
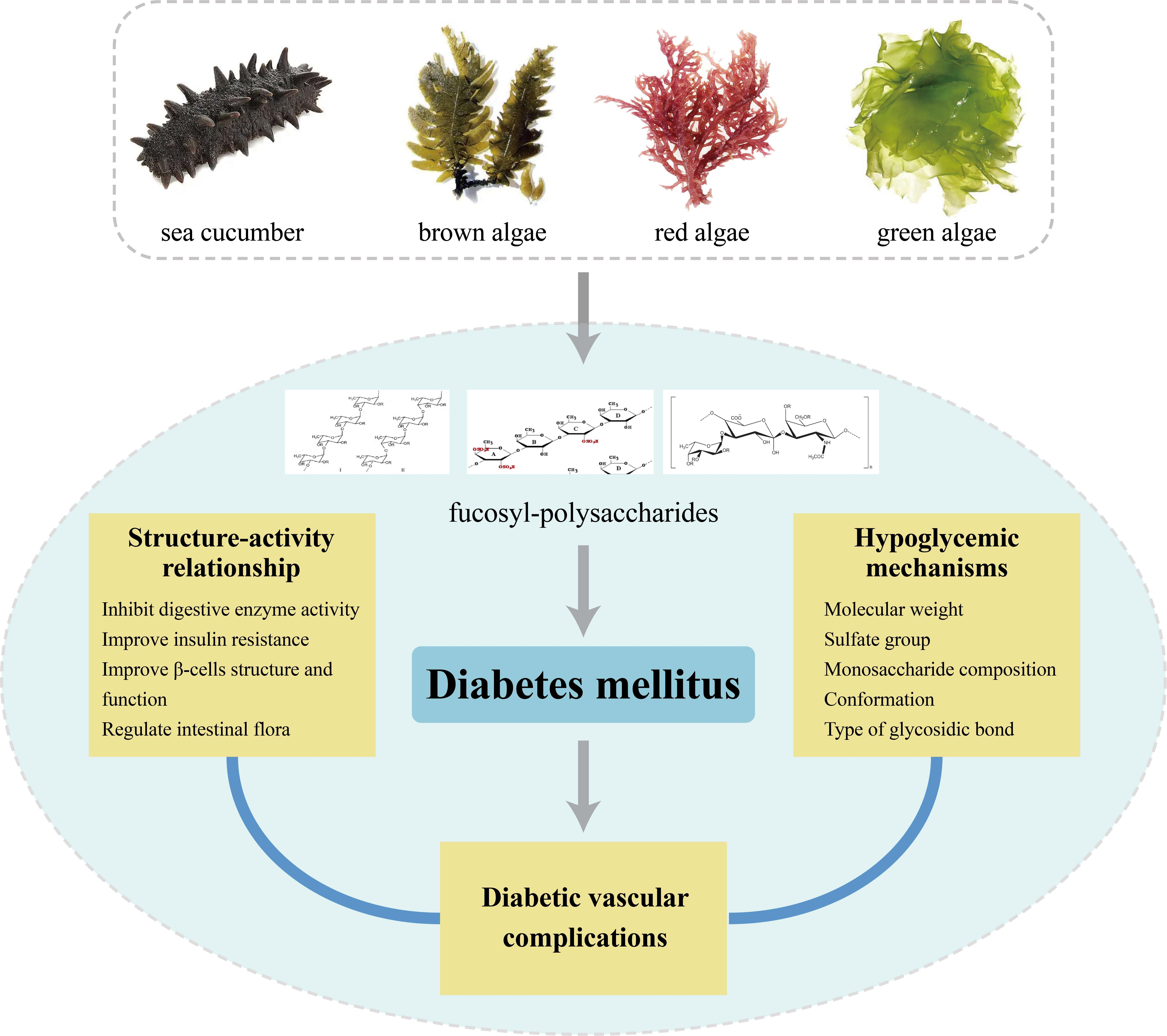
Potential Application of Marine Fucosyl-Polysaccharides in Regulating Blood Glucose and Hyperglycemic Complications
Abstract
:
1. Introduction
2. Marine Polysaccharides with Hypoglycemic Activity
3. Hypoglycemic Mechanisms of Marine Fucosyl-Polysaccharides
3.1. Inhibit Digestive Enzyme Activity
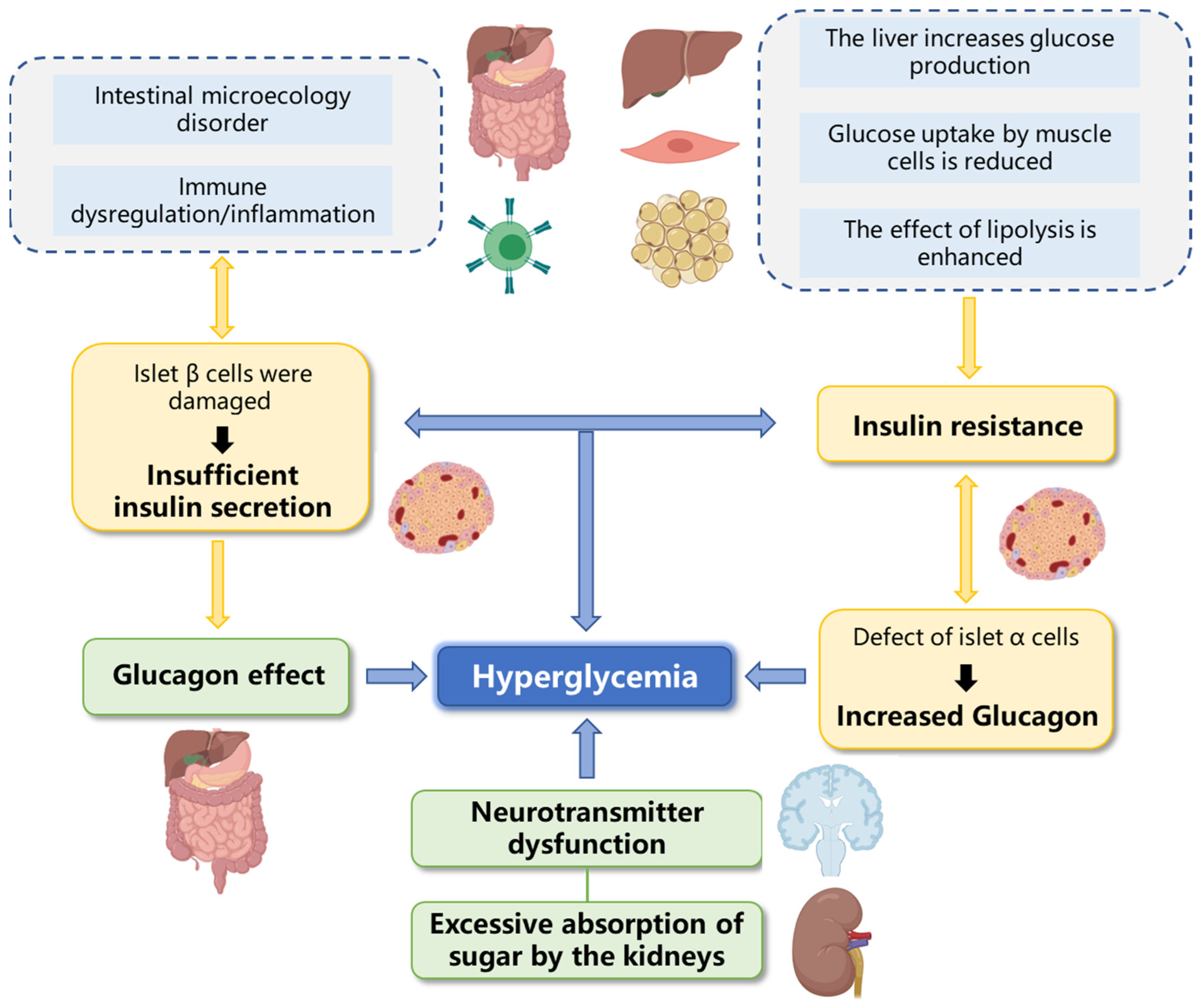
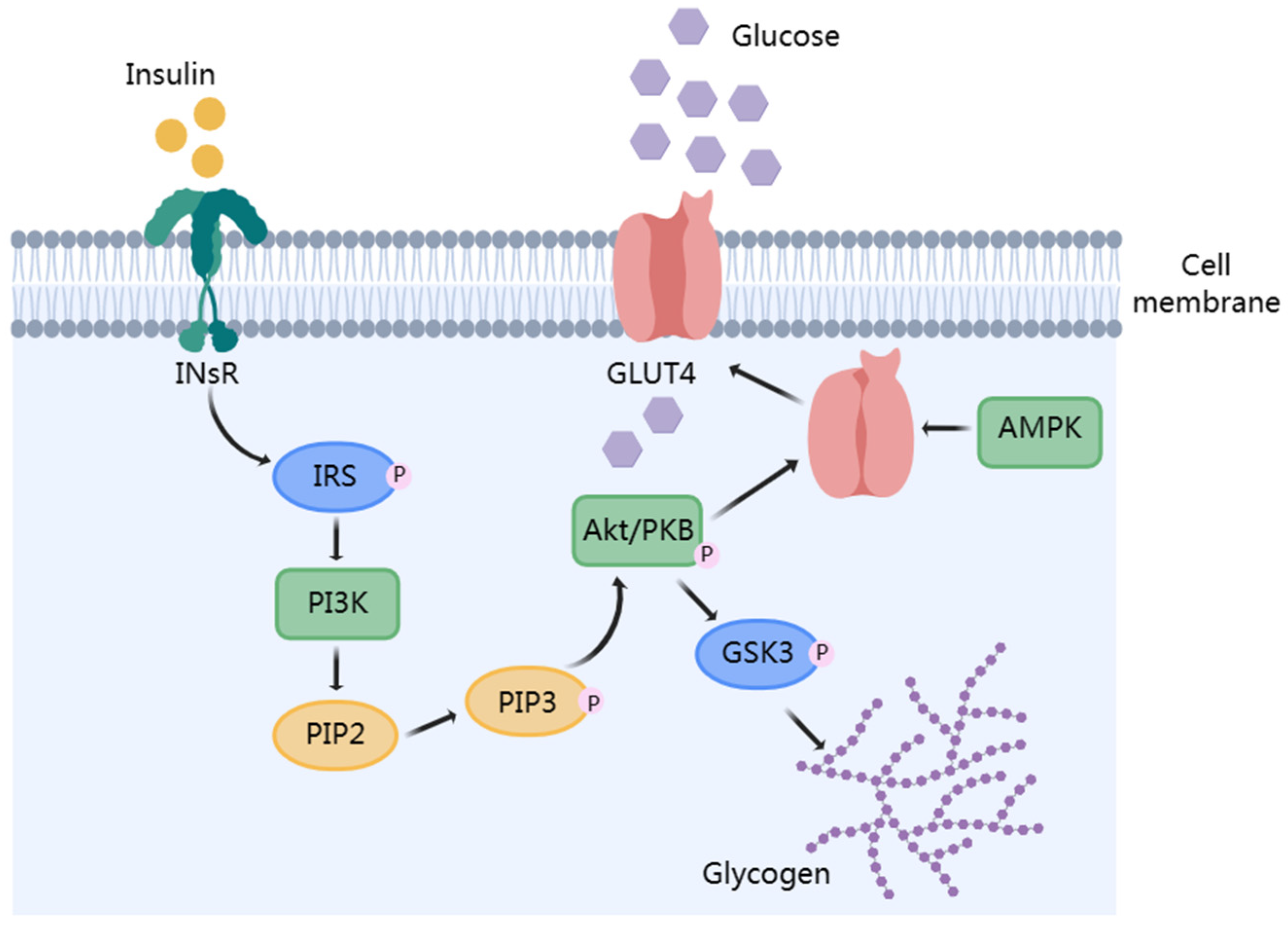
3.2. Improve Insulin Resistance
3.3. Improve β-Cells Structure and Function
3.4. Regulate Intestinal Microflora
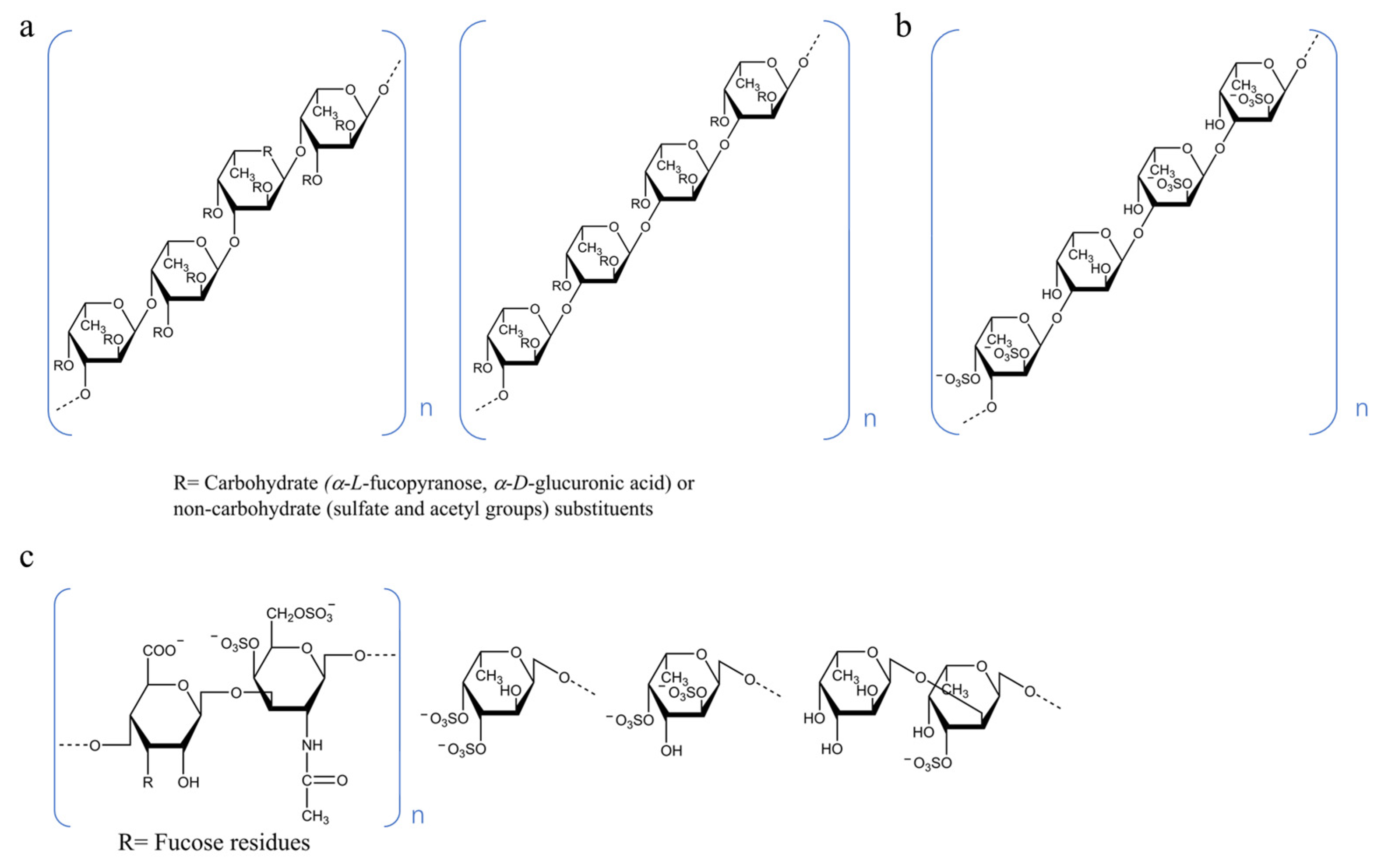
4. Structure–Activity Relationship (SAR)
4.1. Molecular Weight
4.2. Sulfate Group
4.2.1. Content of Sulfate Group
4.2.2. Substitution Position of the Sulfate Group
4.3. Monosaccharide Composition
4.4. Conformation
4.5. Type of Glycosidic Bond
5. Effect of Marine Fucosyl-Polysaccharides on Diabetic Vascular Complications
6. Discussion and Prospects
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kovatchev, B.P.; Cox, D.J.; Gonder-Frederick, L.A.; Clarke, W. Symmetrization of the blood Glucose measurement scale and its applications. Diabetes Care 1997, 20, 1655–1658. [Google Scholar] [CrossRef] [PubMed]
- American Diabetes Association. 15. Diabetes care in the hospital: Standards of medical care in diabetes—2020. Diabetes Care 2020, 43, S193–S202. [Google Scholar] [CrossRef] [Green Version]
- Marik, P.E.; Bellomo, R. Stress hyperglycemia: An essential survival response! Crit. Care 2013, 17, 305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganesan, K.; Xu, B.; Froldi, G. Anti-diabetic effects and mechanisms of dietary polysaccharides. Molecules 2019, 24, 2556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Punthakee, Z.; Goldenberg, R.; Katz, P.; Diabetes, C.C.P.G. Definition, classification and diagnosis of diabetes, prediabetes and metabolic syndrome. Can. J. Diabetes 2018, 42, S10–S15. [Google Scholar] [CrossRef] [Green Version]
- Chiang, J.L.; Maahs, D.M.; Garvey, K.C.; Hood, K.K.; Laffel, L.M.; Weinzimer, S.A.; Wolfsdorf, J.I.; Schatz, D. Type 1 diabetes in children and adolescents: A position statement by the American diabetes association. Diabetes Care 2018, 41, 2026–2044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padhi, S.; Nayak, A.K.; Behera, A. Type II diabetes mellitus: A review on recent drug based therapeutics. Biomed. Pharmacother. 2020, 131, 110708. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.; Wang, D.; Coresh, J.; Selvin, E. Trends in diabetes treatment and control in U.S. adults, 1999–2018. N. Engl. J. Med. 2021, 384, 2219–2228. [Google Scholar] [CrossRef] [PubMed]
- Kharroubi, A.T.; Darwish, H.M. Diabetes mellitus: The epidemic of the century. World J. Diabetes 2015, 6, 850–867. [Google Scholar] [CrossRef]
- Al-Ishaq, R.K.; Abotaleb, M.; Kubatka, P.; Kajo, K.; Büsselberg, D. Flavonoids and their anti-diabetic effects: Cellular mechanisms and effects to improve blood sugar levels. Biomolecules 2019, 9, 430. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Xiong, R.; Wang, L.; Cui, J.; Shi, L.; Liu, Y.; Luo, B. Associations of dietary habits, physical activity and cognitive views with gestational diabetes mellitus among Chinese women. Public Health Nutr. 2014, 17, 1850–1857. [Google Scholar] [CrossRef] [Green Version]
- American, D.A. Diagnosis and classification of diabetes mellitus. Diabetes Care 2014, 37 (Suppl. S1), S81–S90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- American, D.A. Diagnosis and classification of diabetes mellitus. Diabetes Care 2008, 31 (Suppl. S1), S55–S60. [Google Scholar] [CrossRef] [Green Version]
- Chiefari, E.; Arcidiacono, B.; Foti, D.; Brunetti, A. Gestational diabetes mellitus: An updated overview. J. Endocrinol. Investig. 2017, 40, 899–909. [Google Scholar] [CrossRef]
- Han, H.S.; Kang, G.; Kim, J.S.; Choi, B.H.; Koo, S.H. Regulation of glucose metabolism from a liver-centric perspective. Exp. Mol. Med. 2016, 48, e218. [Google Scholar] [CrossRef] [Green Version]
- Yaribeygi, H.; Farrokhi, F.R.; Butler, A.E.; Sahebkar, A. Insulin resistance: Review of the underlying molecular mechanisms. J. Cell. Physiol. 2019, 234, 8152–8161. [Google Scholar] [CrossRef]
- Roden, M.; Shulman, G.I. The integrative biology of type 2 diabetes. Nature 2019, 576, 51–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ansari, P.; Akther, S.; Khan, J.T.; Islam, S.S.; Masud, M.S.R.; Rahman, A.; Seidel, V.; Abdel-Wahab, Y.H.A. Hyperglycaemia-linked diabetic foot complications and their management using conventional and alternative therapies. Appl. Sci. 2022, 12, 11777. [Google Scholar] [CrossRef]
- Stehouwer, C.D.A. Microvascular dysfunction and hyperglycemia: A vicious cycle with widespread consequences. Diabetes 2018, 67, 1729–1741. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Saeedi, P.; Karuranga, S.; Pinkepank, M.; Ogurtsova, K.; Duncan, B.B.; Stein, C.; Basit, A.; Chan, J.C.N.; Mbanya, J.C.; et al. IDF diabetes atlas: Global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes Res. Clin. Pract. 2022, 183, 109119. [Google Scholar] [CrossRef]
- Chen, H.; Nie, Q.; Hu, J.; Huang, X.; Zhang, K.; Pan, S.; Nie, S. Hypoglycemic and hypolipidemic effects of glucomannan extracted from konjac on type 2 diabetic rats. J. Agr. Food Chem. 2019, 67, 5278–5288. [Google Scholar] [CrossRef] [PubMed]
- Philippe, J.; Raccah, D. Treating type 2 diabetes: How safe are current therapeutic agents? Int. J. Clin. Pract. 2009, 63, 321–332. [Google Scholar] [CrossRef]
- Kerru, N.; Singh-Pillay, A.; Awolade, P.; Singh, P. Current anti-diabetic agents and their molecular targets: A review. Eur. J. Med. Chem. 2018, 152, 436–488. [Google Scholar] [CrossRef]
- Setter, S.M.; Iltz, J.L.; Thams, J.; Campbell, R.K. Metformin hydrochloride in the treatment of type 2 diabetes mellitus: A clinical review with a focus on dual therapy. Clin. Ther. 2003, 25, 2991–3026. [Google Scholar] [CrossRef]
- American Diabetes Association. 9. Pharmacologic approaches to glycemic treatment: Standards of medical care in diabetes—2020. Diabetes Care 2020, 43, S98–S110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petite, S.E. Noninsulin medication therapy for hospitalized patients with diabetes mellitus. Am. J. Health Syst. Pharm. 2018, 75, 1361–1368. [Google Scholar] [CrossRef]
- Hwang, J.; Yadav, D.; Lee, P.C.; Jin, J.O. Immunomodulatory effects of polysaccharides from marine algae for treating cancer, infectious disease, and inflammation. Phytother. Res. 2022, 36, 761–777. [Google Scholar] [CrossRef]
- Cheng, J.; Xiao, M.; Ren, X.; Secundo, F.; Yu, Y.; Nan, S.; Chen, W.; Zhu, C.; Kong, Q.; Huang, Y.; et al. Response of Salmonella enterica serovar Typhimurium to alginate oligosaccharides fermented with fecal inoculum: Integrated transcriptomic and metabolomic analyses. Mar. Life Sci. Technol. 2023, 5, 242–256. [Google Scholar] [CrossRef]
- Anisha, G.S.; Padmakumari, S.; Patel, A.K.; Pandey, A.; Singhania, R.R. Fucoidan from marine macroalgae: Biological actions and applications in regenerative medicine, drug delivery systems and food industry. Bioengineering 2022, 9, 472. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhang, X.; Tang, Y.; Mao, J. Composition, isolation, purification and biological activities of Sargassum fusiforme polysaccharides: A review. Carbohydr. Polym. 2020, 228, 115381. [Google Scholar] [CrossRef]
- Wang, X.; Ye, H.; Cui, J.; Chi, Y.; Liu, R.; Wang, P. Hypolipidemic effect of chromium-modified enzymatic product of sulfated rhamnose polysaccharide from Enteromorpha prolifera in type 2 diabetic mice. Mar. Life Sci. Technol. 2022, 4, 245–254. [Google Scholar] [CrossRef]
- Das, A.; Chen, C.; Mu, S.; Yang, S.; Ju, Y.; Li, S. Medicinal components in edible mushrooms on diabetes mellitus treatment. Pharmaceutics 2022, 14, 436. [Google Scholar] [CrossRef]
- Jiang, X.; Meng, W.; Li, L.; Meng, Z.; Wang, D. Adjuvant therapy with mushroom polysaccharides for diabetic complications. Front. Pharmacol. 2020, 11, 168. [Google Scholar] [CrossRef]
- Khursheed, R.; Singh, S.K.; Wadhwa, S.; Gulati, M.; Awasthi, A. Therapeutic potential of mushrooms in diabetes mellitus: Role of polysaccharides. Int. J. Biol. Macromol. 2020, 164, 1194–1205. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Jia, Y.; Guo, Q. Chapter 6-Polysaccharides and polysaccharide complexes as potential sources of antidiabetic compounds: A review. Stud. Nat. Prod. Chem. 2020, 67, 199–220. [Google Scholar]
- Govindan, S.; Shanmugam, J.; Rajendran, G.; Ramani, P.; Unni, D.; Venkatachalam, B.; Janardhanan, A.; Aswini, K.; Rajendran, R.L.; Gangadaran, P.; et al. Antidiabetic activity of polysaccharide from Hypsizygus ulmarius in streptozotocin-nicotinamide induced diabetic rats. Bioact. Carbohydr. Diet. Fibre 2023, 29, 100350. [Google Scholar] [CrossRef]
- Wang, Y.; Xing, M.; Cao, Q.; Ji, A.; Liang, H.; Song, S. Biological activities of fucoidan and the factors mediating its therapeutic effects: A review of recent studies. Mar. Drugs 2019, 17, 183. [Google Scholar] [CrossRef] [Green Version]
- Jeong, Y.; Kim, Y.D.; Jung, Y.; Park, D.; Lee, D.; Ku, S.; Li, X.; Lu, Y.; Chao, G.H.; Kim, K.; et al. Low molecular weight fucoidan improves endoplasmic reticulum stress-reduced insulin sensitivity through AMP-activated protein kinase activation in L6 myotubes and restores lipid homeostasis in a mouse model of type 2 diabetes. Mol. Pharmacol. 2013, 84, 147–157. [Google Scholar] [CrossRef]
- Kim, K.; Rioux, L.; Turgeon, S.L. Molecular weight and sulfate content modulate the inhibition of α-amylase by fucoidan relevant for type 2 diabetes management. Pharmanutrition 2015, 3, 108–114. [Google Scholar] [CrossRef]
- Ren, B.; Chen, C.; Li, C.; Fu, X.; You, L.; Liu, R.H. Optimization of microwave-assisted extraction of Sargassum thunbergii polysaccharides and its antioxidant and hypoglycemic activities. Carbohydr. Polym. 2017, 173, 192–201. [Google Scholar] [CrossRef]
- Makkar, F.; Chakraborty, K. Antidiabetic and anti-inflammatory potential of sulphated polygalactans from red seaweeds Kappaphycus alvarezii and Gracilaria opuntia. Int. J. Food Prop. 2017, 20, 1326–1337. [Google Scholar] [CrossRef] [Green Version]
- Wen, L.; Zhang, Y.; Sun-Waterhouse, D.; You, L.; Fu, X. Advantages of the polysaccharides from Gracilaria lemaneiformis over metformin in antidiabetic effects on streptozotocin-induced diabetic mice. RSC Adv. 2017, 7, 9141–9151. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.; Luo, X.; Wang, M.; Wang, Z.; Guo, J.; Kong, F.; Bi, Y. Synthesis, characterization, in vitro antioxidant and hypoglycemic activities of selenium nanoparticles decorated with polysaccharides of Gracilaria emaneiformis. Int. J. Biol. Macromol. 2021, 193, 923–932. [Google Scholar] [CrossRef]
- Cao, C.; Huang, Q.; Zhang, B.; Li, C.; Fu, X. Physicochemical characterization and in vitro hypoglycemic activities of polysaccharides from Sargassum pallidum by microwave-assisted aqueous two-phase extraction. Int. J. Biol. Macromol. 2018, 109, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Huang, Q.; Zhang, B.; Li, C.; Liu, R.H.; Cao, C. Structure and in vitro hypoglycemic activity of a homogenous polysaccharide purified from Sargassum pallidum. Food Funct. 2019, 10, 2828–2838. [Google Scholar] [CrossRef]
- Xiao, H.; Fu, X.; Cao, L.; Li, C.; Chen, C.; Huang, Q. Sulfated modification, characterization, antioxidant and hypoglycemic activities of polysaccharides from Sargassum pallidum. Int. J. Biol. Macromol. 2019, 121, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Li, C.; Chen, Q.; Huang, Q.; Pérez, M.E.M.; Fu, X. Physicochemical characterization, potential antioxidant and hypoglycemic activity of polysaccharide from Sargassum pallidum. Int. J. Biol. Macromol. 2019, 139, 1009–1017. [Google Scholar] [CrossRef] [PubMed]
- Heng, X.; Chun, C.; Chao, L.; Qiang, H.; Xiong, F. Physicochemical characterization, antioxidant and hypoglycemic activities of selenized polysaccharides from Sargassum pallidum. Int. J. Biol. Macromol. 2019, 132, 308–315. [Google Scholar] [CrossRef]
- Yang, C.; Lai, S.; Chen, Y.; Liu, D.; Liu, B.; Ai, C.; Wan, X.; Gao, L.; Chen, X.; Zhao, C. Anti-diabetic effect of oligosaccharides from seaweed Sargassum confusum via JNK-IRS1/PI3K signalling pathways and regulation of gut microbiota. Food Chem. Toxicol. 2019, 131, 110562. [Google Scholar] [CrossRef]
- Jia, R.; Li, Z.; Wu, J.; Ou, Z.; Zhu, Q.; Sun, B.; Lin, L.; Zhao, M. Physicochemical properties of polysaccharide fractions from Sargassum fusiforme and their hypoglycemic and hypolipidemic activities in type 2 diabetic rats. Int. J. Biol. Macromol. 2020, 147, 428–438. [Google Scholar] [CrossRef]
- Jia, R.B.; Wu, J.; Luo, D.; Lin, L.; Chen, C.; Xiao, C.; Zhao, M. The beneficial effects of two polysaccharide fractions from Sargassum fusiform against diabetes mellitus ccompanied by dyslipidemia in rats and their underlying mechanisms. Foods 2022, 11, 1416. [Google Scholar] [CrossRef]
- Zhang, Y.; Zuo, J.; Yan, L.; Cheng, Y.; Li, Q.; Wu, S.; Chen, L.; Thring, R.W.; Yang, Y.; Gao, Y.; et al. Sargassum fusiforme fucoidan alleviates high-fat diet-induced obesity and insulin resistance associated with the improvement of hepatic oxidative stress and gut microbiota profile. J. Agric. Food Chem. 2020, 68, 10626–10638. [Google Scholar] [CrossRef]
- Daub, C.D.; Mabate, B.; Malgas, S.; Pletschke, B.I. Fucoidan from Ecklonia maxima is a powerful inhibitor of the diabetes-related enzyme, alpha-glucosidase. Int. J. Biol. Macromol. 2020, 151, 412–420. [Google Scholar] [CrossRef]
- Zeng, A.; Yang, R.; Yu, S.; Zhao, W. A novel hypoglycemic agent: Polysaccharides from laver (Porphyra spp.). Food Funct. 2020, 11, 9048–9056. [Google Scholar] [CrossRef]
- Setyaningsih, I.; Prasetyo, H.; Agungpriyono, D.R.; Tarman, K. Antihyperglycemic activity of Porphyridium cruentum biomass and extra-cellular polysaccharide in streptozotocin-induced diabetic rats. Int. J. Biol. Macromol. 2020, 156, 1381–1386. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Q.; Zhou, T.; Qiu, W.; Wang, Y.; Xu, Q.; Ke, S.; Wang, S.; Jin, W.; Chen, J.; Zhang, H.; et al. Characterization and hypoglycemic effects of sulfated polysaccharides derived from brown seaweed Undaria pinnatifida. Food Chem. 2021, 341, 128148. [Google Scholar] [CrossRef]
- Jia, R.B.; Wu, J.; Li, Z.R.; Ou, Z.R.; Lin, L.; Sun, B.; Zhao, M. Structural characterization of polysaccharides from three seaweed species and their hypoglycemic and hypolipidemic activities in type 2 diabetic rats. Int. J. Biol. Macromol. 2020, 155, 1040–1049. [Google Scholar] [CrossRef]
- Jia, R.B.; Li, Z.R.; Lin, L.; Luo, D.; Chen, C.; Zhao, M. The potential mechanisms of Macrocystis pyrifera polysaccharides mitigating type 2 diabetes in rats. Food Funct. 2022, 13, 7918–7929. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Wu, S.; Cheng, Y.; Zhang, Z.; Mao, G.; Li, S.; Yang, Y.; Zhang, X.; Wu, M.; Tong, H. Sargassum fusiforme fucoidan modifies gut microbiota and intestinal metabolites during alleviation of hyperglycemia in type 2 diabetic mice. Food Funct. 2021, 12, 3572–3585. [Google Scholar] [CrossRef]
- Li, Z.R.; Jia, R.B.; Luo, D.; Lin, L.; Zheng, Q.; Zhao, M. The positive effects and underlying mechanisms of Undaria pinnatifida polysaccharides on type 2 diabetes mellitus in rats. Food Funct. 2021, 12, 11898–11912. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Meng, Q.; Jiang, B.; Chen, J.; Zhang, T.; Zhou, L. Structure characterization and in vitro hypoglycemic effect of partially degraded alginate. Food Chem. 2021, 356, 129728. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Jia, R.; Ou, Z.; Li, Z.; Zhao, M.; Luo, D.; Lin, L. Comparative study on the structural characterization and α-glucosidase inhibitory activity of polysaccharide fractions extracted from Sargassum fusiforme at different pH conditions. Int. J. Biol. Macromol. 2022, 194, 602–610. [Google Scholar] [CrossRef]
- Siddiqui, N.Z.; Rehman, A.U.; Yousuf, W.; Khan, A.I.; Farooqui, N.A.; Zang, S.; Xin, Y.; Wang, L. Effect of crude polysaccharide from seaweed, Dictyopteris divaricata (CDDP) on gut microbiota restoration and anti-diabetic activity in streptozotocin (STZ)-induced T1DM mice. Gut Pathog. 2022, 14, 39. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Ouyang, Y.; Chen, X.; Chen, R.; Ruan, Q.; Farag, M.A.; Chen, X.; Zhao, C. Hypoglycaemic and anti-ageing activities of green alga Ulva lactuca polysaccharide via gut microbiota in ageing-associated diabetic mice. Int. J. Biol. Macromol. 2022, 212, 97–110. [Google Scholar] [CrossRef]
- Zhan, H.; Yu, G.; Zheng, M.; Zhu, Y.; Ni, H.; Oda, T.; Jiang, Z. Inhibitory effects of a low-molecular-weight sulfated fucose-containing saccharide on alpha-amylase and alpha-glucosidase prepared from ascophyllan. Food Funct. 2022, 13, 1119–1132. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Wu, N.; Wang, J.; Geng, L.; Yue, Y.; Wang, F.; Zhang, Q. Low molecular weight fucoidan fraction LF2 improves metabolic syndrome via up-regulating PI3K-AKT-mTOR axis and increasing the abundance of Akkermansia muciniphila in the gut microbiota. Int. J. Biol. Macromol. 2021, 193, 789–798. [Google Scholar] [CrossRef]
- Liu, J.; Zhu, X.; Sun, L.; Gao, Y. Characterization and anti-diabetic evaluation of sulfated polysaccharide from Spirulina platensis. J Funct. Foods 2022, 95, 105155. [Google Scholar] [CrossRef]
- Koh, H.S.A.; Lu, J.; Zhou, W. Structural dependence of sulfated polysaccharide for diabetes management: Fucoidan from Undaria pinnatifida inhibiting α-glucosidase more strongly than α-amylase and amyloglucosidase. Front. Pharmacol. 2020, 11, 831. [Google Scholar] [CrossRef] [PubMed]
- Shan, X.; Liu, X.; Hao, J.; Cai, C.; Fan, F.; Dun, Y.; Zhao, X.; Liu, X.; Li, C.; Yu, G. In vitro and in vivo hypoglycemic effects of brown algal fucoidans. Int. J. Biol. Macromol. 2016, 82, 249–255. [Google Scholar] [CrossRef]
- Hu, S.; Xia, G.; Wang, J.; Wang, Y.; Li, Z.; Xue, C. Fucoidan from sea cucumber protects against high-fat high-sucrose diet-induced hyperglycaemia and insulin resistance in mice. J. Funct. Foods 2014, 10, 128–138. [Google Scholar] [CrossRef]
- Hu, S.; Wang, J.; Wang, J.; Yang, H.; Yan, X.; Su, L. Fucoidan from Acaudina molpadioides improves insulin resistance by altering. J. Funct. Foods 2019, 57, 59–67. [Google Scholar] [CrossRef]
- Hu, S.; Chang, Y.; He, M.; Wang, J.; Wang, Y.; Xue, C. Fucosylated chondroitin sulfate from sea cucumber improves insulin sensitivity via activation of PI3K/PKB pathway. J. Food Sci. 2014, 79, H1424–H1429. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Wang, J.; Xu, H.; Wang, Y.; Li, Z.; Xue, C. Fucosylated chondroitin sulphate from sea cucumber inhibits high-fat-sucrose diet-induced apoptosis in mouse pancreatic islets via down-regulating mitochondrial signaling pathway. J. Funct. Foods 2014, 7, 517–526. [Google Scholar] [CrossRef]
- Li, S.; Jiang, W.; Hu, S.; Song, W.; Ji, L.; Wang, Y.; Cai, L. Fucosylated chondroitin sulphate from Cusumaria frondosa mitigates hepatic endoplasmic reticulum stress and inflammation in insulin resistant mice. Food Funct. 2015, 6, 1547–1556. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Tian, Y.; Chang, Y.; Li, Z.; Xue, C.; Wang, Y. Fucosylated chondroitin sulfate from sea cucumber improves glucose metabolism and activates insulin signaling in the liver of insulin-resistant mice. J. Med. Food 2014, 17, 749–757. [Google Scholar] [CrossRef]
- Wang, J.; Hu, S.; Jiang, W.; Song, W.; Cai, L.; Wang, J. Fucoidan from sea cucumber may improve hepatic inflammatory response and insulin resistance in mice. Int. Immunopharmacol. 2016, 31, 15–23. [Google Scholar] [CrossRef]
- Li, S.; Li, J.; Mao, G.; Wu, T.; Hu, Y.; Ye, X.; Tian, D.; Linhardt, R.J.; Chen, S. A fucoidan from sea cucumber Pearsonothuria graeffei with well-repeated structure alleviates gut microbiota dysbiosis and metabolic syndromes in HFD-fed mice. Food Funct. 2018, 9, 5371–5380. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, H.; Wang, Y.; Shuang, Y.; Mingming, Y.; Tingfu, J.; Zhihua, L. Glycosaminoglycan from Apostichopus japonicus inhibits hepatic glucose production via activating Akt/FoxO1 and inhibiting PKA/CREB signaling pathways in insulin resistant hepatocytes. Food Funct. 2019, 10, 7565–7575. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, J.; Mao, G.; Wu, T.; Lin, D.; Hu, Y.; Ye, X.; Tian, D.; Chai, W.; Linhardt, R.J.; et al. Fucosylated chondroitin sulfate from Isostichopus badionotus alleviates metabolic syndromes and gut microbiota dysbiosis induced by high-fat and high-fructose diet. Int. J. Biol. Macromol. 2019, 124, 377–388. [Google Scholar] [CrossRef]
- Yuan, Y.; Li, C.; Zheng, Q.; Wu, J.; Zhu, K.; Shen, X.; Cao, J. Effect of simulated gastrointestinal digestion in vitro on the antioxidant activity, molecular weight and microstructure of polysaccharides from a tropical sea cucumber (Holothuria leucospilota). Food Hydrocoll. 2019, 89, 735–741. [Google Scholar] [CrossRef]
- Zhao, F.; Liu, Q.; Cao, J.; Xu, Y.; Pei, Z.; Fan, H.; Yuan, Y.; Shen, X.; Li, C. A sea cucumber (Holothuria leucospilota) polysaccharide improves the gut microbiome to alleviate the symptoms of type 2 diabetes mellitus in Goto-Kakizaki rats. Food Chem. Toxicol. 2020, 135, 110886. [Google Scholar] [CrossRef]
- Zhu, Q.; Lin, L.; Zhao, M. Sulfated fucan/fucosylated chondroitin sulfate-dominated polysaccharide fraction from low-edible-value sea cucumber ameliorates type 2 diabetes in ats: New prospects for sea cucumber polysaccharide based-hypoglycemic functional food. Int. J. Biol. Macromol. 2020, 159, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Gong, P.; Li, Q.; Wu, Y.; Lu, W.; Zeng, J.; Li, H. Structural elucidation and antidiabetic activity of fucosylated chondroitin sulfate from sea cucumber Stichopus japonicas. Carbohydr. Polym. 2021, 262, 117969. [Google Scholar] [CrossRef] [PubMed]
- Gong, P.X.; Wu, Y.C.; Liu, Y.; Lv, S.Z.; You, Y.; Zhou, Z.L.; Chen, X.; Li, H.J. Structure and hypoglycemic effect of a neutral polysaccharide isolated from sea cucumber Stichopus japonicus. Int. J. Biol. Macromol. 2022, 216, 14–23. [Google Scholar] [CrossRef]
- Hu, S.; Chen, S.; Zhu, H.; Du, M.; Jiang, W.; Liu, Y.; Gao, X.; Su, L.; Xu, Y. Low molecular weight, 4-O-sulfation, and sulfation at meta-fucose positively promote the activities of sea cucumber fucoidans on improving insulin resistance in HFD-fed mice. Mar. Drugs 2022, 20, 37. [Google Scholar] [CrossRef] [PubMed]
- Vo, T.; Kim, S. Fucoidans as a natural bioactive ingredient for functional foods. J. Funct. Foods 2013, 5, 16–27. [Google Scholar] [CrossRef]
- Abioye, R.O.; Okagu, I.U.; Udenigwe, C.C. Targeting glucose transport proteins for diabetes management: Regulatory roles of food-derived compounds. J. Agric. Food Chem. 2022, 70, 5284–5290. [Google Scholar] [CrossRef]
- Khotimchenko, Y. Pharmacological potential of sea cucumbers. Int. J. Mol. Sci. 2018, 19, 1342. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, R.J.C.; Mourão, P.A.S. Pharmacological activities of sulfated fucose-rich polysaccharides after oral administration: Perspectives for the development of new carbohydrate-based drugs. Mar. Drugs 2021, 19, 425. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.Y.; Nguyen, T.H.; Kurihara, H.; Kim, S.M. α-glucosidase inhibitory activity of bromophenol purified from the red alga Polyopes lancifolia. J. Food Sci. 2010, 75, H147–H150. [Google Scholar] [CrossRef]
- Cho, M.; Han, J.H.; You, S. Inhibitory effects of fucan sulfates on enzymatic hydrolysis of starch. Food Sci. Technol. 2011, 44, 1164–1171. [Google Scholar] [CrossRef]
- Samuel, V.T.; Shulman, G.I. The pathogenesis of insulin resistance: Integrating signaling pathways and substrate flux. J. Clin. Investig. 2016, 126, 12–22. [Google Scholar] [CrossRef] [Green Version]
- Ho, C.K.; Sriram, G.; Dipple, K.M. Insulin sensitivity predictions in individuals with obesity and type II diabetes mellitus using mathematical model of the insulin signal transduction pathway. Mol. Genet. Metab. 2016, 119, 288–292. [Google Scholar] [CrossRef]
- Hu, S.; Chang, Y.; Wang, J.; Xue, C.; Shi, D.; Xu, H.; Wang, Y. Fucosylated chondroitin sulfate from Acaudina molpadioides improves hyperglycemia via activation of PKB/GLUT4 signaling in skeletal muscle of insulin resistant mice. Food Funct. 2013, 4, 1639–1646. [Google Scholar] [CrossRef]
- Agarwal, S.; Singh, V.; Chauhan, K. Antidiabetic potential of seaweed and their bioactive compounds: A review of developments in last decade. Crit. Rev. Food Sci. Nutr. 2022, 1–32. [Google Scholar] [CrossRef]
- Li, Y.; Liu, Y.; Liang, J.; Wang, T.; Sun, M.; Zhang, Z. Gymnemic acid ameliorates hyperglycemia through PI3K/AKT-and AMPK-mediated signaling pathways in type 2 diabetes mellitus rats. J. Agric. Food Chem. 2019, 67, 13051–13060. [Google Scholar] [CrossRef] [PubMed]
- Reaven, G.M. Role of insulin resistance in human disease. Diabetes 1988, 37, 1595–1607. [Google Scholar] [CrossRef]
- Wan, Y.; Xu, X.; Gilbert, R.G.; Sullivan, M.A. A review on the structure and anti-diabetic (type 2) functions of β-glucans. Foods 2022, 11, 57. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.J.; Kwon, M.J.; Kim, I.H.; Nam, T.J. The effect of polysaccharide extracted from the marine alga Capsosiphon fulvescens on ethanol administration. Food Chem. Toxicol. 2008, 46, 2653–2657. [Google Scholar] [CrossRef] [PubMed]
- Nakavachara, P.; Kajchamaporn, W.; Pooliam, J.; Viprakasit, V. Early development of decreased β-cell insulin secretion in children and adolescents with hemoglobin H disease and its relationship with levels of anemia. Pediatr. Blood Cancer 2020, 67, e28109. [Google Scholar] [CrossRef]
- Woldeamlak, B.; Yirdaw, K.; Biadgo, B. Role of gut microbiota in type 2 diabetes mellitus and its complications: Novel insights and potential intervention strategies. Korean J. Gastroenterol. 2019, 74, 314. [Google Scholar] [CrossRef]
- Lu, J.; Ma, K.L.; Ruan, X.Z. Dysbiosis of gut microbiota contributes to the development of diabetes mellitus. J. Infect. Dis. 2019, 1, 43–48. [Google Scholar] [CrossRef]
- Adeshirlarijaney, A.; Gewirtz, A.T. Considering gut microbiota in treatment of type 2 diabetes mellitus. Gut Microbes 2020, 11, 253–264. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Qin, J.; Cheng, Y.; Lv, D.; Li, M.; Qi, Y.; Lan, J.; Zhao, Q.; Li, Z. Marine sulfated polysaccharides: Preventive and therapeutic effects on metabolic syndrome: A Review. Mar. Drugs 2021, 19, 608. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Yoon, K.; Lee, B. Fucoidan regulate blood glucose homeostasis in C57BL/KSJ m+/+db and C57BL/KSJ db/db mice. Fitoterapia 2012, 83, 1105–1109. [Google Scholar] [CrossRef]
- Xing, M.; Cao, Q.; Wang, Y.; Xiao, H.; Zhao, J.; Zhang, Q.; Ji, A.; Song, S. Advances in research on the bioactivity of alginate oligosaccharides. Mar. Drugs 2020, 18, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, C.; Hao, J.; Wang, W.; Han, Z.; Li, G.; Zhang, L.; Zhao, X.; Yu, G. Insulin sensitizing effects of oligomannuronate-chromium (III) complexes in C2C12 skeletal muscle cells. PLoS ONE 2011, 6, e24598. [Google Scholar] [CrossRef]
- Huang, L.; Shen, M.; Morris, G.A.; Xie, J. Sulfated polysaccharides: Immunomodulation and signaling mechanisms. Trends Food Sci. Technol. 2019, 92, 1–11. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, Z.; Yang, L.; Yang, X.; Wang, X.; Zhang, Z. In vitro antioxidant activities of sulfated derivatives of polysaccharides extracted from Auricularia auricular. Int. J. Mol. Sci. 2011, 12, 3288–3302. [Google Scholar] [CrossRef]
- Gunasekaran, S.; Govindan, S.; Ramani, P. Sulfated modification, characterization and bioactivities of an acidic polysaccharide fraction from an edible mushroom Pleurotus eous (Berk.) Sacc. Heliyon 2021, 7, e5964. [Google Scholar] [CrossRef]
- Rizkyana, A.D.; Ho, T.C.; Roy, V.C.; Park, J.; Kiddane, A.T.; Kim, G.; Chun, B. Sulfation and characterization of polysaccharides from Oyster mushroom (Pleurotus ostreatus) extracted using subcritical water. J. Supercrit. Fluids 2022, 179, 105412. [Google Scholar] [CrossRef]
- Wang, Z.; Xie, J.; Shen, M.; Nie, S.; Xie, M. Sulfated modification of polysaccharides: Synthesis, characterization and bioactivities. Trends Food Sci. Technol. 2018, 74, 147–157. [Google Scholar] [CrossRef]
- Kang, J.; Jia, X.; Wang, N.; Xiao, M.; Song, S.; Wu, S.; Li, Z.; Wang, S.; Cui, S.W.; Guo, Q. Insights into the structure-bioactivity relationships of marine sulfated polysaccharides: A review. Food Hydrocoll. 2022, 123, 107049. [Google Scholar] [CrossRef]
- Huang, W.; Tan, H.; Nie, S. Beneficial effects of seaweed-derived dietary fiber: Highlights of the sulfated polysaccharides. Food Chem. 2022, 373, 131608. [Google Scholar] [CrossRef] [PubMed]
- Caputo, H.E.; Straub, J.E.; Grinstaff, M.W. Design, synthesis, and biomedical applications of synthetic sulphated polysaccharides. Chem. Soc. Rev. 2019, 48, 2338–2365. [Google Scholar] [CrossRef]
- Koh, H.; Lu, J.; Zhou, W. Structure characterization and antioxidant activity of fucoidan isolated from Undaria pinnatifida grown in New Zealand. Carbohydr. Polym. 2019, 212, 178–185. [Google Scholar] [CrossRef]
- Wu, S.; Liu, Y.; Jiang, P.; Xu, Y.; Zheng, W.; Song, S.; Ai, C. Effect of sulfate group on sulfated polysaccharides-induced improvement of metabolic syndrome and gut microbiota dysbiosis in high fat diet-fed mice. Int. J. Biol. Macromol. 2020, 164, 2062–2072. [Google Scholar] [CrossRef]
- Guo, X.; Ye, X.; Sun, Y.; Wu, D.; Wu, N.; Hu, Y.; Chen, S. Ultrasound effects on the degradation kinetics, structure, and antioxidant activity of sea cucumber fucoidan. J. Agr. Food Chem. 2014, 62, 1088–1095. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Li, S.; Li, J.; Ye, X.; Ding, T.; Liu, D.; Chen, J.; Ge, Z.; Chen, S. Identification of a highly sulfated fucoidan from sea cucumber Pearsonothuria graeffei with well-repeated tetrasaccharides units. Carbohydr. Polym. 2015, 134, 808–816. [Google Scholar] [CrossRef]
- Li, S.; Li, J.; Zhi, Z.; Wei, C.; Wang, W.; Ding, T.; Ye, X.; Hu, Y.; Linhardt, R.J.; Chen, S. Macromolecular properties and hypolipidemic effects of four sulfated polysaccharides from sea cucumbers. Carbohydr. Polym. 2017, 173, 330–337. [Google Scholar] [CrossRef]
- Li, S.; Li, J.; Zhi, Z.; Hu, Y.; Ge, J.; Ye, X.; Tian, D.; Linhardt, R.J.; Chen, S. 4-O-Sulfation in sea cucumber fucodians contribute to reversing dyslipidiaemia caused by HFD. Int. J. Biol. Macromol. 2017, 99, 96–104. [Google Scholar] [CrossRef] [Green Version]
- Agyekum, I.; Pepi, L.; Yu, Y.; Li, J.; Yan, L.; Linhardt, R.J.; Chen, S.; Amster, I.J. Structural elucidation of fucosylated chondroitin sulfates from sea cucumber using FTICR-MS/MS. Eur. J. Mass Spectrom. 2018, 24, 157–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Zhou, Q.; Liu, B.; Chen, F.; Wang, M. Holothurian fucosylated chondroitin sulfates and their potential benefits for human health: Structures and biological activities. Carbohydr. Polym. 2022, 275, 118691. [Google Scholar] [CrossRef]
- Chen, S.; Li, G.; Wu, N.; Guo, X.; Liao, N.; Ye, X.; Liu, D.; Xue, C.; Chai, W. Sulfation pattern of the fucose branch is important for the anticoagulant and antithrombotic activities of fucosylated chondroitin sulfates. Biochim. Biophys. Acta. Bioenerg. 2013, 1830, 3054–3066. [Google Scholar] [CrossRef] [PubMed]
- Ustyuzhanina, N.E.; Bilan, M.I.; Dmitrenok, A.S.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. The structure of a fucosylated chondroitin sulfate from the sea cucumber Cucumaria frondosa. Carbohydr. Polym. 2017, 165, 7–12. [Google Scholar] [CrossRef]
- Yang, J.; Wang, Y.; Jiang, T.; Lv, Z. Novel branch patterns and anticoagulant activity of glycosaminoglycan from sea cucumber Apostichopus japonicus. Int. J. Biol. Macromol. 2015, 72, 911–918. [Google Scholar] [CrossRef]
- Mabate, B.; Daub, C.D.; Malgas, S.; Edkins, A.L.; Pletschke, B.I. Fucoidan structure and its impact on glucose metabolism: Implications for diabetes and cancer therapy. Mar. Drugs 2021, 19, 30. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Gao, L.; Zhou, H.; Ai, C.; Huang, X.; Wang, M.; Zhang, Y.; Zhao, C. Opportunities and challenges of algal fucoidan for diabetes management. Trends Food Sci. Technol. 2021, 111, 628–641. [Google Scholar] [CrossRef]
- Zhang, W.; Su, R.; Gong, L.; Yang, W.; Chen, J.; Yang, R.; Wang, Y.; Pan, W.; Lu, Y.; Chen, Y. Structural characterization and in vitro hypoglycemic activity of a glucan from Euryale ferox Salisb. seeds. Carbohydr. Polym. 2019, 209, 363–371. [Google Scholar] [CrossRef]
- Xiao, Z.; Zhou, W.; Zhang, Y. Fungal polysaccharides. Adv. Pharmacol. 2020, 87, 277–299. [Google Scholar] [CrossRef]
- Yan, M.; Fengzhi, L.; Xiaojuan, X.; Lina, Z. Recent progress in chain conformation and function of triple helical polysaccharides. Biomacromolecules 2020, 21, 1653–1677. [Google Scholar] [CrossRef]
- Zhang, L.; Li, X.; Xu, X.; Zeng, F. Correlation between antitumor activity, molecular weight, and conformation of lentinan. Carbohyd. Res. 2005, 340, 1515–1521. [Google Scholar] [CrossRef]
- Kony, D.B.; Damm, W.; Stoll, S.; van Gunsteren, W.F.; Hünenberger, P.H. Explicit-solvent molecular dynamics simulations of the polysaccharide schizophyllan in water. Biophys. J. 2007, 93, 442–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.; Rioux, L.; Turgeon, S.L. Alpha-amylase and alpha-glucosidase inhibition is differentially modulated by fucoidan obtained from Fucus vesiculosus and Ascophyllum nodosum. Phytochemistry 2014, 98, 27–33. [Google Scholar] [CrossRef]
- Katakami, N.; Osaka, U.G.S.O.; Department, O.M.A.A.; Department, O.M.M. Mechanism of development of atherosclerosis and cardiovascular disease in diabetes mellitus. J. Atheroscler. Thromb. 2018, 25, 27–39. [Google Scholar] [CrossRef] [Green Version]
- Zaporozhets, T.; Besednova, N. Prospects for the therapeutic application of sulfated polysaccharides of brown algae in diseases of the cardiovascular system: Review. Pharm. Biol. 2016, 54, 3126–3135. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Yim, E. Fucoidan for cardiovascular application and the factors mediating its activities. Carbohydr. Polym. 2021, 270, 118347. [Google Scholar] [CrossRef]
- Shang, F.; Mou, R.; Zhang, Z.; Gao, N.; Lin, L.; Li, Z.; Wu, M.; Zhao, J. Structural analysis and anticoagulant activities of three highly regular fucan sulfates as novel intrinsic factor Xase inhibitors. Carbohydr. Polym. 2018, 195, 257–266. [Google Scholar] [CrossRef]
- Tsai, S.; Wang, J.; Liao, W.; Hsu, Y.; Lin, C.; Liao, M.; Huang, P.; Lin, S. Fucoidan attenuates angiotensin II-induced abdominal aortic aneurysms through the inhibition of c-Jun N-terminal kinase and nuclear factor κB activation. J. Vasc. Surg. 2018, 68, 72S–81S. [Google Scholar] [CrossRef]
- Kuznetsova, T.A.; Ivanushko, L.A.; Persiyanova, E.V.; Ermakova, S.P.; Besednova, N.N. Markers of systemic inflammation in experimental dyslipidemia induced by P-407: Modulation with fucoidan from brown alga Fucus evanescens. Bull. Exp. Biol. Med. 2019, 166, 766–769. [Google Scholar] [CrossRef]
- Rübsam, A.; Parikh, S.; Fort, P. Role of inflammation in diabetic retinopathy. Int. J. Mol. Sci. 2018, 19, 942. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Zhang, Q.; Luo, D.; Wang, J.; Duan, D. Low molecular weight fucoidan modulates P-selectin and alleviates diabetic nephropathy. Int. J. Biol. Macromol. 2016, 91, 233–240. [Google Scholar] [CrossRef]
- Xu, J.; Wang, Y.; Wang, Z.; Guo, L.; Li, X. Fucoidan mitigated diabetic nephropathy through the downregulation of PKC and modulation of NF-κB signaling pathway: In vitro and in vivo investigations. Phytother. Res. 2021, 35, 2133–2144. [Google Scholar] [CrossRef] [PubMed]
- Mukhamejanov, E.; Kon, G.; Erjanova, S.; Kirgizbaeva, A.; Muhamadieva, E. Fucoidan—New principle prevention and teatment of diabetes. J. Pharm. Pharmacol. 2019, 7, 316–322. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhao, H.; Wang, Q.; Liang, H.; Jiang, X. Fucoidan protects ARPE-19 cells from oxidative stress via normalization of reactive oxygen species generation through the Ca2+-dependent ERK signaling pathway. Mol. Med. Rep. 2015, 11, 3746–3752. [Google Scholar] [CrossRef] [Green Version]
- Patil, N.P.; Gomez-Hernandez, A.; Zhang, F.; Cancel, L.; Feng, X.; Yan, L.; Xia, K.; Takematsu, E.; Yang, E.Y.; Le, V.; et al. Rhamnan sulfate reduces atherosclerotic plaque formation and vascular inflammation. Biomaterials 2022, 291, 121865. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Diabetes | Etiology and Pathogenesis |
|---|---|
| Type I diabetes mellitus (T1DM) | Normal β-cells and antibodies in the body are produced during an immune reaction, causing β-cells to be destroyed, which causes the body to not to be able to secrete the insulin, which, in turn, causes an absolute lack of insulin, leading to abnormal metabolic disease. |
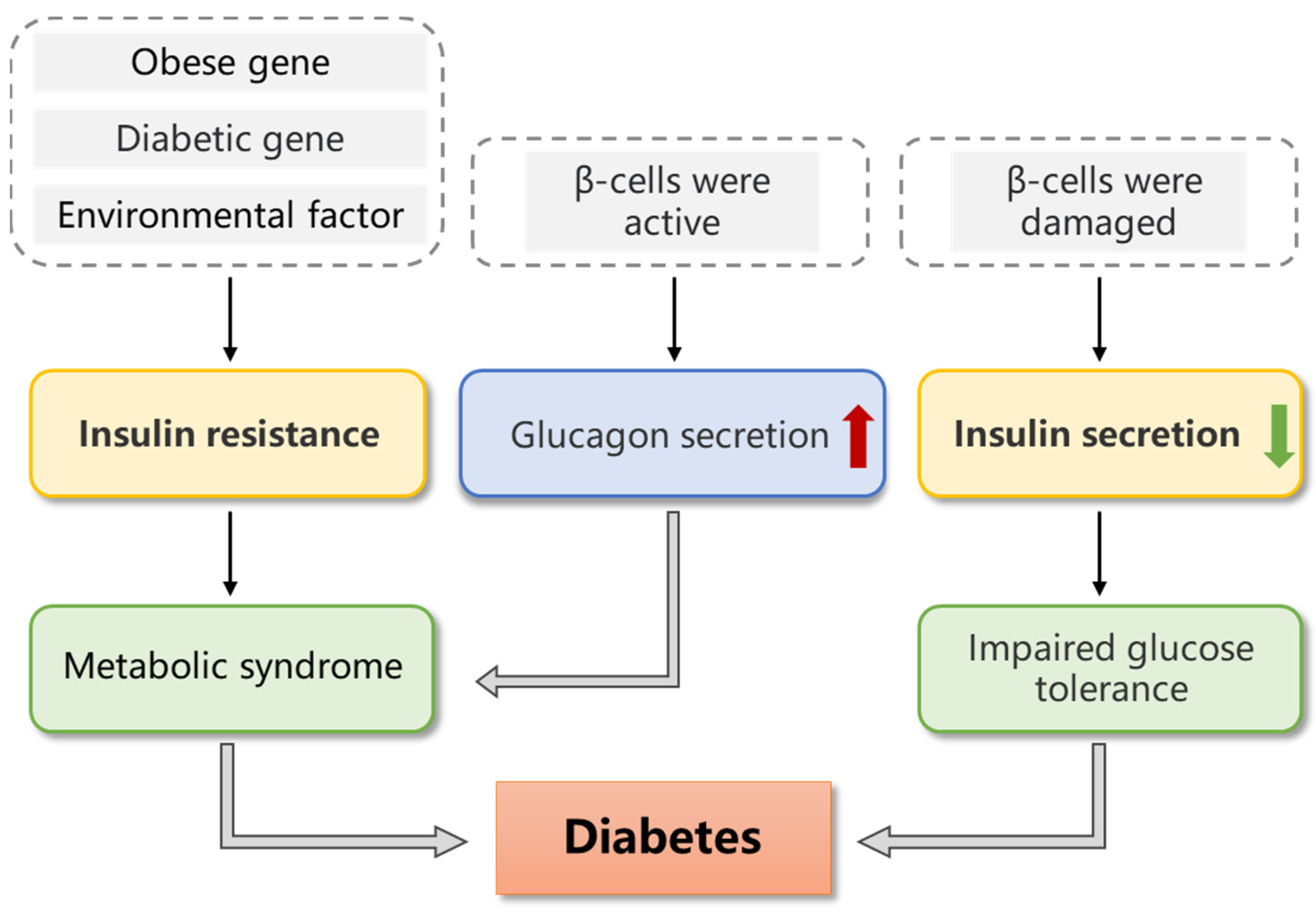
| Type II diabetes mellitus (T2DM) | (1) A genetic defect in insulin resistance caused by abnormal gene regulation. (2) A genetic defect in beta cells that prevents them from properly secreting insulin. Epigenetic environmental factors, such as obesity, aging, stress, dietary habits, glucose toxicity and lipotoxicity, oxidative stress and endoplasmic reticulum stress, are also closely associated with T2DM. |
| Gestational diabetes mellitus (GDM) | As a woman gains weight and reduces their physical activity during pregnancy, peripheral insulin resistance develops, and glucose intolerance may occur. This, in turn, undermines pancreatic β-cell function and may contribute to the increased risk of GDM. Behavioral factors, excess weight and inherited genes are associated with insulin resistance, and there is a risk of its further development into T2DM. |
| Special types of diabetes mellitus | According to their etiology, they are classified into eight major categories: genetic defects of the β-cell, genetic defects in insulin action, diseases of the exocrine pancreas, endocrinopathies, drug- or chemical-induced diabetes, infections, uncommon forms of immune-mediated diabetes and other genetic syndromes sometimes associated with diabetes. |
| No. | Name | Source | Structural Information | Effect | Reference | ||||
|---|---|---|---|---|---|---|---|---|---|
| Molecular Weight (kDa) | Monosaccharide Composition | Sulfate Group Content (%) | Structure | ||||||
| 1 | LMWF | Undaria pinnatifida | Brown algae | 1 ± 0.2 | - | - | - | Improve glucose homeostasis and insulin resistance due to endoplasmic reticulum stress (in vivo) | [38] |
| 2 | - | Ascophyllum nodosum | Brown algae | 637 | Fucose/Galactose/Xylose/ Mannose/Glucuronic acid = 31.1/4.1/6.4/2.9/2.8 | 20.6 | Fucosyl residue/Galactosyl residue/Xyloxyl residue = 82.9/1.2/13.9 | The inhibition rate of α-amylase was 83.2% (in vitro) | [39] |
| 3 | STP-1 | Sargassum thunbergii | Brown algae | 190.4 | Arabinose/Galactose/Gluco-se/Xylose/Mannose/Galact-uronic acid/Glucuronic acid = 1.94/30.7/4.54/23.2/17.6/8.11/13.9 | 15.2 | - | The inhibitory rate of α-glucosidase was 75.0%, which increased the glucose consumption of HepG2 cells (in vitro) | [40] |
| 4 | - | Kappaphycus alvarezii | Red algae | - | - | - | →4)-4-O-sulfonato- (2-O-methyl)-β-D-galactopyranosyl-(1→4)-3,6-anhydro-(2-O-methyl)-α-D-galactopyranan | The IC50 of α-amylase and α-glucosidase were 0.15 mg/mL and 0.09 mg/mL, respectively (in vitro) | [41] |
| 5 | - | Gracilaria opuntia | Red algae | - | - | - | →3)-4-O-sulfonato-(6-O-acetyl)-β-D-galactopyranosyl-(1→4)-3,6- anhydro-(2-O-sulfonato)-α-D-galactopyranosyl-(1→3)-4-O-sulfonato-(6-O-acetyl)-β-D-xylosyl-(1→3)-4-O-sulfonato-(6-O-acetyl)-β-D-galactopyranosyl-(1→4)-3,6-anhydro-(2-O-sulfonato)-α-D-galactopyranan | The IC50 of α-amylase and α-glucosidase were 0.04 mg/mL and 0.09 mg/mL, respectively (in vitro) | [41] |
| 6 | GLP | Gracilaria lemaneiformis | Red algae | 21.2 | Rhamnose/Arabinose/ Xylose/Mannose/Glucose/Galactose/Uronic acids | 19.64 | - | The IC50 of α-amylase is 3.94 μg/mL, which can regulate glucose and lipid metabolism, repair β-cells, protect liver and kidney function, and promote the activity of endogenous antioxidant enzymes (in vitro and in vivo) | [42] |
| 7 | GLPs-SeNPs | G. lemaneiformis | Red algae | 382.3 | Fucose/Galactose/Glucose/Xylose/Ribose/Glucuronic acid | The IC50 of the α-amylase was 1.550 mg/mL and the IC50 of α-glucosidase was 2.139 mg/mL (in vitro) | [43] | ||
| 8 | SPP-1 | Sargassum pallidum | Brown algae | 1518.6 | Fucose/Arabinos/Galactose/Mannose/Glucose/Xylose/Glucuronic acid/Galacturonic acid = 4.97/1.00/9.75/6.44/1.71/1.82/6.07/2.20 | 2.61 | - | α-amylase and -glucosidase inhibitory activities, and remarkably improve glucose consumption in insulin resistance (IR) model cells (in vitro) | [44] |
| 9 | PSP-2 | S. pallidum | Brown algae | 144.8 | Fucose/Arabinose/Galactos-e/Glucose/Xylose/Mannose/Galacturonic acid/Glucuronic acid = 21.6/2.5/22.4/2.2/18.8/1.2/7.7/23.6 | 4.71 | →1)-β-D-Xylp-(3→, →1,3)-β-L-Fucp-(4→, →1)-α-D-Galp-(6→, and →1)-α-D-GlcpNAc-(2→, and the side chains were composed of →1,3,6)-α-D-Galp-(2→, →3)-β-L-Fucp-(1,4→, β-D-GalpNAc-(1→, and α-D-Manp-(1→ | Improve insulin resistance HepG2 cell glucose consumption, promote glycogen synthesis and improve insulin resistance (in vitro) | [45] |
| 10 | S-SPP1-8 | S. pallidum | Brown algae | 1734 | Fucose/Arabinose/Galactos-e/Glucose/Xylose/Mannose/Galacturonic acid/Glucuronic acid = 51.72/5.50/13.74/0.81/1.32/6.60/10.31/9.99 | 13.36 | - | The inhibition rate of α-glucosidase was 98.4%, improve insulin resistance (in vitro) | [46] |
| 11 | PSP-1 | S. pallidum | Brown algae | 1036 | Fucose/Arabinose/Galactos-e/Glucose/Xylose/Mannose/Galacturonic acid/Glucuronic acid = 18.45/2.15/19.06/1.89/16.07/1.00/5.74/20.09 | 10.69 | - | It has certain inhibitory effect onα-amylase and -glucosidase, improve insulin resistance HepG2 cell glucose consumption (in vitro) | [47] |
| 12 | Se-SPP | S. pallidum | Brown algae | 3610, 1630 | Fucose/Arabinose/Galactos-e/Glucose/Xylose/Mannose/Galacturonic acid/Glucuronic acid = 32.13/1.15/19.39/21.0/4.42/11.61/1.99/8.31 | - | - | The IC50 of α-glucosidase were 0.896 mg/mL (in vitro) | [48] |
| 13 | SCO | Sargassum confusum | Brown algae | - | Fucose/Arabinose/Xylose/Glucose | - | SCO was a sulfated oligosaccharide containing one Gal unit and one anGal unit, sulfated galactose, sulfated anhydrogalactose and methyl sulfated galactoside units. | Improve insulin resistance and regulate intestinal microflora (in vivo) | [49] |
| 14 | SFP-2 | Sargassum fusiforme | Brown algae | 84.99, 14.33 | Fucose/Mannose/Rhamnose/Glucose/Galactose/Glucuronic acid = 41.22/16.79/2.65/6.67/19.27/13.40 | 3.24 | - | Improve hyperinsulinemia and insulin resistance, regulate intestinal microflora (in vivo) | [50,51] |
| 15 | SFF | S. fusiforme | Brown algae | - | Mannose/Rhamnose/ Glucose/Glucuronic acid/Galacturonic acid/Galactose/Xylose/Fucose = 10.89/3.29/4.32/4.53/14.02/18.33/3.57/41.05 | 17.36 | - | Improve insulin resistance and sensitivity, reduce fasting blood glucose and IR index along with improve glucose tolerance, increase the abundance and diversity of gut microbiota, improve intestinal integrity and inflammation (in vivo) | [52] |
| 16 | - | Ecklonia maxima | Brown algae | 10 | Fructose/Fucose/Galactose/Glucose/Mannose/Xylose = 12.78/4.45/1.44/1.09/4.30/0.79 | 6.01 | - | The range of α-glucosidase IC50 is 0.27–0.31 mg/mL (in vitro) | [53] |
| 17 | PD-1 | Porphyra spp. | Red algae | 2.59 | Galactose/Glucose = 98.6/1.4 | - | - | The IC50 of α-amylase was 12.72 mg/mL, and the inhibitory activity of α-amylase was about 98.78% (in vitro) | [54] |
| 18 | EPs | Porphyridium cruentum | Red algae | - | - | - | - | The inhibition rate of α-glucosidase was 71.57%, increase the number of pancreatic beta cells (in vitro and in vivo) | [55] |
| 19 | Up4 | U. pinnatifida | Brown algae | 41.4 | Mannose/Rhamnose/Galact-ose/Fucose/Glucuronic acid | 8.74 | Both α-configuration and β-configuration exist | The IC50 of α-glycosidase was 50.5 µg/mL, improve insulin resistance HepG2 cell glucose consumption, Lower fasting blood glucose, improve glucose metabolism disorder, improve insulin sensitivity, increase liver glycogen synthesis (in vitro and in vivo) | [56] |
| 20 | MAP | Macrocystis pyrifera | Brown algae | 472.2, 137.6, 26.8 | Fucose/Mannose/Rhamnose/Glucose/Galactose/Xylose/Glucuronic acid = 27.75/26.43/2.14/1.11/6.54/18.77/17.26 | 7.18 | - | It has a positive effect on the control of LDL-C level in diabetic rats, regulation of glucose metabolism and intestinal microflora (in vivo) | [57,58] |
| 21 | SFF | S. fusiforme | Brown algae | 205.8 | Fucose/Mannose/Rhamnose/Glucose/Galactose/Xylose = 55.67/4.45/3.34/5.44/20.83/3.70 | 14.55 | - | Reduce fasting blood glucose, inhibit oxidative stress, regulate intestinal microflora (in vivo) | [59] |
| 22 | UPP | U. pinnatifida | Brown algae | 185.5 | Fucose/Galactose/ Glucuronic acid/Mannose/Glucose/ Rhamnose/Galacturonic acid = 33.46/29.49/19.17/12.30/2.06%/1.94%/1.58% | - | - | Reduce the level of fasting blood glucose, relieve insulin resistance and regulate the abundance of intestinal microflora (in vivo) | [60] |
| 23 | PDA4 | - | Brown algae | 40 | - | - | M/G = 1.8, it appears as random coil and compact spherical coil in solution | It has the ability of glucose adsorption and diffusion inhibition (in vitro) | [61] |
| 24 | SFP-7-40 | S. fusiforme | Brown algae | 41.27 | Fucose/Mannose/Rhamnose/Xylose/Glucuronic acid = 2.5/33.20/2.5/18.02/43.78 | 32.81 | - | The IC50 of α-glycosidase was 0.304 mg/mL (in vitro) | [62] |
| 25 | CDDP | Dictyopteris divaricata | Brown algae | 63.06 | Mannose/Ribose/ Rhamnose/Glucuronic acid/Glucose/Galactose/ Xylose/Arabinose/Fucose = 15.02/9.90/1.28/17.54/1.86/17.19/4.54/0.55/32.13 | - | - | Improves fasting blood glucose and insulin abnormalities, modulates the gut microbiota and maintains the integrity of the gut barrier (in vivo) | [63] |
| 26 | ULP-1 | Ulva lactuca | Green algae | 62.12 | Mannose/Rhamnose/ Glucuronic acid/Glucose/Galactose/Arabinose/Xylose = 0.22/22.88/9.41/0.44/0.50/3.44/0.60 | 8.99 | (ULP-1) comprised β-D-Xylp-(1→3)-β-D-Arap-(1→6)-β-D-Galp-(1→6)-β-D-Glcp linked to [→α-L-Rhap-(1→4)-β-D-GlcpA→]n and α-D-Manp-(1→4)-α-L-Rhap(2SO3-)-(1→2)-α-L-Rhap(4SO3)-(1→2)-α-L-Arap-(1→2)-α-L-Rhap-(1→as its side chains at β-D-Glcp | Improve glucose tolerance and regulate intestinal microflora (in vivo) | [64] |
| 27 | LMWAs-H | A. Nodosum | Brown algae | 33.48 | - | - | →4)-α-L-Fucp-(1→4)-α-L-Fucp-(1→3)-β-D-Xylp-(1→3)-α-L-Fucp4S(1→ as main chain, and T-α-D-Glcp-(1→ and →3)-β-D-ManpAred residues were attached to the ends of main chain as non-reducing- and reducing-end residues, respectively, the 4-deoxy-L-erythro-hex-4-enuronosyluronate linked the O-4 position of →3,4)-β-D-ManpAred residue as side branches | The IC50 values of α-amylase and α-glucosidase were 1150 ± 10 μg/mL and 560 ± 10 μg/mL, respectively (in vitro) | [65] |
| 28 | LF2 | Laminaria japonica | Brown algae | 7.2 | Fucose/Mannose/Rhamnose/Xylose/Galactosamine/Gl-ucose/Galactose/Arabinose | 29.3 | The backbone was (1→3)-linked α-L-fucopyranose residues and a few (1→4)-α-L-fucopyranose linkages. The branch points were at C-4 of 3-linked α-L-fucopyranose residues by β-D-galactopyranose unites or at C-2 of 3-linked α-L-fucopyranose residues by non-reducing terminal fucose unites | Reduce fasting blood glucose, improve insulin secretion and metabolic syndrome, regulation of intestinal microflora (in vivo) | [66] |
| 29 | PSP3 | Spirulina platensis | Blue-green algae | 10–30 | - | 12.01 | - | The IC50 of α-glycosidase was 0.85 mg/mL, improve oral glucose tolerance and insulin resistance (in vitro and in vivo) | [67] |
| 30 | - | U. pinnatifida | Brown algae | - | Fucose/Galactose/Glucose/Glucuronic acid | 22.83 | The backbone was a repeating structure of alternatively linked α-(1–3) and α-(1–4) fucose and galactose units, with a high degree of sulfation | Inhibit α-glycosidase and α-amylase activities (in vitro) | [68] |
| 31 | FvF | Fucus vesiculosus | Brown algae | - | Fucose/Galactose/ Xylose/Glucose | - | 1→3/1→4 Linkage mode | The IC50 of α-glycosidase was 67.9 μg/mL (in vitro) | [69] |
| 32 | Am-FUC | Acaudina molpadioides | Sea cucumber | 1614.1 | Fucose | 26.3 | [→3-α-L-Fucp-1→3-α-L-Fucp2,4(OS3−)-1→3-α-L-Fucp-1→3-α-L-Fucp2(OS3−)-1]n | Improve hyperglycemia and insulin resistance, regulates intestinal microflora (in vivo) | [70,71] |
| 33 | Cf-CHS | Cucumaria frondosa | Sea cucumber | 14.76 | Glucuronic acid/Galactosamine/Fucose | 30.07 | The backbone was CHS E, (4-β-D-GlcA-1→3-β-D-GalNAc)n | Improve insulin sensitivity and insulin resistance, repair pancreatic islets apoptosis (in vivo) | [72,73,74] |
| 34 | CHS | A. molpadioides | Sea cucumber | 21.53 | Glucuronic acid/Galactosamine/Fucose | 27.81 | - | Increased insulin signaling pathway, improve glucose metabolism (in vivo) | [75] |
| 35 | Ib-FUC | Isostichopus badionotus | Sea cucumber | 450 | Fucose | 32.9 | [→3Fuc(2S,4S)α1→3Fuc(2S)α1→ 3Fuc(2S)α1→3Fucα1→]n | Improve insulin resistance and inhibite inflammatory response (in vivo) | [76] |
| 36 | fuc-Pg | Pearsonothuria graeffei | Sea cucumber | - | Fucose | - | - | Regulates metabolism and intestinal microflora (in vivo) | [77] |
| 37 | AHG | Apostichopus japonicus | Sea cucumber | 98.07 | Glucuronic acid/N-acetylgalactosamine/Fucose | 33.2 | The backbone structure of→4)GlcUAβ(1→3)GalNAcβ(1→, with O-4 and/or O-6 positions of sulfation. The sulfated fucose branches occurred at the O-3 position of the D-GlcUA moiety or the O-4/6 position of D-GalNAc | Improve insulin resistance (in vivo) | [78] |
| 38 | fCS-Ib | I. badionotus | Sea cucumber | 10.9 | Glucuronic acid/N-acetylgalactosamine/Fucose | - | - | Reduce fasting blood glucose, reduce inflammation, regulate intestinal microflora (in vivo) | [79] |
| 39 | HLP | Holothuria leucospilota | Sea cucumber | 52.80 | Rhamnose/Fucose/Glucuro-nic acids/Galactose/Glucose/Xylose = 39.08/35.72/10.72/8.43/4.23/1.83 | - | - | Lower fasting blood glucose, regulate metabolic pathways and intestinal microflora (in vivo) | [80,81] |
| 40 | TAPF | Thelenota ananas | Sea cucumber | 1284 | Fucose | 13.15 | [→3-α-L-Fucp-1→3-α-L-Fucp-1→3α-L-Fucp2, 4 (OSO3−)-1→3-α-L-Fucp2 (OSO3−)-1→]n | Lower fasting blood glucose, improve glucose tolerance, promote insulin secretion or enhance insulin sensitivity, improve insulin resistance and promote liver glycogen accumulation (in vivo) | [82] |
| 41 | CFPF | C. frondosa | Sea cucumber | 30 | Glucuronic acid/N-acetylgalactosamine/Fucose | 8.21 | →3)-β-D-GalNAc4S6S-(1→4)-β-D-GlcA3S-(1→ and →3)-β-D-GalNAc4S-(1→4) -β-D-GlcA3S-(1→ | ||
| 42 | FCSsj | Stichopus japonicas | Sea cucumber | 60.99 | Glucuronic acid/Galactosamine/Fucose | 3.71 | - | Improve glucose uptake and glucose consumption of HepG2 cells and promote glycogen synthesis of HepG2 cells (in vitro) | [83] |
| 43 | NPsj | Stichopus japonicus | Sea cucumber | 301.75 | Glucose | - | (1→4)-α-D-glucoses with β-D-glucose (1→) branches substituted at O-6 every 7–9 of 1, 4 linked glucoses | Improve glucose uptake and glucose consumption of HepG2 cells and 3 T3-L1 cells model, improve insulin resistance (in vitro) | [84] |
| 32 | Am-FUC | A. molpadioides | Sea cucumber | 1614.1 | Fucose | 26.3 | [→3-α-L-Fucp-1→3-α-L-Fucp2,4(OS3−)-1→3-α-L-Fucp-1→3-α-L-Fucp2(OS3−)-1]n | Lower fasting blood glucose, eliminate insulin resistance (in vivo) | [85] |
| 44 | Ib-FUC | I. badionotus | Sea cucumber | 435.3 | Fucose | 32.9 | [→3-α-L-Fucp2(OS3−)-1→3-α-L-Fucp2,4(OS3−)-1→3-α-L-Fucp2(OS3−)-1→3-α-L-Fucp-1]n | ||
| 45 | Ta-FUC | T. ananas | Sea cucumber | 1380.0 | Fucose | 28.2 | [→3-α-L-Fucp2(OS3−)-1→3-α-L-Fucp2,4(OS3−)-1→3-α-L-Fucp-1→3-α-L-Fucp-1]n | ||
| 46 | Ht-FUC | Holothuria tubulosa | Sea cucumber | 1567.6 | Fucose | 27.4 | [→3-α-L-Fucp-1→3-α-L-Fucp2,4(OS3−)-1→3-α-L-Fucp2(OS3−)-1→3-α-L-Fucp2(OS3−)-1]n | ||
| 47 | Pg-FUC | P. graeffei | Sea cucumber | 310.5 | Fucose | 28.2 | [→3-α-L-Fucp-1→3-α-L-Fucp2,4(OS3−)-1→3-α-L-Fucp-1→3-α-L-Fucp4(OS3−)-1]n | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, L.; Xiao, M.; Cai, S.; Mou, H.; Li, D. Potential Application of Marine Fucosyl-Polysaccharides in Regulating Blood Glucose and Hyperglycemic Complications. Foods 2023, 12, 2600. https://doi.org/10.3390/foods12132600
Tang L, Xiao M, Cai S, Mou H, Li D. Potential Application of Marine Fucosyl-Polysaccharides in Regulating Blood Glucose and Hyperglycemic Complications. Foods. 2023; 12(13):2600. https://doi.org/10.3390/foods12132600
Chicago/Turabian StyleTang, Luying, Mengshi Xiao, Shenyuan Cai, Haijin Mou, and Dongyu Li. 2023. "Potential Application of Marine Fucosyl-Polysaccharides in Regulating Blood Glucose and Hyperglycemic Complications" Foods 12, no. 13: 2600. https://doi.org/10.3390/foods12132600