
Study on Characteristics and Lignification Mechanism of Postharvest Banana Fruit during Chilling Injury
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. Physiological Parameters
2.3. Statistical Analysis
3. Results and Discussion
3.1. Chilling Symptoms
3.2. Oxidative Stress, O2− and H2O2
3.3. Peel Browning, PPO, POD and PAL
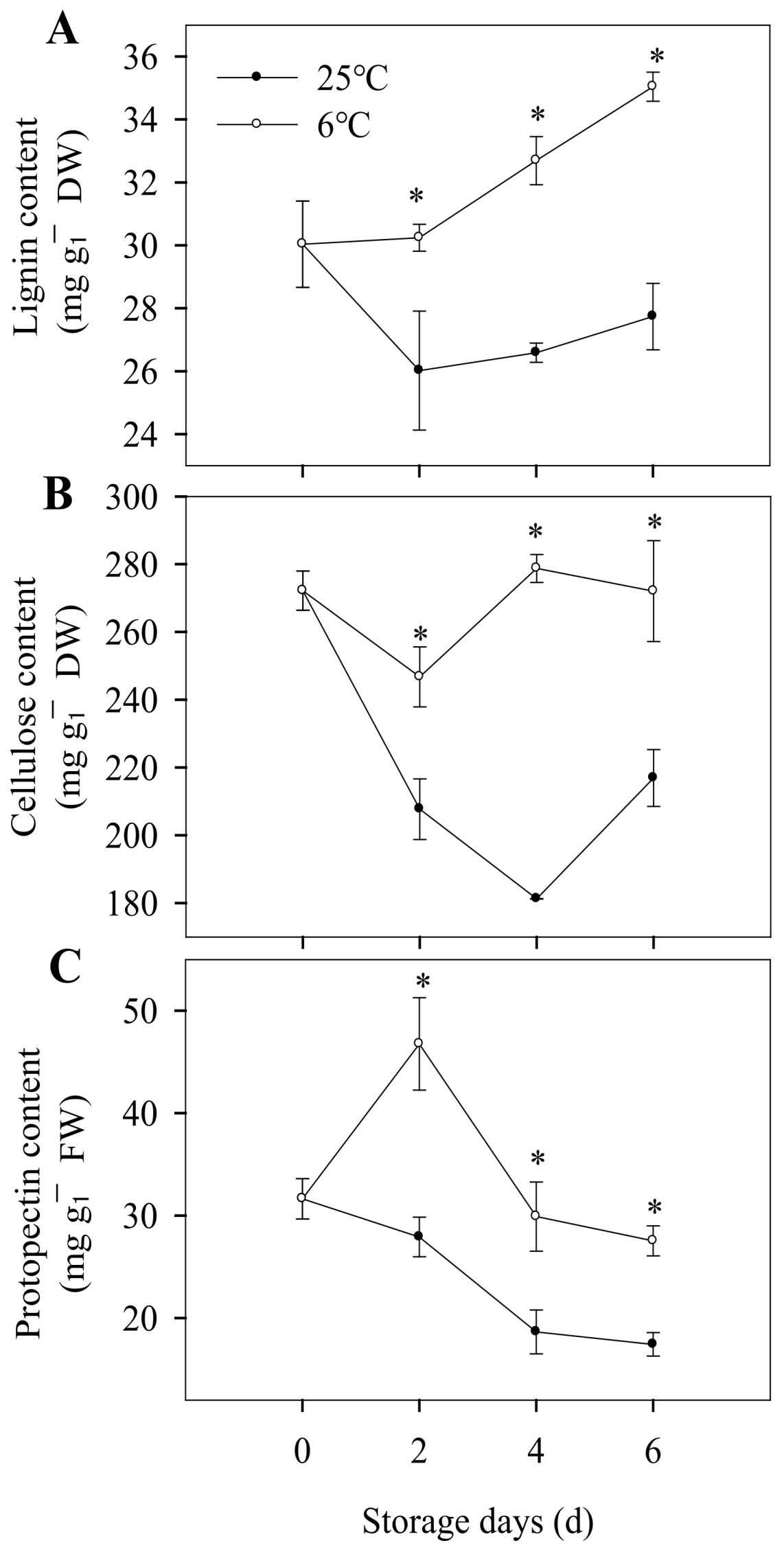
3.4. Cell Wall Metabolism, Content of Lignin, Cellulose and Protopectin, Activity of Cellulase and Pectinase
3.5. Ultrastructural Changes of Banana Peel after Chilling Injury
3.6. Expression Profiles of Many Genes Involved in Lignification Caused by CI
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sevillano, L.; Sanchez-Ballesta, M.T.; Romojaro, F.; Flores, F.B. Physiological, hormonal and molecular mechanisms regulating chilling injury in horticultural species. Postharvest technologies applied to reduce its impact. J. Sci. Food Agric. 2009, 89, 555–573. [Google Scholar] [CrossRef]
- Wang, C.Y. Chilling injury of tropical horticultural commodities. HortScience 1994, 29, 986–988. [Google Scholar] [CrossRef] [Green Version]
- Mulas, M.; Schirra, M. The effect of heat conditioning treatments on the postharvest quality of horticultural crops. Stewart Postharvest Rev. 2007, 3, 1–6. [Google Scholar]
- Lafuente, M.T.; Zacarias, L. Postharvest physiological disorders in citrus fruit. Stewart Postharvest Rev. 2006, 2, 1–9. [Google Scholar]
- Lafuente, M.T.; Zacarias, L.; Sala, J.M.; Lluch, Y.; Granell, A.; Sánchez-Ballesta, M.T.; Gosalbes, M.J.; Marcos, J.F.; González-Candelas, L. Understanding the basis of chilling injury in citrus fruit. Acta Hortic. 2005, 682, 831–842. [Google Scholar] [CrossRef]
- Lurie, S.; Laamim, M.; Lapsker, Z.; Fallik, E. Heat treatments to decrease chilling injury in tomato fruit. Effects on lipids, pericarp lesions and fungal growth. Physiol. Plant. 1997, 100, 297–302. [Google Scholar] [CrossRef]
- Almeida, D.P.F.; Huber, D.J. In vivo pectin solubility in ripening and chill-injured tomato fruit. Plant Sci. 2008, 174, 174–182. [Google Scholar] [CrossRef]
- Manganaris, G.A.; Vicente, A.R.; Crisosto, C.H.; Labavitch, J.M. Cell wall modifications in chilling-injured plum fruit (Prunus salicina). Postharvest Biol. Technol. 2008, 48, 77–83. [Google Scholar] [CrossRef]
- Lurie, S.; Crisosto, C.H. Chilling injury in peach and nectarine. Postharvest Biol. Technol. 2005, 37, 195–208. [Google Scholar] [CrossRef]
- Brummell, D.A.; Dal Cin, V.; Lurie, S.; Crisosto, C.H.; Labavitch, J.M. Cell wall metabolism during the development of chilling injury in cold-stored peach fruit: Association of mealiness with arrested disassembly of cell wall pectins. J. Exp. Bot. 2004, 55, 2041–2052. [Google Scholar] [CrossRef] [Green Version]
- Cai, C.; Li, X.; Chen, K.S. Acetylsalicylic acid alleviates chilling injury of postharvest loquat (Eriobotrya japonica Lindl.) fruit. Eur. Food Res. Technol. 2006, 223, 533–539. [Google Scholar] [CrossRef]
- Deewatthanawong, R.; Doom, S.K. Chilling injury in mangosteen fruit. J. Horticulural Sci. Biotechnol. 2003, 78, 559–562. [Google Scholar]
- Song, L.L.; Gao, H.Y.; Chen, W.X.; Chen, H.J.; Mao, J.L.; Zhou, Y.J.; Duan, X.W.; Joyce, D.C. The role of 1-methylcyclopropene in lignification and expansin gene expression in peeled water bamboo shoot (Zizania caduciflora L.). J. Sci. Food Agric. 2011, 91, 2679–2683. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.L.; Li, Z.J.; Zhang, X.F.; Wang, R.; Yang, S.L. Expression analysis of lignin-associated genes in hard end pear (Pyrus pyrifolia Whangkeumbae) and its response to calcium chloride treatment conditions. J. Plant Growth Regul. 2014, 34, 251–262. [Google Scholar] [CrossRef]
- Vanholme, R.; Demedts, B.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin Biosynthesis and Structure. Plant Physiol. 2010, 153, 895–905. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Dixon, R.A. Transcriptional networks for lignin biosynthesis: More complex than we thought? Trends Plant Sci. 2011, 16, 227–233. [Google Scholar] [CrossRef] [Green Version]
- Suo, J.T.; Li, H.; Ban, Q.Y.; Han, Y.; Meng, K.; Jin, M.J.; Zhang, Z.K.; Rao, J.P. Characteristics of chilling injury-induced lignification in kiwifruit with different sensitivities to low temperatures. Postharvest Biol. Technol. 2018, 135, 8–18. [Google Scholar] [CrossRef]
- Nguyen, T.B.T.; Ketsa, S.; Doorn, W.G. Relationship between browning and the activities of polyphenoloxidase and phenylalanine ammonia lyase in banana peel during low temperature storage. Postharvest Biol. Technol. 2003, 30, 187–193. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, J.Y.; Jiang, Y.M.; Lu, W.J. Cloning and expression analysis of phenylalanine ammonia-lyase in relation to chilling tolerance in harvested banana fruit. Postharvest Biol. Technol. 2007, 44, 34–41. [Google Scholar] [CrossRef]
- Jiang, Y.M.; Joyce, D.C.; Jiang, W.B.; Lu, W.J. Effects of chilling temperatures on ethylene binding by banana fruit. Plant Growth Regul. 2004, 43, 109–115. [Google Scholar] [CrossRef]
- Chen, J.Y.; He, L.H.; Jiang, Y.M.; Wang, Y.; Joyce, D.C.; Ji, Z.L.; Lu, W.J. Role of phenylalanine ammonia-lyase in heat pretreatment-induced chilling tolerance in banana fruit. Physiol. Plant. 2008, 132, 318–328. [Google Scholar] [CrossRef]
- McGuire, R.G. Reporting of objective color measurements. Horstscience 1992, 27, 1254–1255. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.Y.; Li, L.; Limwachiranon, J.; Xie, J.; Luo, Z.S. Effect of UV-C on ripening of tomato fruits in response to wound. Sci. Hortic. 2016, 213, 104–109. [Google Scholar] [CrossRef]
- Bruce, R.J.; West, C.A. Elicitation of lignin biosynthesis and isoperoxidase activity by pectic fragments in suspension cultures of castor bean 1. Plant Physiol. 1989, 91, 889–897. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y. Role of anthocyanins, polyphenol oxidase and phenols in lychee pericarp browning. J. Sci. Food Agric. 2000, 80, 305–310. [Google Scholar] [CrossRef]
- Assis, J.S.; Muñoz, T.; Escribano, M.I.; Merodio, C. Effect of high carbon dioxide concentration on PAL activity and phenolic contents in ripening cherimoya fruit. Postharvest Biol. Technol. 2001, 23, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Zhong, H.Y.; Kuang, J.F.; Li, J.G.; Lu, W.J.; Chen, J.Y. Validation of reference genes for RT-qPCR studies of gene expression in banana fruit under different experimental conditions. Planta 2011, 234, 377–390. [Google Scholar] [CrossRef]
- Kasamalla, K.K.; Bhagwan, A.; Kumar, A.K. Effect of antioxidants on chilling injury and storage life of guava cv. Allahabad safeda. Int. J. Chem. Stud. 2017, 5, 777–783. [Google Scholar]
- Sayyari, M.; Babalar, M.; Kalantari, S.; Serrano, M.; Valero, D. Effect of salicylic acid treatment on reducing chilling injury in stored pomegranates. Postharvest Biol. Technol. 2009, 53, 152–154. [Google Scholar] [CrossRef]
- Lyons, J.M. Chilling injury in plants. Annu. Rev. Plant Biol. 1973, 24, 445–466. [Google Scholar] [CrossRef]
- Sharom, M.; Willemot, C.; Thompson, J.E. Chilling injury induces lipid phase changes in membranes of tomato fruit. Plant Physiol. 1994, 105, 305–308. [Google Scholar] [CrossRef] [Green Version]
- Li, H.X.; Xiao, Y.; Cao, L.L.; Yan, X.; Li, C.; Shi, H.Y.; Wang, J.W.; Ye, Y.H. Cerebroside C increases tolerance to chilling injury and alters lipid composition in wheat roots. PLoS ONE 2013, 8, e73380. [Google Scholar] [CrossRef] [Green Version]
- Parkin, K.L.; Marangon, A.; Jackman, R.L.; Yada, R.Y.; Stanley, D.W. Chilling injury. A review of possible mechanism. J. Food Biochem. 1988, 13, 127–153. [Google Scholar] [CrossRef]
- Scandalio, J.G. Oxygen stress and superoxide dismutases. Plant Physiol. 1993, 101, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Allen, R.D. Dissection of oxidative stress tolerance using transgenic plants. Plant Physiol. 1995, 107, 1049–1054. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, K.; Kanematsu, S.; Asada, K. Generation of superoxide anion and localization of CuZn-superoxide dismutase in the vascular tissue of spinach hypocotyls: Their association with lignification. Plant Cell Physiol. 1997, 38, 1118–1126. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.S.; Wu, X.; Xie, Y.; Chen, C. Alleviation of chilling injury and browning of postharvest bamboo shoot by salicylic acid treatment. Food Chem. 2012, 131, 456–461. [Google Scholar] [CrossRef]
- Trakulnaleumsai, C.; Ketsa, S.; van Doorn, W.G. Temperature effects on peel spotting in ‘Sucrier’ banana fruit. Postharvest Biol. Technol. 2006, 39, 285–290. [Google Scholar] [CrossRef]
- Martinez, M.V.; Whitaker, J.R. The biochemistry and control of enzymatic browning. Trends Food Sci. Technol. 1995, 6, 195–200. [Google Scholar] [CrossRef]
- Subramanian, N.; Venkatesh, P.; Ganguli, S.; Sinkar, V.P. Role of polyphenol oxidase and peroxidase in the generation of black tea theaflavins. J. Agric. Food Chem. 1999, 47, 2571–2578. [Google Scholar] [CrossRef]
- Richard-Forget, F.C.; Gauillard, F.A. Oxidation of chlorogenic acid, catechins, and 4-methylcatechol in model solutions by combinations of pear (Pyrus communis Cv. Williams) polyphenol oxidase and peroxidase: A possible involvement of peroxidase in enzymatic browning. J. Agric. Food Chem. 1997, 45, 2472–2476. [Google Scholar] [CrossRef]
- Long, G.H.; Wu, P.Y.; Fu, J.Z.; Lu, H.L.; Zhang, R. Research progress on regulation of peroxidase on lignin synthesis. Mod. Agric. Sci. Technol. 2021, 23, 47–54. [Google Scholar]
- Dixon, R.A.; Choudhary, A.D.; Dalkin, K.; Edwards, R.; Fahrendorf, T.; Gowri, G.; Harrison, M.J.; Lamb, C.J.; Loake, G.J.; Maxwell, C.A.; et al. Molecular biology of stress-induced phenylpropanoid and isoflavonoid biosynthesis in Alfalfa. Phenolic Metab. Plants 1992, 26, 91–138. [Google Scholar]
- Lafuente, M.T.; Sala, J.M.; Zacarias, L. Active oxygen detoxifying enzymes and phenylalanine ammonia-lyase in the ethylene-induced chilling tolerance in citrus Fruit. J. Agric. Food Chem. 2004, 52, 3606–3611. [Google Scholar] [CrossRef]
- Lafuente, M.T.; Zacarias, L.; Martínez-Téllez, M.A.; Sanchez-Ballesta, M.T.; Granell, A. Phenylalanine ammonia-lyase and ethylene in relation to chilling injury as affected by fruit age in citrus. Postharvest Biol. Technol. 2003, 29, 309–318. [Google Scholar] [CrossRef]
- Lafuente, M.T.; Zacarias, L.; Martínez-Téllez, M.A.; Sanchez-Ballesta, M.T.; Dupille, E. Phenylalanine ammonia-lyase as related to ethylene in the development of chilling symptoms during cold storage of citrus fruits. J. Agric. Food Chem. 2001, 49, 6020–6025. [Google Scholar] [CrossRef]
- Choehom, R.; Ketsa, S.; van Doorn, W.G. Senescent spotting of banana peel is inhibited by modified atmosphere packaging. Postharvest Biol. Technol. 2004, 31, 167–175. [Google Scholar] [CrossRef]
- Simon, C.; Lion, C.; Biot, C.; Gierlinger, N.; Hawkins, S. Lignification and Advances in Lignin Imaging in Plant Cell Walls. Annual Plant Reviews. 2018, 1, 1–32. [Google Scholar]
- Simerjeet, K.; Xu, Z.; Amita, M.; Dong, H.; Prashant, V.; Sukhwinder, S.; Zhang, Z.; Gill, K.S.; Dhugga, K.S.; Jaswinder, S. Genome-wide association study reveals novel genes associated with culm cellulose content in bread wheat (Triticum aestivum, L.). Front. Plant Sci. 2017, 8, 1913. [Google Scholar]
- Zhang, L.F.; Qian, W.U.; Chen, F.S.; Yang, G.S. Research on the structure, function and methods of modified pectin. Cereals Oils 2015, 28, 1–5. [Google Scholar]
- Huang, H.; Wang, S.F.; Han, Y.M. Research progress on the mechanism of cell wall influence on fruit texture. Sci. Technol. Food Ind. 2019, 40, 350–355. [Google Scholar]
- Zheng, H.Z.; Cui, C.L.; Zhang, Y.T.; Wang, D.; Jing, Y.; Kim, K.Y. Active changes of lignification-related enzymes in pepper response to Glomus intraradices and/or Phytophthora capsici. J. Zhejiang Univ. Sci. B 2005, 6, 778–786. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.Y.; Liu, Z.H.; Zhu, L.; Zhang, C.J.; Chen, Y.; Zhou, Y.; Li, F.G.; Li, X.B. Overexpression of cotton (Gossypium hirsutum) dirigent1 gene enhances lignification that blocks the spread of Verticillium dahliae. Acta Biochim. Biophys Sin. 2012, 44, 555–564. [Google Scholar] [CrossRef] [Green Version]
- Benske, A. Laccase-dependent lignification of secondary cell walls of protoxylem tracheary elements in Arabidopsis thaliana. Master’s Thesis, University of British Columbia, Vancouver, BC, Canada, 2014. [Google Scholar] [CrossRef]
- Wang, Y.; Bouchabke-Coussa, O.; Lebris, P.; Antelme, S.; Soulhat, C. LACCASE5 is required for lignification of the Brachypodium distachyon Culm. Plant Physiol. 2015, 168, 192–204. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.Y.; Zhang, S.C.; Yang, Y.; Luo, Y.C.; Liu, Q.; Zhang, Y.C.; Qu, L.H.; Lucas, W.; Wang, X.J.; Chen, Y.Q. MiR397b regulates both lignin content and seed number in Arabidopsis via modulating a laccase involved in lignin biosynthesis. Plant Biotechnol. J. 2014, 12, 1132–1142. [Google Scholar] [CrossRef]
- Sun, Q.; Liu, X.; Yang, J.; Liu, W.; Du, Q.; Wang, H.; Fu, C.; Li, W.X. microRNA528 Affects lodging resistance of Maize by regulating lignin biosynthesis under Nitrogen-Luxury conditions. Mol. Plant 2018, 11, 806–814. [Google Scholar] [CrossRef] [Green Version]
- Yin, N.; Li, B.; Liu, X.; Liang, Y.; Lian, J.; Xue, Y.; Qu, C.; Lu, K.; Wei, L.; Wang, R. Cinnamoyl-CoA Reductase 1 (CCR1) and CCR2 Function Divergently in Tissue Lignification, Flux Control and Cross-Talk with Glucosinolate Pathway in Brassica napus; Cold Spring Harbor Laboratory: Long Island, NY, USA, 2021. [Google Scholar] [CrossRef]
- Mansell, R.L.; Gross, G.G.; Stöckigt, J.; Franke, H.; Zenk, M.H. Purification and properties of cinnamyl alcohol dehydrogenase from higher plants involved in lignin biosynthesis. Phytochemistry 1974, 13, 2427–2435. [Google Scholar] [CrossRef]
- Brill, E.; Abrahams, S.; Hayes, C.M.; Jenkins, C.L.D.; Watson, J.M. Molecular characterisation and expression of a wound-inducible cDNA encoding a novel cinnamyl-alcohol dehydrogenase enzyme in lucerne (Medicago sativa L.). Plant Mol. Biol. 1999, 41, 279–291. [Google Scholar] [CrossRef]
- Lynch, D.; Lidgett, A.; Mcinnes, R.; Huxley, H.; Jones, E.; Mahoney, N.; Spangenberg, G. Isolation and characterisation of three cinnamyl alcohol dehydrogenase homologue cDNAs from perennial ryegrass (Lolium perenne L.). J. Plant Physiol. 2002, 159, 653–660. [Google Scholar] [CrossRef]
- Tsuruta, S.; Ebina, M.; Nakagawa, H.; Kawamura, O.; Akashi, R. Isolation and characterization of cDNA encoding cinnamyl alcohol dehydrogenase (CAD) in sorghum (Sorghum bicolor (L.) Moench). Grassl. Sci. 2007, 53, 103–109. [Google Scholar] [CrossRef]
- Allina, S.M.; Pri-Hadash, A.; Theilmann, D.A.; Ellis, B.E.; Douglas, C.J. 4-Coumarate: Coenzyme a ligase in hybrid poplar. Plant Physiol. 1998, 116, 743–754. [Google Scholar] [CrossRef] [Green Version]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, L.; Jiang, X.; Deng, Y.; Xu, K.; Duan, X.; Wan, K.; Tang, X. Study on Characteristics and Lignification Mechanism of Postharvest Banana Fruit during Chilling Injury. Foods 2023, 12, 1097. https://doi.org/10.3390/foods12051097
Xiao L, Jiang X, Deng Y, Xu K, Duan X, Wan K, Tang X. Study on Characteristics and Lignification Mechanism of Postharvest Banana Fruit during Chilling Injury. Foods. 2023; 12(5):1097. https://doi.org/10.3390/foods12051097
Chicago/Turabian StyleXiao, Lu, Xunyuan Jiang, Yicai Deng, Kaihang Xu, Xuewu Duan, Kai Wan, and Xuemei Tang. 2023. "Study on Characteristics and Lignification Mechanism of Postharvest Banana Fruit during Chilling Injury" Foods 12, no. 5: 1097. https://doi.org/10.3390/foods12051097