
High-Efficiency Fermentation of Nattokinase by Recombinant PSP2 Using Oyster Protein Hydrolysate as a Substrate
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain and Culture Conditions
2.2. Chemicals
2.3. Preparation of Oyster Protein Hydrolysate
2.4. Enzymatic Activity Assay
2.5. Best Conditions of Fermentation Conditions
2.6. ACE Inhibitory Activity
2.7. Analysis of Fermentation Liquid Components
2.8. Flavor Analysis of the Fermentation Liquid
2.9. Determination of TVB-N and Heavy Metal Levels
2.10. Statistical Analysis
3. Results and Discussion
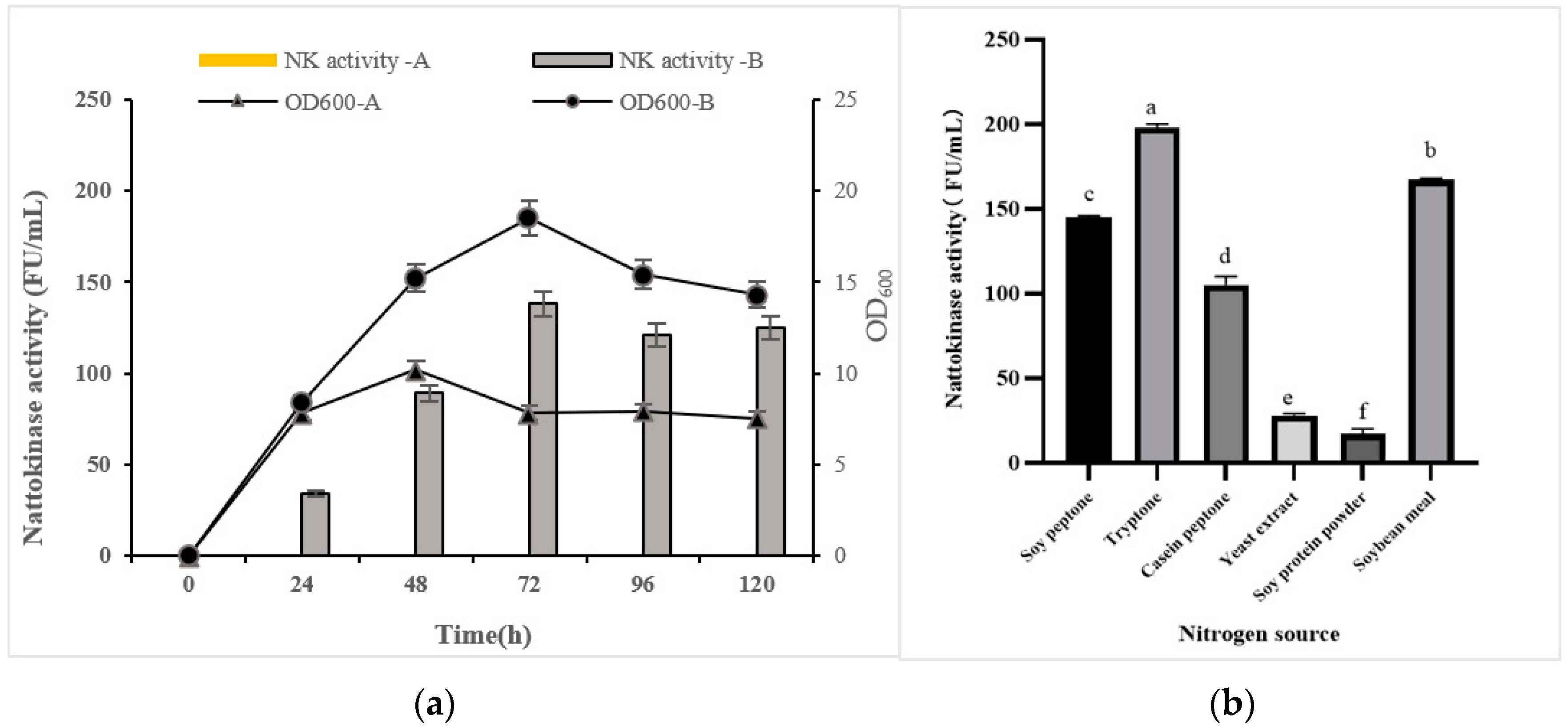
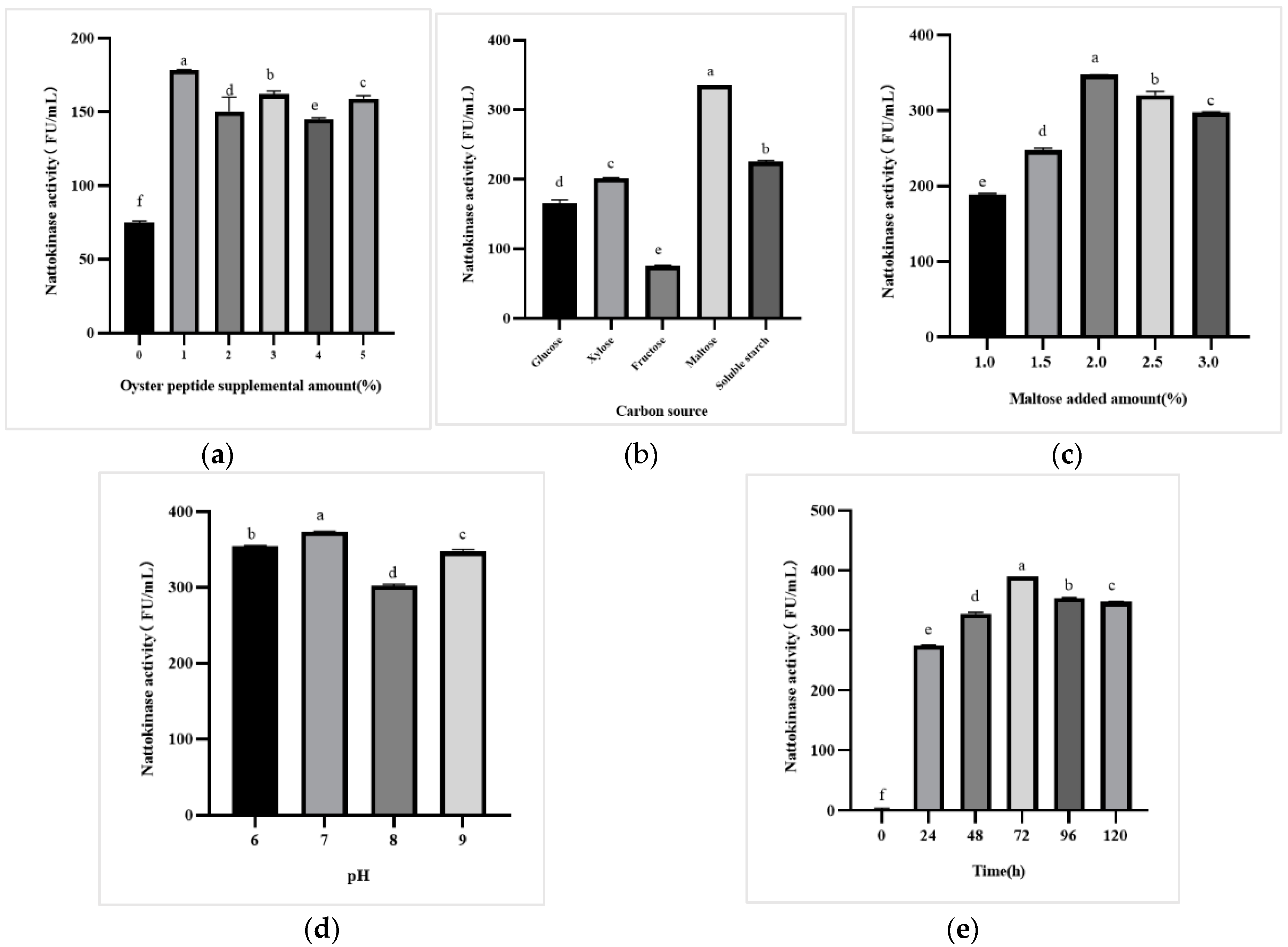
3.1. Best Conditions of Fermentation
3.2. Analysis of the Fermentation Liquid Components
3.3. Flavor Analysis of the Fermentation Liquid
3.4. Determination of TVB-N and Heavy Metal Levels
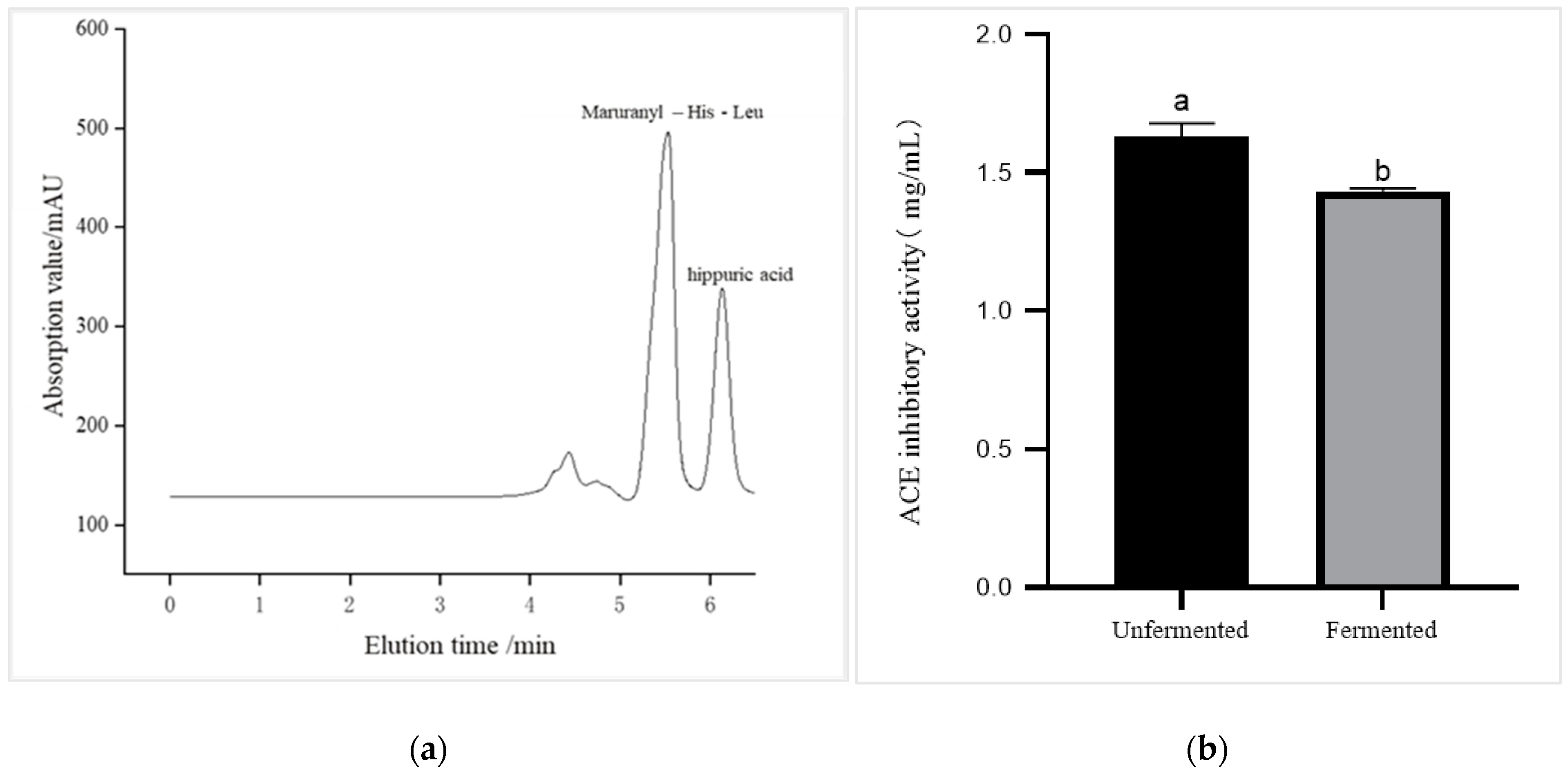
3.5. ACE Inhibitory Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Buckley, L.F.; Carbone, S.; Aldemerdash, A.; Fatani, N.; Fanikos, J. Novel and emerging therapeutics for primary prevention of cardiovascular disease. Am. J. Med. 2019, 132, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Ramalho, S.H.; Shah, A.M. Lung function and cardiovascular disease: A link. Trends Cardiovasc. Med. 2021, 31, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Zhu, C.; Chen, S. Microbial production of nattokinase: Current progress, challenge and prospect. World J. Microbiol. Biotechnol. 2017, 33, 84. [Google Scholar] [CrossRef] [PubMed]
- Hara, T.; Aumayr, A.; Fujio, Y.; Ueda, S. Elimination of plasmid-linked polyglutamate production by Bacillus subtilis (natto) with acridine orange. Appl. Environ. Microbiol. 1982, 44, 1456–1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Li, Y.; Liang, X.; Liu, H.; Xiao, J. Research progress of liquid fermentation of nattokinase. Food Ferment. Ind. 2017, 43, 256–262. [Google Scholar] [CrossRef]
- Hsia, C.-H.; Shen, M.-C.; Lin, J.-S.; Wen, Y.-K.; Hwang, K.-L.; Cham, T.-M.; Yang, N.-C. Nattokinase decreases plasma levels of fibrinogen, factor VII, and factor VIII in human subjects. Nutr. Res. 2009, 29, 190–196. [Google Scholar] [CrossRef]
- Zhang, X.; Lyu, X.; Tong, Y.; Wang, J.; Ye, J.; Yang, R. Chitosan/casein based microparticles with a bilayer shell–core structure for oral delivery of nattokinase. Food Funct. 2020, 11, 10799–10816. [Google Scholar] [CrossRef]
- Bhatt, P.C.; Pathak, S.; Kumar, V.; Panda, B.P. Attenuation of neurobehavioral and neurochemical abnormalities in animal model of cognitive deficits of Alzheimer’s disease by fermented soybean nanonutraceutical. Inflammopharmacology 2018, 26, 105–118. [Google Scholar] [CrossRef]
- Takano, A.; Ogasawara, K.; Inomata, Y.; Hirata, A.; Sagara, N.; Kawaji, T.; Tanihara, H. Posterior Vitreous Detachment Induced by Nattokinase (Subtilisin NAT): A Novel Enzyme for Pharmacologic Vitreolysis. Investig. Opthalmology Vis. Sci. 2006, 47, 2075–2079. [Google Scholar] [CrossRef]
- Chen, Y.; Li, P.; Liao, L.; Qin, Y.; Jiang, L.; Liu, Y. Characteristic fingerprints and volatile flavor compound variations in Liuyang Douchi during fermentation via HS-GC-IMS and HS-SPME-GC-MS. Food Chem. 2021, 361, 130055. [Google Scholar] [CrossRef]
- Ageno, W.; Becattini, C.; Brighton, T.; Selby, R.; Kamphuisen, P.W. Cardiovascular risk factors and venous thromboembolism: A meta-analysis. Circulation 2008, 117, 93–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, A.; Patterson, C.; Yarnell, J.; Rumley, A.; Ben-Shlomo, Y.; Lowe, G. Which Hemostatic Markers Add to the Predictive Value of Conventional Risk Factors for Coronary Heart Disease and Ischemic Stroke? The Caerphilly Study. Circulation 2005, 112, 3080–3087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nani, J.; Yonamine, C.M.; Musial, D.C.; Mas, C.D.; Mari, J.J.; Hayashi, M.A.F. ACE activity in blood and brain axis in an animal model for schizophrenia: Effects of dopaminergic manipulation with antipsychotics and psychostimulants. World J. Biol. Psychiatry 2019, 21, 53–63. [Google Scholar] [CrossRef]
- Zhang, T.; Li, M.; Fu, X.; Mou, H. Purification and charicterization of angiotensin I-converting enzyme (ACE) inhibitory peptides with specific structure X-Pro. Eur. Food Res. Technol. 2019, 245, 1743–1753. [Google Scholar] [CrossRef]
- Wang, J.; Hu, J.; Cui, J.; Bai, X.; Du, Y.; Miyaguchi, Y.; Lin, B. Purification and identification of a ACE inhibitory peptide from oyster proteins hydrolysate and the antihypertensive effect of hydrolysate in spontaneously hypertensive rats. Food Chem. 2008, 111, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Feng, X.; Lan, X.; Xu, Y.; Liao, D. Purification and identification of Angiotensin-I Converting Enzyme (ACE) inhibitory peptide from lizard fish (Saurida elongata) hydrolysate. J. Funct. Foods 2015, 13, 295–299. [Google Scholar] [CrossRef]
- Yao, G.-L.; Chai, Y.; Chen, J.; Wu, Y.-G. Separation and identification of ACE inhibitory peptides from cashew nut (Anacardium occidentale Linnaeus) protein. Int. J. Food Prop. 2017, 20, S981–S991. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Ma, L.; Zhang, Y. Influence of pretreatment on enzymatic hydrolysis of collagen and release of angiotensin converting enzyme (ACE) inhibitory peptides. Food Sci. 2014, 35, 21–25. [Google Scholar] [CrossRef]
- Song, X.; Yang, Y.; Liang, N.; Yang, F.; Chen, S.; Zhou, L.; Zhou, K.; Wang, Y. Quantitative pharmacokinetic evaluation of Subtilisin QK-2 after a bolus IV injection in a rat model using a novel sandwich enzyme-linked immunosorbent assay. J. Pharm. Biomed. Anal. 2020, 186, 113264. [Google Scholar] [CrossRef] [PubMed]
- Minh, N.H.; Trang, H.T.Q.; Van, T.B.; Loc, N.H. Production and purification of nattokinase from Bacillus subtilis. Food Biotechnol. 2022, 36, 1–21. [Google Scholar] [CrossRef]
- Liang, S.; Zhang, T.; Fu, X.; Zhu, C.; Mou, H. Partially degraded chitosan-based flocculation to achieve effective deodorization of oyster (Crassostrea gigas) hydrolysates. Carbohydr. Polym. 2020, 234, 115948. [Google Scholar] [CrossRef] [PubMed]
- Pagnoncelli, M.; Fernandes, M.; Rodrigues, C.; Soccol, C. 22—Nattokinases. In Current Developments in Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2017; pp. 509–526. [Google Scholar] [CrossRef]
- Liu, J.; Xing, J.; Chang, T.; Ma, Z.; Liu, H. Optimization of nutritional conditions for nattokinase production by Bacillus natto NLSSE using statistical experimental methods. Process. Biochem. 2005, 40, 2757–2762. [Google Scholar] [CrossRef]
- Tian, Y. Study on Plasminase Production by Natto Bacteria Liquid Fermentation. Master’s Thesis, South China Agricultural University, Guangzhou, China, 2001. (In Chinese). [Google Scholar]
- Suwanmanon, K.; Hsieh, P.-C. Isolating Bacillus subtilis and optimizing its fermentative medium for GABA and nattokinase production. CyTA-J. Food 2014, 12, 282–290. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Lan, G.; Tian, X.; He, L.; Li, C.; Tao, H.; Zeng, X.; Wang, X. Effect of Adding Bifidobacterium animalis BZ25 on the Flavor, Functional Components and Biogenic Amines of Natto by Bacillus subtilis GUTU09. Foods 2022, 11, 2674. [Google Scholar] [CrossRef] [PubMed]
- Mariutti, L.R.; Bragagnolo, N. Influence of salt on lipid oxidation in meat and seafood products: A review. Food Res. Int. 2017, 94, 90–100. [Google Scholar] [CrossRef]
- Pino, J.A. Odour-active compounds in papaya fruit cv. Red Maradol. Food Chem. 2014, 146, 120–126. [Google Scholar] [CrossRef]
- F2017C00333; Australia New Zealand Food Standards Code—Schedule 19 Maximum Levels of Contaminants and Natural Toxicants. Food Standards Australia New Zealand: Canberra, Australia, 2017.
- Beveridge, T.J.; Murray, R.G. Sites of metal deposition in the cell wall of Bacillus subtilis. J. Bacteriol. 1980, 141, 876–887. [Google Scholar] [CrossRef] [Green Version]
- Hua, X.; Sun, L.; Zhong, C.; Wu, Q.; Xiao, P.; Yoshida, A.; Liu, G.; Cao, M. Successive digestion of tilapia collagen by serine proteinase and proline specific endopeptidase to produce novel angiotensin I-converting enzyme inhibitory peptides. Mar. Life Sci. Technol. 2020, 2, 268–278. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Oyster Fermentation Liquid (0 h) | Oyster Fermentation Liquid (72 h) | |
|---|---|---|
| Solid content (%) | 4.82 ± 0.23 a | 3.81 ± 0.18 b |
| Total sugar (%) | 3.73 ± 0.54 b | 8.92 ± 1.37 a |
| Total protein (%) | 69.29 ± 1.25 b | 78.21 ± 0.74 a |
| Asp (%) | 10.14 ± 0.47 a | 9.86 ± 0.25 a |
| Thr (%) | 5.28 ± 0.23 a | 5.03 ± 0.12 a |
| Ser (%) | 5.03 ± 0.15 a | 4.08 ± 0.22 b |
| Glu (%) | 14.95 ± 0.76 a | 14.24 ± 0.42 a |
| Pro (%) | 4.54 ± 0.47 a | 3.72 ± 0.22 b |
| Gly (%) | 6.53 ± 0.81 a | 6.45 ± 0.13 a |
| Ala (%) | 10.52 ± 1.12 a | 10.42 ± 0.94 a |
| Val (%) | 5.47 ± 0.14 a | 4.63 ± 0.29 b |
| Met (%) | 2.25 ± 0.11 a | 2.09 ± 0.45 a |
| Ile (%) | 4.51 ± 0.12 a | 3.63 ± 0.33 b |
| Leu (%) | 7.47 ± 0.47 a | 7.03 ± 0.65 a |
| Tyr (%) | 3.88 ± 0.05 a | 3.08 ± 0.03 b |
| Phe (%) | 4.46 ± 0.12 a | 3.66 ± 0.22 b |
| Lys (%) | 10.20 ± 0.98 a | 10.14 ± 0.74 a |
| His (%) | 1.52 ± 0.03 a | 1.48 ± 0.05 a |
| Arg (%) | 3.05 ± 0.08 a | 2.99 ± 0.14 a |
| Samples | Molecular Weight Distribution | |
|---|---|---|
| Oyster fermentation liquid (0 h) | Molecular weight range (kDa) | Proportion (%) |
| 3~50 | 94.72 ± 1.25 | |
| 50~150 | 3.53 ± 0.54 | |
| 150~14,800 | 1.75 ± 0.70 | |
| Oyster fermentation liquid (72 h) | 60~85 | 42.08 ± 1.56 |
| 85~310 | 54.30 ± 1.18 | |
| More than 310 | 3.62 ± 0.35 | |
| Volatile Substance | Smell | Oyster Protein Hydrolysate (Unfermented) | Oyster Protein Hydrolysate (Fermented) |
|---|---|---|---|
| Isovaleraldehyde | Stench | 4.07 ± 0.48 | |
| Hexanal | Clams taste | 3.15 ± 0.63 | |
| Heptylaldehyde | Clams taste stench | 5.10 ± 1.14 | |
| Nonylaldehyde | Clams taste | 2.83 ± 0.53 | |
| Benzaldehyde | Contribute to pleasant smells | 9.21 ± 1.25 | 14.16 ± 1.18 |
| 2-Furyl alcohol | Contribute to pleasant smells | 7.48 ± 0.68 | |
| 3,4-Dimethylbenzaldehyde | Contribute to pleasant smells | 2.54 ± 0.82 | |
| 1,2, 3-Trimethylbenzene | Contribute to pleasant smells | 0.94 ± 0.07 | |
| 1,3, 5-Trimethylbenzene | Contribute to pleasant smells | 1.70 ± 0.20 | |
| Silane | Contribute to pleasant smells | 7.36 ± 0.94 | |
| Furfurylalcohol | Contribute to pleasant smells | 2.51 ± 0.73 | 5.12 ± 0.04 |
| Cyanobenzene | Contribute to pleasant smells | 10.40 ± 0.91 | |
| O-methyl benzonitrile | Contribute to pleasant smells | 29.02 ± 1.44 | |
| Methoxy-phenyl-oxime | Stench | 6.51 ± 1.26 | |
| 2, 4-Di-tert-butylphenol | Anxious burnt smell | 5.73 ± 0.97 | |
| Dimethyl disulfide | Contribute to pleasant smells | 2.01 ± 0.45 | |
| 2-Heptyl ketone | Contribute to pleasant smells | 7.18 ± 0.38 | |
| 6-Methyl-2-heptanone | Contribute to pleasant smells | 2.57 ± 0.18 | |
| 2-Methylpyrazine | Contribute to pleasant smells | 0.31 ± 0.22 | |
| 2, 5-Dimethylpyrazine | Contribute to pleasant smells | 35.31 ± 1.44 | |
| 2-Ethyl-5-methylpyrazine | Contribute to pleasant smells | 2.07 ± 1.01 | |
| 2,3, 5-Trimethylpyrazine | Contribute to pleasant smells | 1.16 ± 0.08 | |
| 2-Decyl ketone | Contribute to pleasant smells | 0.68 ± 0.36 | |
| 2-Ethylhexanol | Contribute to pleasant smells | 0.94 ± 0.53 | |
| Benzene ethanol | Contribute to pleasant smells | 6.79 ± 1.12 | |
| Phenylacetonitrile | Contribute to pleasant smells | 5.90 ± 0.90 |
| Contents | Oyster Protein Hydrolysate (Unfermented) | Oyster Protein Hydrolysate (Fermented) |
|---|---|---|
| TVB-N (mg/100 g) | 7.18 ± 0.12 a | 6.84 ± 0.04 b |
| Total arsenic (mg/kg) | 0.100 ± 0.003 a | 0.087 ± 0.012 a |
| Total mercury (mg/kg) | <0.003 | <0.003 |
| Lead (mg/kg) | <0.020 | <0.020 |
| Cadmium (mg/kg) | 0.056 ± 0.005 a | 0.039 ± 0.001 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, M.; Ning, C.; Peng, S.; Li, D.; Jin, R.; Zhang, Y.; Liu, Z.; Mou, H.; Zhu, C. High-Efficiency Fermentation of Nattokinase by Recombinant PSP2 Using Oyster Protein Hydrolysate as a Substrate. Foods 2023, 12, 1252. https://doi.org/10.3390/foods12061252
Tian M, Ning C, Peng S, Li D, Jin R, Zhang Y, Liu Z, Mou H, Zhu C. High-Efficiency Fermentation of Nattokinase by Recombinant PSP2 Using Oyster Protein Hydrolysate as a Substrate. Foods. 2023; 12(6):1252. https://doi.org/10.3390/foods12061252
Chicago/Turabian StyleTian, Ming, Chen Ning, Siyuan Peng, Deyu Li, Renyi Jin, Yang Zhang, Zhemin Liu, Haijin Mou, and Changliang Zhu. 2023. "High-Efficiency Fermentation of Nattokinase by Recombinant PSP2 Using Oyster Protein Hydrolysate as a Substrate" Foods 12, no. 6: 1252. https://doi.org/10.3390/foods12061252