [PubMed]
- Semaida, A.I.; El-Khashab, M.A.; Saber, A.A.; Hassan, A.I.; Elfouly, S.A. Effects of Sargassum virgatum extracts on the testicular measurements, genomic DNA and antioxidant enzymes in irradiated rats. Int. J. Radiat. Biol. 2022, 98, 191–204. [Google Scholar] [CrossRef] [PubMed]
- Generalić Mekinić, I.; Šimat, V.; Botić, V.; Crnjac, A.; Smoljo, M.; Soldo, B.; Ljubenkov, I.; Čagalj, M.; Skroza, D. Bioactive Phenolic Metabolites from Adriatic Brown Algae Dictyota dichotoma and Padina pavonica (Dictyotaceae). Foods 2021, 10, 1187. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Long, X.; Liu, P.S.; Xie, X. The interplay of oncogenic signaling, oxidative stress and ferroptosis in cancer. Int. J. Cancer, 2023; ahead of print. [Google Scholar] [CrossRef]
- Tsukatani, T.; Ide, S.; Ono, M.; Matsumoto, K. New tetrazolium method for phosphatase assay using ascorbic acid 2-phosphate as a substrate. Talanta 2007, 73, 471–475. [Google Scholar] [CrossRef]
- Zubia, M.; Robledo, D.; Freile-Pelegrin, Y. Antioxidant activities in tropical marine macroalgae from the Yucatan Peninsula, Mexico. J. Appl. Phycol. 2007, 19, 449–458. [Google Scholar] [CrossRef]
- Kosanić, M.; Ranković, B.; Stanojković, T. Biological potential of marine macroalgae of the genus Cystoseira. Acta Biol. Hung. 2015, 66, 374–384. [Google Scholar] [CrossRef] [Green Version]
- Mhadhebi, L.; Mhadhebi, A.; Robert, J.; Bouraoui, A. Antioxidant, Anti-inflammatory and Antiproliferative Effects of Aqueous Extracts of Three Mediterranean Brown Seaweeds of the Genus Cystoseira. Iran. J. Pharm. Res. 2014, 13, 207–220. [Google Scholar]
- Al-Enazi, N.M.; Awaad, A.S.; Zain, M.E.; Alqasoumi, S.I. Antimicrobial, antioxidant and anticancer activities of Laurencia catarinensis, Laurencia majuscula and Padina pavonica extracts. Saudi Pharm. J. 2018, 26, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Chouh, A.; Nouadri, T.; Catarino, M.D.; Silva, A.; Cardoso, S.M. Phlorotannins of the Brown Algae Sargassum vulgare from the Mediterranean Sea Coast. Antioxidants 2022, 11, 1055. [Google Scholar] [CrossRef] [PubMed]
- Apak, R.; Özyürek, M.; Güçlü, K.; Çapanoğlu, E. Antioxidant Activity/Capacity Measurement. 2. Hydrogen Atom Transfer (HAT)-Based, Mixed-Mode (Electron Transfer (ET)/HAT), and Lipid Peroxidation Assays. J. Agric. Food Chem. 2016, 64, 1028–1045. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Reactive oxygen species (ROS), oxygen radicals and antioxidants: Where are we now, where is the field going and where should we go? Biochem. Biophysic. Res. Communic. 2022, 633, 17–19. [Google Scholar] [CrossRef]
- Liu, Z.Q. Enhancing Antioxidant Effect against Peroxyl Radical-Induced Oxidation of DNA: Linking with Ferrocene Moiety! Chem. Rec. 2019, 19, 2385–2397. [Google Scholar] [CrossRef] [PubMed]
- Sellimi, S.; Younes, I.; Ayed, H.B.; Maalej, H.; Montero, V.; Rinaudo, M.; Dahia, M.; Mechichi, T.; Hajji, M.; Nasri, M. Structural, physicochemical and antioxidant properties of sodium alginate isolated from a Tunisian brown seaweed. Int. J. Biol. Macromol. 2015, 72, 1358–1367. [Google Scholar] [CrossRef] [PubMed]
- Celikler, S.; Tas, S.; Ziyanok-Ayvalik, S.; Vatan, O.; Yildiz, G.; Ozel, M. Protective antigenotoxic effect of Ulva rigida, C. Agardh in experimental hypothyroid. Acta Biol. Hung. 2014, 65, 13–26. [Google Scholar] [CrossRef]
- Alhmoud, J.F.; Woolley, J.F.; Al Moustafa, A.E.; Malki, M.I. DNA Damage/Repair Management in Cancers. Cancers 2020, 12, 1050. [Google Scholar] [CrossRef]
- Cotas, J.; Leandro, A.; Monteiro, P.; Pacheco, D.; Figueirinha, A.; Gonçalves, A.; da Silva, G.J.; Pereira, L. Seaweed Phenolics: From Extraction to Applications. Mar. Drugs 2020, 18, 384. [Google Scholar] [CrossRef]
- Dini, I.; Grumetto, L. Recent Advances in Natural Polyphenol Research. Molecules 2022, 27, 8777. [Google Scholar] [CrossRef]
- Carpena Rodríguez, M.; Caleja, C.; Pereira, E.; Pereira, C.; Ćirić, A.; Soković, M.; Soria Lopez, A.; Fraga Corral, M.; Simal Gándara, J.; Ferreira, I.C.; et al. Red Seaweeds as a Source of Nutrients and Bioactive Compounds: Optimization of the Extraction. Chemosensors 2021, 9, 132. [Google Scholar] [CrossRef]
- Carpena, M.; Garcia-Perez, P.; Garcia-Oliveira, P.; Chamorro, F.; Otero, P.; Lourenço-Lopes, C.; Cao, H.; Simal-Gandara, J.; Prieto, M.A. Biological properties and potential of compounds extracted from red seaweeds. Phytochem. Rev. 2022, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Al Monla, R.; Dassouki, Z.; Sari-Chmayssem, N.; Mawlawi, H.; Gali-Muhtasib, H. Fucoidan and Alginate from the Brown Algae Colpomenia sinuosa and Their Combination with Vitamin C Trigger Apoptosis in Colon Cancer. Molecules 2022, 27, 358. [Google Scholar] [CrossRef] [PubMed]
- Sellimi, S.; Maalej, H.; Rekik, D.M.; Benslima, A.; Ksouda, G.; Hamdi, M.; Sahnoun, Z.; Li, S.; Nasri, M.; Hajji, M. Antioxidant, antibacterial and in vivo wound healing properties of laminaran purified from Cystoseira barbata seaweed. Int. J. Biol. Macromol. 2018, 119, 633–644. [Google Scholar] [CrossRef] [PubMed]
- Sellimi, S.; Benslima, A.; Barragan-Montero, V.; Hajji, M.; Nasri, M. Polyphenolic-protein-polysaccharide ternary conjugates from Cystoseira barbata Tunisian seaweed as potential biopreservatives: Chemical, antioxidant and antimicrobial properties. Int. J. Biol. Macromol. 2017, 105 Pt 2, 1375–1383. [Google Scholar] [CrossRef]
- Rocha de Souza, M.C.; Marques, C.T.; Guerra Dore, C.M.; Ferreira da Silva, F.R.; Oliveira Rocha, H.A.; Leite, E.L. Antioxidant activities of sulfated polysaccharides from brown and red seaweeds. J. Appl. Phycol. 2007, 19, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Ruberto, G.; Baratta, M.T.; Biondi, D.M. Antioxidant activity of extracts of the marine algal genus Cystoseira in a micellar model system. J. Appl. Phycol. 2001, 13, 403–407. [Google Scholar] [CrossRef]
- Pereira, L. Population studies and carrageenan properties in eight Gigartinales (Rhodophyta) from Western Coast of Portugal. Sci. World J. 2013, 2013, 939830. [Google Scholar] [CrossRef] [Green Version]
- Olasehinde, T.A.; Mabinya, L.V.; Olaniran, A.O.; Okoh, A.I. Chemical characterization of sulfated polysaccharides from Gracilaria gracilis and Ulva lactuca and their radical scavenging, metal chelating, and cholinesterase inhibitory activities. Int. J. Food Prop. 2019, 22, 100–110. [Google Scholar] [CrossRef] [Green Version]
- El Rashed, Z.; Lupidi, G.; Grasselli, E.; Canesi, L.; Khalifeh, H.; Demori, I. Antioxidant and Antisteatotic Activities of Fucoidan Fractions from Marine and Terrestrial Sources. Molecules 2021, 26, 4467. [Google Scholar] [CrossRef]
- Wang, L.; Oh, J.; Je, J.; Jayawardena, T.U.; Kim, Y.; Ko, J.Y.; Fu, X.; Jeon, Y. Protective effects of sulfated polysaccharides isolated from the enzymatic digest of Codium fragile against hydrogen peroxide-induced oxidative stress in in vitro and in vivo models. Algal Res. 2020, 48, 101891. [Google Scholar] [CrossRef]
- Pereira, T.; Barroso, S.; Mendes, S.; Amaral, R.A.; Dias, J.R.; Baptista, T.; Saraiva, J.A.; Alves, N.M.; Gil, M.M. Optimization of phycobiliprotein pigments extraction from red algae Gracilaria gracilis for substitution of synthetic food colorants. Food Chem. 2020, 321, 126688. [Google Scholar] [CrossRef]
- Banerjee, K.; Ghosh, R.; Homechaudhuri, S.; Mitra, A. Biochemical Composition of Marine Macroalgae from Gangetic Delta at the Apex of Bay of Bengal. African J. Basic Appl. Sci. 2009, 1, 96–104. [Google Scholar]
- Al Monla, R.; Dassouki, Z.; Kouzayha, A.; Salma, Y.; Gali-Muhtasib, H.; Mawlawi, H. The Cytotoxic and Apoptotic Effects of the Brown Algae Colpomenia sinuosa are Mediated by the Generation of Reactive Oxygen Species. Molecules 2020, 25, 1993. [Google Scholar] [CrossRef]
- El-Sheekh, M.M.; Nassef, M.; Bases, E.; Shafay, S.E.; El-Shenody, R. Antitumor immunity and therapeutic properties of marine seaweeds-derived extracts in the treatment of cancer. Cancer Cell Int. 2022, 22, 267. [Google Scholar] [CrossRef]
- Guerra Dore, C.M.; Faustino Alves, M.G.; Santos, N.D.; Cruz, A.K.; Câmara, R.B.; Castro, A.J.; Guimarães Alves, L.; Nader, H.B.; Leite, E.L. Antiangiogenic activity and direct antitumor effect from a sulfated polysaccharide isolated from seaweed. Microvasc. Res. 2013, 88, 12–18. [Google Scholar] [CrossRef]
- Kim, A.D.; Lee, Y.; Kang, S.H.; Kim, G.Y.; Kim, H.S.; Hyun, J.W. Cytotoxic effect of clerosterol isolated from Codium fragile on A2058 human melanoma cells. Mar. Drugs 2013, 11, 418–430. [Google Scholar] [CrossRef] [Green Version]
- Park, H.B.; Hwang, J.; Zhang, W.; Go, S.; Kim, J.; Choi, I.; You, S.; Jin, J.O. Polysaccharide from Codium fragile Induces Anti-Cancer Immunity by Activating Natural Killer Cells. Mar. Drugs 2020, 18, 626. [Google Scholar] [CrossRef]
- Wang, Y.; An, E.K.; Kim, S.J.; You, S.; Jin, J.O. Intranasal Administration of Codium fragile Polysaccharide Elicits Anti-Cancer Immunity against Lewis Lung Carcinoma. Int. J. Mol. Sci. 2021, 22, 10608. [Google Scholar] [CrossRef]
- Jin, J.O.; Yadav, D.; Madhwani, K.; Puranik, N.; Chavda, V.; Song, M. Seaweeds in the Oncology Arena: Anti-Cancer Potential of Fucoidan as a Drug-A Review. Molecules 2022, 27, 6032. [Google Scholar] [CrossRef]
- Nazarudin, M.F.; Isha, A.; Mastuki, S.N.; Ain, N.M.; Mohd Ikhsan, N.F.; Abidin, A.Z.; Aliyu-Paiko, M. Chemical Composition and Evaluation of the α-Glucosidase Inhibitory and Cytotoxic Properties of Marine Algae Ulva intestinalis, Halimeda macroloba, and Sargassum ilicifolium. Evid. Based Complement. Alternat. Med. 2020, 2020, 2753945. [Google Scholar] [CrossRef] [PubMed]
- Pal, A.; Verma, P.; Paul, S.; Majumder, I.; Kundu, R. Two species of Ulva inhibits the progression of cervical cancer cells SiHa by means of autophagic cell death induction. 3 Biotech 2021, 11, 52. [Google Scholar] [CrossRef]
- Lopes, D.; Melo, T.; Rey, F.; Meneses, J.; Monteiro, F.L.; Helguero, L.A.; Abreu, M.H.; Lillebø, A.I.; Calado, R.; Domingues, M.R. Valuing Bioactive Lipids from Green, Red and Brown Macroalgae from Aquaculture, to Foster Functionality and Biotechnological Applications. Molecules 2020, 25, 3883. [Google Scholar] [CrossRef] [PubMed]
- Cotas, J.; Marques, V.; Afonso, M.B.; Rodrigues, C.; Pereira, L. Antitumour Potential of Gigartina pistillata Carrageenans against Colorectal Cancer Stem Cell-Enriched Tumourspheres. Mar. Drugs 2020, 18, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Stations | Taxonomic Group (Phylum/Classis) | Functional-Form Group |
|---|---|---|---|
| Codium fragile subsp. fragile (Suringar) Hariot | 1.1 | Chlorophyta/Ulvophyceae | Coarsely-branched |
| Ulva intestinalis Linnaeus | 1.1 | Sheet | |
| Ulva rigida C. Agardh | 1.2 | Sheet | |
| Colpomenia sinuosa var. peregrina Sauvageau | 1.1 | Ochrophyta/Phaeophyceae | Coarsely-branched |
| Cystoseira barbata (Stackhouse) C. Agardh | 1.1 | Thick–leathery | |
| Cystoseira compressa (Esper) Gerloff & Nizamuddin | 2 | Thick–leathery | |
| Cystoseira amentacea C. Agardh Bory de Saint-Vincent var. amantaceaPadina pavonica (Linnaeus) Thivy | 1.1 2 | Thick–leathery Thick–leathery | |
| Sargassum vulgare C. Agardh | 2 | Thick–leathery | |
| Gigartina pistillata (S.G. Gmelin) Stackhouse | 3 | Rhodophyta/Florideophyceae | Coarsely-branched |
| Gigartina teedei (Mertens ex Roth) J.V. Lamouroux | 3 | Coarsely-branched | |
| Gracilaria bursa-pastoris S.G.Gmelin) P.C. Silva Gracilaria gracilis (Stackhouse) C. Steentoft, L.M. Irvine & Farnham Gracilaria sp. | 4 1.1 3 | Thick–leathery Thick–leathery Thick–leathery |
| IC50 (mg/mL) | mg GAE/g dw Extract | |||||
|---|---|---|---|---|---|---|
| DPPH•a | ABTS•+ a | •OH a | O2•− a | TPC c | Extraction Yield b (%) | |
| Macroalgae species | ||||||
| Green macroalgae | ||||||
| Ulva rigida | 5.50 ± 0.20 * | 0.95 ± 0.04 * | 0.70 ± 0.01 * | ND | 4.15 ± 0.16 | 18.0 ± 0.90 |
| Ulva intestinalis | 10.00 ± 0.11 * | 0.98 ± 0.02 * | 0.45 ± 0.03 * | ND | 2.11 ± 0.07 | 24.0 ± 0.96 |
| Codium fragile | 79.00 ± 10.27 * | 15.00 ± 0.60 * | 1.70 ± 0.07 * | 3.30 ± 0.26 * | 0.55 ± 0.04 | 46.7 ± 3.74 |
| Red macroalgae | ||||||
| Gracilaria gracilis | 13.50 ± 1.20 * | 1.45 ± 0.12 * | 0.65 ± 0.02 * | 6.40 ± 0.80 * | 3.01 ± 0.08 | 25.4 ± 0.51 |
| Gracilaria sp. | 10.00 ± 0.80 * | 0.47 ± 0.03 * | 1.10 ± 0.09 * | 0.36 ± 0.03 * | 3.16 ± 0.09 | 25.5 ± 1.53 |
| Gracilaria bursa pastoris | 9.20 ± 0.074 * | 1.60 ± 0.060 * | 1.50 ± 0.040 * | 0.14 ± 0.012 * | 2.00 ± 0.03 | 20.2 ± 1.82 |
| Gigartina teedei | 0.31 ± 0.006 * | 0.02 ± 0.001 * | 0.10 ± 0.007 * | 0.05 ± 0.003 * | 12.53 ± 0.88 | 20.3 ± 1.42 |
| Gigartina pistillata | 2.10 ± 0.126 * | 0.16 ± 0.012 * | 0.14 ± 0.011 * | 0.07 ± 0.001 * | 3.70 ± 0.20 | 18.9 ± 0.95 |
| Brown macroalgae | ||||||
| Colpomenia sinuosa | 15.00 ± 1.80 * | 2.5 ± 0.12 * | 10 ± 0.30 * | 1.10 ± 0.04 * | 0.69 ± 0.02 | 19.7 ± 1.57 |
| Cystoseira barbata | 1.40 ± 0.07 * | 0.43 ± 0.03 * | 2.60 ± 0.18 * | 1.20 ± 0.04 * | 5.76 ± 0.15 | 22.4 ± 2.46 |
| Cystoseira amentacea | 2.5 ± 0.05 * | 0.58 ± 0.05 * | 0.16 ± 0.01 * | 1.40 ± 0.11 * | 2.54 ± 0.14 | 34.1 ± 1.36 |
| Cystoseira compressa | 2.90 ± 0.06 * | 0.75 ± 0.03 * | 1.40 ± 0.11 * | 1.10 ± 0.10 * | 2.93 ± 0.13 | 28.3 ± 2.26 |
| Sargasum vulgare | 8.20 ± 0.82 * | 1.40 ± 0.07 * | 1.30 ± 0.02 * | 0.60 ± 0.05 * | 2.53 ± 0.03 | 27.0 ± 0.81 |
| Padina pavonica | 6.50 ± 0.45 * | 0.38 ± 0.03 * | 0.40 ± 0.01 * | 0.40 ± 0.04 * | 2.77 ± 0.17 | 18.4 ± 1.29 |
| Positive control | ||||||
| Ascorbic acid | 0.005 ± 0.0002 * | 0.004 ± 0.0001 * | 0.218 ± 0.013 * | ND | NT | NT |
| Methods | ABTS•+ | •OH | O2•− | RP | DNA Protection | XTT HepG2 | TPC |
|---|---|---|---|---|---|---|---|
| DPPH. | 0.825 ** | 0.471 | 0.283 | 0.964 ** | 0.685 ** | 0.099 | −0.737 ** |
| ABTS•+ | 0.653 * | 0.361 | 0.789 ** | 0.607 * | 0.077 | −0.789 ** | |
| •OH | 0.403 | 0.451 | 0.198 | 0.494 | −0.442 | ||
| O2•− | 0.289 | 0.340 | 0.457 | −0.284 | |||
| RP | 0.568 * | 0.075 | −0.697 ** | ||||
| DNA protection | −0.136 | −0.768 ** | |||||
| XTT HepG2 | −0.004 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goutzourelas, N.; Kevrekidis, D.P.; Barda, S.; Malea, P.; Trachana, V.; Savvidi, S.; Kevrekidou, A.; Assimopoulou, A.N.; Goutas, A.; Liu, M.; et al. Antioxidant Activity and Inhibition of Liver Cancer Cells’ Growth of Extracts from 14 Marine Macroalgae Species of the Mediterranean Sea. Foods 2023, 12, 1310. https://doi.org/10.3390/foods12061310
Goutzourelas N, Kevrekidis DP, Barda S, Malea P, Trachana V, Savvidi S, Kevrekidou A, Assimopoulou AN, Goutas A, Liu M, et al. Antioxidant Activity and Inhibition of Liver Cancer Cells’ Growth of Extracts from 14 Marine Macroalgae Species of the Mediterranean Sea. Foods. 2023; 12(6):1310. https://doi.org/10.3390/foods12061310
Chicago/Turabian StyleGoutzourelas, Nikolaos, Dimitrios Phaedon Kevrekidis, Sofia Barda, Paraskevi Malea, Varvara Trachana, Stavroula Savvidi, Alkistis Kevrekidou, Andreana N. Assimopoulou, Andreas Goutas, Ming Liu, and et al. 2023. "Antioxidant Activity and Inhibition of Liver Cancer Cells’ Growth of Extracts from 14 Marine Macroalgae Species of the Mediterranean Sea" Foods 12, no. 6: 1310. https://doi.org/10.3390/foods12061310