
Metabolic Profile of Einkorn, Spelt, Emmer Ancient Wheat Species Sourdough Fermented with Strain of Lactiplantibacillus plantarum ATCC 8014

 ,
,  , ,
, ,  ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Sourdough Formulation, Lactiplantibacillusplantarum ATCC 8014 Activation, and Cell Count Determination
2.3. Sourdough Acidification—Total Titratable Acidity (TTA) and pH Determination
2.4. Determination of Carbohydrates, Organic Acids, and Ethanol Content by HPLC-RID
2.5. Determination of Crude Fiber Content (Crude Cellulose)
2.6. Determination of Micro and Macroelements by Atomic Absorption Spectrophotometry (AAS)
2.7. Determination of Volatile Compounds by ITEX/GC-MS Technique
2.8. Determination of Rheological Properties
2.9. Statistical Analyses
3. Results and Discussion
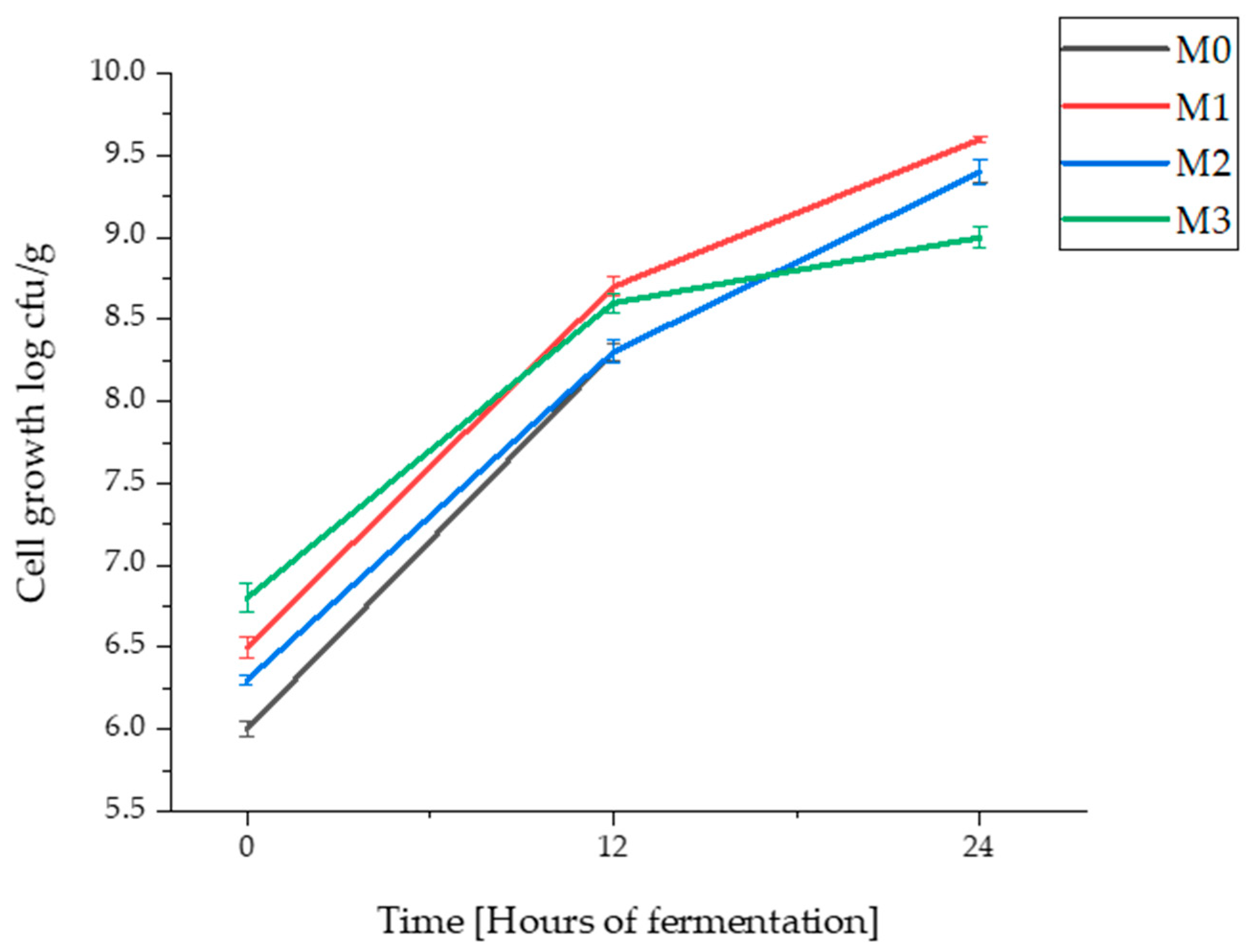
3.1. Cell Viability in Sourdough Samples
3.2. pH and TTA Values
3.3. Carbohydrates and Organic Acids Content
3.4. Crude Cellulose Content
3.5. Minerals Content
3.6. Volatile Compounds Content
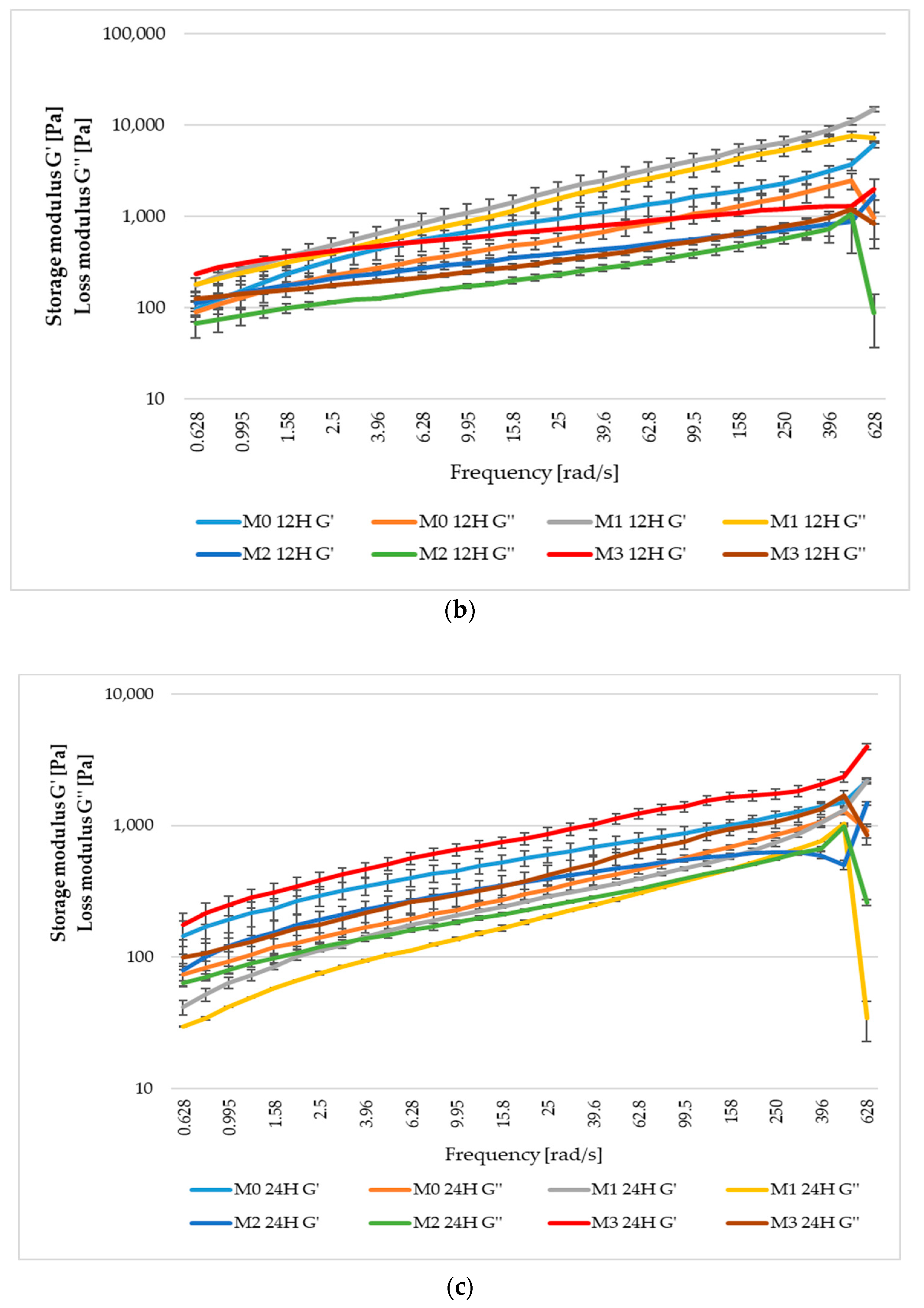
3.7. Rheological Values
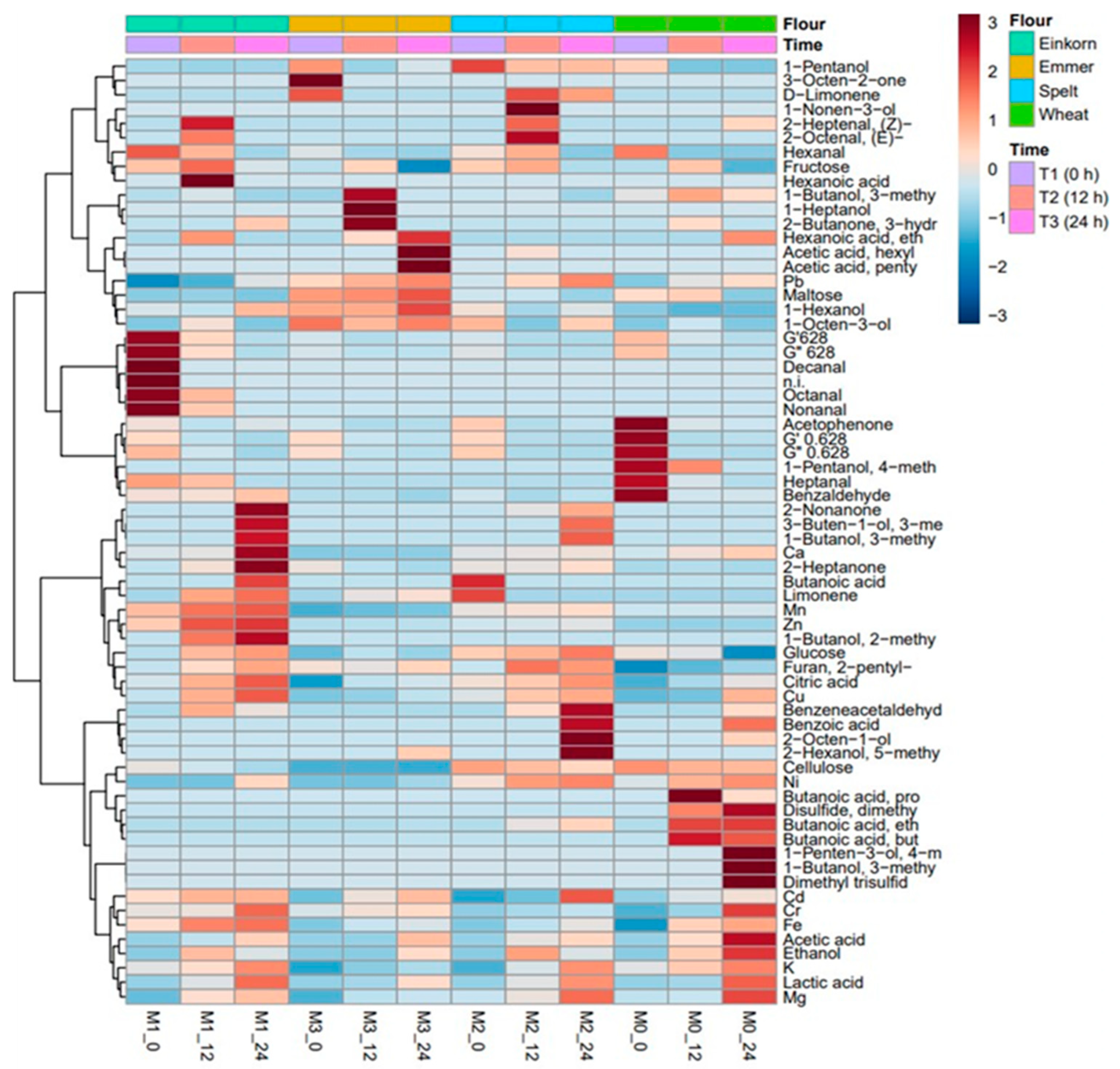
3.8. Effect of Sourdough Type on Metabolic Profile
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Vuyst, L.; Comasio, A.; Kerrebroeck, S. Van Sourdough production: Fermentation strategies, microbial ecology, and use of non-flour ingredients. Crit. Rev. Food Sci. Nutr. 2021, 1–33. [Google Scholar] [CrossRef] [PubMed]
- Arora, K.; Ameur, H.; Polo, A.; Di Cagno, R.; Rizzello, C.G.; Gobbetti, M. Thirty years of knowledge on sourdough fermentation: A systematic review. Trends Food Sci. Technol. 2021, 108, 71–83. [Google Scholar] [CrossRef]
- Cappelle, S.; Guylaine, L.; Gänzle, M.; Gobbetti, M. History and social aspects of sourdough. In Handbook on Sourdough Biotechnology; Gobbetti, M., Gänzle, M., Eds.; Springer: New York, NY, USA, 2013; pp. 1–10. [Google Scholar]
- Blaiotta, G.; Romano, R.; Trifuoggi, M.; Aponte, M.; Miro, A. Development of a Wet-Granulated Sourdough Multiple Starter for Direct Use. Foods 2022, 11, 1278. [Google Scholar] [CrossRef] [PubMed]
- Capozzi, V.; Fragasso, M.; Romaniello, R.; Berbegal, C.; Russo, P.; Spano, G. Spontaneous food fermentations and potential risks for human health. Fermentation 2017, 3, 49. [Google Scholar] [CrossRef]
- Corsetti, A.; Perpetuini, G.; Schirone, M.; Tofalo, R.; Suzzi, G. Application of starter cultures to table olive fermentation: An overview on the experimental studies. Front. Microbiol. 2012, 3, 248. [Google Scholar] [CrossRef] [Green Version]
- Siepmann, F.B.; Sousa de Almeida, B.; Waszczynskyj, N.; Spier, M.R. Influence of temperature and of starter culture on biochemical characteristics and the aromatic compounds evolution on type II sourdough and wheat bread. LWT-Food Sci. Technol. 2019, 108, 199–206. [Google Scholar] [CrossRef]
- Capurso, A.; Capurso, C. The Mediterranean way: Why elderly people should eat wholewheat sourdough bread—A little known component of the Mediterranean diet and healthy food for elderly adults. Aging Clin. Exp. Res. 2020, 32, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Calvert, M.D.; Madden, A.A.; Nichols, L.M.; Haddad, N.M.; Lahne, J.; Dunn, R.R.; McKenney, E.A. A review of sourdough starters: Ecology, practices, and sensory quality with applications for baking and recommendations for future research. PeerJ 2021, 9, e11389. [Google Scholar] [CrossRef]
- Codină, G.G.; Sarion, C.; Dabija, A. Effects of dry sourdough on bread-making quality and acrylamide content. Agronomy 2021, 11, 1977. [Google Scholar] [CrossRef]
- Gil-Cardoso, K.; Saldaña, G.; Luengo, E.; Pastor, J.; Virto, R.; Alcaide-Hidalgo, J.M.; Del Bas, J.M.; Arola, L.; Caimari, A. Consumption of Sourdough Breads Improves Postprandial Glucose Response and Produces Sourdough-Specific Effects on Biochemical and Inflammatory Parameters and Mineral Absorption. J. Agric. Food Chem. 2021, 69, 3044–3059. [Google Scholar] [CrossRef]
- Rašević, V.; Vranac, A.; Žuljević, S.O. Impact of sourdough addition on the bread quality. In Proceedings of the Radovi Poljoprivrednog Fakulteta Univerziteta u Sarajevu\Works of the Faculty of Agriculture University of Sarajevo, Sarajevo, Bosnia, 27–29 September 2017; Volume 62, pp. 401–410. [Google Scholar]
- Menezes, L.A.A.; Minervini, F.; Filannino, P.; Sardaro, M.L.S.; Gatti, M.; Lindner, J.D.D. Effects of sourdough on FODMAPs in bread and potential outcomes on irritable bowel syndrome patients and healthy subjects. Front. Microbiol. 2018, 9, 1972. [Google Scholar] [CrossRef] [Green Version]
- Gobbetti, M.; Rizzello, C.G.; Di Cagno, R.; De Angelis, M. How the sourdough may affect the functional features of leavened baked goods. Food Microbiol. 2014, 37, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Park, D.M.; Bae, J.H.; Kim, M.S.; Kim, H.; Kang, S.D.; Shim, S.; Lee, D.; Seo, J.H.; Kang, H.; Han, N.S. Suitability of Lactobacillus plantarum SPC-SNU 72-2 as a Probiotic Starter for Sourdough Fermentation. J. Microbiol. Biotechnol. 2019, 29, 1729–1738. [Google Scholar] [CrossRef] [PubMed]
- Zaidiyah; Lubis, Y.M.; Putri, C.A.R.G.; Rohaya, S. Physicochemical properties of sourdough bread made from local variety sweet potato and pineapple juice. IOP Conf. Ser. Earth Environ. Sci. 2020, 425, 012079. [Google Scholar] [CrossRef]
- Quattrini, M.; Liang, N.; Fortina, M.G.; Xiang, S.; Curtis, J.M.; Gänzle, M. Exploiting synergies of sourdough and antifungal organic acids to delay fungal spoilage of bread. Int. J. Food Microbiol. 2019, 302, 8–14. [Google Scholar] [CrossRef]
- Arendt, E.K.; Ryan, L.A.M.; Dal Bello, F. Impact of sourdough on the texture of bread. Food Microbiol. 2007, 24, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Rizzello, C.G.; Portincasa, P.; Montemurro, M.; di Palo, D.M.; Lorusso, M.P.; de Angelis, M.; Bonfrate, L.; Genot, B.; Gobbetti, M. Sourdough fermented breads are more digestible than those started with baker’s yeast alone: An in vivo challenge dissecting distinct gastrointestinal responses. Nutrients 2019, 11, 2954. [Google Scholar] [CrossRef] [Green Version]
- Șerban, L.R.; Păucean, A.; Man, S.M.; Chiş, M.S.; Mureşan, V. Ancient wheat species: Biochemical profile and impact on sourdough bread characteristics—A review. Processes 2021, 9, 8. [Google Scholar] [CrossRef]
- Woo, S.H.; Shin, Y.J.; Jeong, H.M.; Kim, J.S.; Ko, D.S.; Hong, J.S.; Choi, H.D.; Shim, J.H. Effects of maltogenic amylase from Lactobacillus plantarum on retrogradation of bread. J. Cereal Sci. 2020, 93, 102976. [Google Scholar] [CrossRef]
- Lavermicocca, P.; Reguant, C.; Bautista-Gallego, J. Editorial: Lactic Acid Bacteria Within the Food Industry: What Is New on Their Technological and Functional Role. Front. Microbiol. 2021, 12, 711013. [Google Scholar] [CrossRef]
- Zhang, L.; Taal, M.; Boom, R.M.; Chen, X.D.; Schutyser, M.A.I. Viability of Lactobacillus plantarum P8 in Bread during Baking and Storage. In Proceedings of the The 20th International Drying Symposium, Giju, Japan, 7–10 August 2015. [Google Scholar]
- Chiș, S.M.; Păucean, A.; Stan, L.; Mureșan, V. Lactobacillus plantarum ATCC 8014 in quinoa sourdough adaptability and antioxidant potential. Rom. Biotechnol. Lett. 2018, 23, 13581–13591. [Google Scholar]
- da Silva Sabo, S.; Vitolo, M.; González, J.M.D.; Oliveira, R.P.d.S. Overview of Lactobacillus plantarum as a promising bacteriocin producer among lactic acid bacteria. Food Res. Int. 2014, 64, 527–536. [Google Scholar] [CrossRef]
- Di Renzo, T.; Reale, A.; Boscaino, F.; Messia, M.C. Flavoring production in Kamut®, Quinoa and wheat doughs fermented by Lactobacillus paracasei, Lactobacillus plantarum, and Lactobacillus brevis: A SPME-GC/MS study. Front. Microbiol. 2018, 9, 429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohsen, S.M.; Aly, M.H.; Attia, A.A.; Osman, D.B. Effect of Sourdough on Shelf Life, Freshness and Sensory Characteristics of Egyptian Balady Bread. J. Appl. Environ. Microbiol. 2016, 4, 39–45. [Google Scholar] [CrossRef]
- Coda, R.; Nionelli, L.; Rizzello, C.G.; De Angelis, M.; Tossut, P.; Gobbetti, M. Spelt and emmer flours: Characterization of the lactic acid bacteria microbiota and selection of mixed starters for bread making. J. Appl. Microbiol. 2010, 108, 925–935. [Google Scholar] [CrossRef]
- Păucean, A.; Vodnar, D.C.; Socaci, S.A.; Socaciu, C. Carbohydrate metabolic conversions to lactic acid and volatile derivatives, as influenced by Lactobacillus plantarum ATCC 8014 and Lactobacillus casei ATCC 393 efficiency during in vitro and sourdough fermentation. Eur. Food Res. Technol. 2013, 237, 679–689. [Google Scholar] [CrossRef]
- Bartkiene, E.; Bartkevics, V.; Pugajeva, I.; Krungleviciute, V.; Mayrhofer, S.; Domig, K. Parameters of rye, wheat, barley, and oat sourdoughs fermented with Lactobacillus plantarum LUHS135 that influence the quality of mixed rye–wheat bread, including acrylamide formation. Int. J. Food Sci. Technol. 2017, 52, 1473–1482. [Google Scholar] [CrossRef]
- Čurná, V.; Lacko-Bartošová, M. Chemical composition and nutritional value of emmer wheat (Triticum dicoccon schrank): A review. J. Cent. Eur. Agric. 2017, 18, 117–134. [Google Scholar] [CrossRef] [Green Version]
- Bencze, S.; Makádi, M.; Aranyos, T.J.; Földi, M.; Hertelendy, P.; Mikó, P.; Bosi, S.; Negri, L.; Drexler, D. Re-introduction of ancient wheat cultivars into organic agriculture-Emmer and Einkorn cultivation experiences under marginal conditions. Sustainability 2020, 12, 1584. [Google Scholar] [CrossRef] [Green Version]
- Van Boxstael, F.; Aerts, H.; Linssen, S.; Latré, J.; Christiaens, A.; Haesaert, G.; Dierickx, I.; Brusselle, J.; De Keyzer, W. A comparison of the nutritional value of Einkorn, Emmer, Khorasan and modern wheat: Whole grains, processed in bread, and population-level intake implications. J. Sci. Food Agric. 2020, 100, 4108–4118. [Google Scholar] [CrossRef]
- Hidalgo, A.; Brandolini, A.; Pompei, C.; Piscozzi, R. Carotenoids and tocols of einkorn wheat (Triticum monococcum ssp. monococcum L.). J. Cereal Sci. 2006, 44, 182–193. [Google Scholar] [CrossRef]
- Costanzo, A.; Amos, D.C.; Dinelli, G.; Sferrazza, R.E.; Accorsi, G.; Negri, L.; Bosi, S. Performance and nutritional properties of Einkorn, Emmer and Rivet Wheat in response to different rotational position and soil tillage. Sustainability 2019, 11, 6304. [Google Scholar] [CrossRef] [Green Version]
- Geisslitz, S.; Scherf, K.A. Rediscovering Ancient Wheats. Cereal Foods World 2020, 65, 13. [Google Scholar] [CrossRef]
- Longin, C.F.H.; Ziegler, J.; Schweiggert, R.; Koehler, P.; Carle, R.; Würschum, T. Comparative study of hulled (einkorn, emmer, and spelt) and naked wheats (durum and bread wheat): Agronomic performance and quality traits. Crop Sci. 2016, 56, 302–311. [Google Scholar] [CrossRef]
- Escarnot, E.; Jacquemin, J.M.; Agneessens, R.; Paquot, M. Comparative study of the content and profiles of macronutrients in spelt and wheat, a review. Biotechnol. Agron. Société Environ. 2012, 16, 243–256. [Google Scholar]
- Şahin, Y.; Yıldırım, A.; Yücesan, B.; Zencirci, N.; Erbayram, Ş.; Gürel, E. Phytochemical content and antioxidant activity of einkorn (Triticum monococcum ssp. monococcum), bread (Triticum aestivum L.), and durum (Triticum durum Desf.) wheat. Nutrition 2017, 19, 450–459. [Google Scholar] [CrossRef]
- Hidalgo, A.; Brandolini, A. Nutritional properties of einkorn wheat (Triticum monococcum L.). J. Sci. Food Agric. 2014, 94, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Zencirci, N.; Karakas, F.P.; Ordu, B. Macro-Micro Element Variation in Traditionally Grown Einkorn (Triticum monococcum L. subsp. monococcum) and Emmer Wheat (Triticum dicoccon Schrank). Int. J. Second. Metab. 2021, 8, 227–245. [Google Scholar] [CrossRef]
- Frakolaki, G.; Giannou, V.; Topakas, E.; Tzia, C. Chemical characterization and breadmaking potential of spelt versus wheat flour. J. Cereal Sci. 2018, 79, 50–56. [Google Scholar] [CrossRef]
- Kohajdová, Z.; Karovičová, J. Nutritional Value and Baking Applications of Spelt Wheat *. ACTA Acta Sci. Pol. Technol. Aliment 2008, 7, 5–14. [Google Scholar]
- Dinu, M.; Whittaker, A.; Pagliai, G.; Benedettelli, S.; Sofi, F. Ancient wheat species and human health: Biochemical and clinical implications. J. Nutr. Biochem. 2018, 52, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Thorup, A.C.; Gregersen, S.; Jeppesen, P.B. Ancient Wheat Diet Delays Diabetes Development in a Type 2 Diabetes Animal Model. Rev. Diabet. Stud. 2014, 11, 245–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shewry, P.R.; Hey, S. Do “ancient” wheat species differ from modern bread wheat in their contents of bioactive components? J. Cereal Sci. 2015, 65, 236–243. [Google Scholar] [CrossRef] [Green Version]
- Guerrini, L.; Parenti, O.; Angeloni, G.; Zanoni, B. The bread making process of ancient wheat: A semi-structured interview to bakers. J. Cereal Sci. 2019, 87, 9–17. [Google Scholar] [CrossRef]
- Sereti, V.; Lazaridou, A.; Biliaderis, C.G.; Valamoti, S.M. Reinvigorating modern breadmaking based on ancient practices and plant ingredients, with implementation of a physicochemical approach. Foods 2021, 10, 789. [Google Scholar] [CrossRef]
- Păucean, A.; Mureșan, V.; Maria-Man, S.; Chiș, M.S.; Mureșan, A.E.; Șerban, L.R.; Pop, A.; Muste, S. Metabolomics as a tool to elucidate the sensory, nutritional and safety quality of wheat bread—A review. Int. J. Mol. Sci. 2021, 22, 8945. [Google Scholar] [CrossRef]
- Cevallos-Cevallos, J.M.; Reyes-De-Corcuera, J.I.; Etxeberria, E.; Danyluk, M.D.; Rodrick, G.E. Metabolomic analysis in food science: A review. Trends Food Sci. Technol. 2009, 20, 557–566. [Google Scholar] [CrossRef]
- Weckx, S.; Van Kerrebroeck, S.; De Vuyst, L. Omics approaches to understand sourdough fermentation processes. Int. J. Food Microbiol. 2019, 302, 90–102. [Google Scholar] [CrossRef]
- Saa, D.L.T.; Nissen, L.; Gianotti, A. Metabolomic approach to study the impact of flour type and fermentation process on volatile profile of bakery products. Food Res. Int. 2019, 119, 510–516. [Google Scholar] [CrossRef]
- Colosimo, R.; Gabriele, M.; Cifelli, M.; Longo, V.; Domenici, V.; Pucci, L. The effect of sourdough fermentation on Triticum dicoccum from Garfagnana: 1H NMR characterization and analysis of the antioxidant activity. Food Chem. 2020, 305, 125510. [Google Scholar] [CrossRef]
- Righetti, L.; Rubert, J.; Galaverna, G.; Folloni, S.; Ranieri, R.; Stranska-Zachariasova, M.; Hajslov, J.; Dall’Asta, C. Characterization and discrimination of ancient grains: A metabolomics approach. Int. J. Mol. Sci. 2016, 17, 1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balestra, F.; Laghi, L.; Taneyo Saa, D.; Gianotti, A.; Rocculi, P.; Pinnavaia, G.G. Physico-chemical and metabolomic characterization of KAMUT® Khorasan and durum wheat fermented dough. Food Chem. 2015, 187, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Ferri, M.; Serrazanetti, D.I.; Tassoni, A.; Baldissarri, M.; Gianotti, A. Improving the functional and sensorial profile of cereal-based fermented foods by selecting Lactobacillus plantarum strains via a metabolomics approach. Food Res. Int. 2016, 89, 1095–1105. [Google Scholar] [CrossRef]
- Shewry, P.R.; America, A.H.P.; Lovegrove, A.; Wood, A.J.; Plummer, A.; Evans, J.; van den Broeck, H.C.; Gilissen, L.; Mumm, R.; Ward, J.L.; et al. Comparative compositions of metabolites and dietary fibre components in doughs and breads produced from bread wheat, emmer and spelt and using yeast and sourdough processes. Food Chem. 2022, 374, 131710. [Google Scholar] [CrossRef] [PubMed]
- Chiş, M.S.; Păucean, A.; Man, S.M.; Vodnar, D.C.; Teleky, B.E.; Pop, C.R.; Stan, L.; Borsai, O.; Kadar, C.B.; Urcan, A.C.; et al. Quinoa sourdough fermented with Lactobacillus plantarum ATCC 8014 designed for gluten-free muffins—A powerful tool to enhance bioactive compounds. Appl. Sci. 2020, 10, 7140. [Google Scholar] [CrossRef]
- Coda, R.; Di Cagno, R.; Rizzello, C.G.; Nionelli, L.; Edema, M.O.; Gobbetti, M. Utilization of African Grains for Sourdough Bread Making. J. Food Sci. 2011, 76, 329–335. [Google Scholar] [CrossRef]
- Pontonio, E.; Arora, K.; Dingeo, C.; Carafa, I.; Celano, G.; Scarpino, V.; Genot, B.; Gobbetti, M.; Di Cagno, R. Commercial Organic Versus Conventional Whole Rye and Wheat Flours for Making Sourdough Bread: Safety, Nutritional, and Sensory Implications. Front. Microbiol. 2021, 12, 674413. [Google Scholar] [CrossRef]
- Hotea, I.; Colibar, O.; Sîrbu, C.; Berbecea, A.; Radulov, I. Considerations on the fiber and protein content interinfluence in oats used as feedstuff in animal diets. Lucr. Științifice-Univ. Științe Agric. a Banat. Timișoara, Med. Vet. 2020, 53, 42–49. [Google Scholar]
- Păucean, A.; Moldovan, O.P.; Mureșan, V.; Socaci, S.A.; Dulf, F.V.; Alexa, E.; Man, S.M.; Mureșan, A.E.; Muste, S. Folic acid, minerals, amino-acids, fatty acids and volatile compounds of green and red lentils. Folic acid content optimization in wheat-lentils composite flours. Chem. Cent. J. 2018, 12, 88. [Google Scholar] [CrossRef] [Green Version]
- Salanţă, L.-C.; Tofană, M.; Socaci, S.A.; Lazar, C.; Michiu, D.; Fărcaş, A. Determination of the Volatile Compounds from Hop and Hop Products using ITEX/GC-MS Technique. J. Agroaliment. Process. Technol. 2012, 18, 110–115. [Google Scholar]
- Socaci, S.A.; Socaciu, C.; Tofanǎ, M.; Raţi, I.V.; Pintea, A. In-tube extraction and GC-MS analysis of volatile components from wild and cultivated sea buckthorn (Hippophae rhamnoides L. ssp. Carpatica) berry varieties and juice. Phytochem. Anal. 2013, 24, 319–328. [Google Scholar] [CrossRef]
- Chiș, M.S.; Păucean, A.; Stan, L.; Suharoschi, R.; Socaci, S.A.; Man, S.M.; Pop, C.R.; Muste, S. Impact of protein metabolic conversion and volatile derivatives on gluten-free muffins made with quinoa sourdough. CYTA-J. Food 2019, 17, 744–753. [Google Scholar] [CrossRef]
- Teleky, B.E.; Martău, A.G.; Ranga, F.; Chețan, F.; Vodnar, D.C. Exploitation of lactic acid bacteria and Baker’s yeast as single or multiple starter cultures of wheat flour dough enriched with soy flour. Biomolecules 2020, 10, 778. [Google Scholar] [CrossRef] [PubMed]
- Tanislav, A.E.; Pușcaș, A.; Păucean, A.; Mureșan, A.E.; Semeniuc, C.A.; Mureșan, V.; Mudura, E. Evaluation of Structural Behavior in the Process Dynamics of Oleogel-Based Tender Dough Products. Gels 2022, 8, 317. [Google Scholar] [CrossRef] [PubMed]
- Çakır, E.; Arıcı, M.; Durak, M.Z.; Karasu, S. The molecular and technological characterization of lactic acid bacteria in einkorn sourdough: Effect on bread quality. J. Food Meas. Charact. 2020, 14, 1646–1655. [Google Scholar] [CrossRef]
- Śliżewska, K.; Chlebicz-Wójcik, A. Growth kinetics of probiotic Lactobacillus strains in the alternative, cost-efficient semi-solid fermentation medium. Biology 2020, 9, 423. [Google Scholar] [CrossRef] [PubMed]
- Coda, R.; Di Cagno, R.; Gobbetti, M.; Rizzello, C.G. Sourdough lactic acid bacteria: Exploration of non-wheat cereal-based fermentation. Food Microbiol. 2014, 37, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Pehlivan Karakas, F.; Keskin, C.N.; Agil, F.; Zencirci, N. Profiles of vitamin B and E in wheat grass and grain of einkorn (Triticum monococcum spp. monococcum), emmer (Triticum dicoccum ssp. dicoccum Schrank.), durum (Triticum durum Desf.), and bread wheat (Triticum aestivum L.) cultivars by LC-ESI-MS/MS analysi. J. Cereal Sci. 2021, 98, 103177. [Google Scholar] [CrossRef]
- Duliński, R.; Starzyńska-Janiszewska, A. Content and in vitro bioavailability of selected b vitamins and myo-inositol in spelt wheat (Triticum spelta L.) subjected to solid-state fermentation. J. Food Nutr. Res. 2020, 59, 1–6. [Google Scholar]
- Korcari, D.; Ricci, G.; Quattrini, M.; Fortina, M.G. Microbial consortia involved in fermented spelt sourdoughs: Dynamics and characterization of yeasts and lactic acid bacteria. Lett. Appl. Microbiol. 2020, 70, 48–54. [Google Scholar] [CrossRef]
- Clément, H.; Prost, C.; Chiron, H.; Ducasse, M.B.; Della Valle, G.; Courcoux, P.; Onno, B. The effect of organic wheat flour by-products on sourdough performances assessed by a multi-criteria approach. Food Res. Int. 2018, 106, 974–981. [Google Scholar] [CrossRef] [PubMed]
- Hamad, S.H. Factors Affecting the Growth of Microorganisms in Food. In Progress in Food Preservation; Bhat, R., Alias, A.K., Paliyath, G., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2012; pp. 405–427. ISBN 9780470655856. [Google Scholar]
- Di Cagno, R.; Pontonio, E.; Buchin, S.; De Angelis, M.; Lattanzi, A.; Valerio, F.; Gobbetti, M.; Calasso, M. Diversity of the lactic acid bacterium and yeast microbiota in the switch from firm- to liquid-sourdough fermentation. Appl. Environ. Microbiol. 2014, 80, 3161–3172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minervini, F.; Lattanzi, A.; De Angelis, M.; Di Cagno, R.; Gobbetti, M. Influence of artisan bakery- or laboratory-propagated sourdoughs on the diversity of lactic acid bacterium and yeast microbiotas. Appl. Environ. Microbiol. 2012, 78, 5328–5340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casado, A.; Álvarez, A.; González, L.; Fernández, D.; Marcos, J.L.; Tornadijo, M.E. Effect of fermentation on microbiological, physicochemical and physical characteristics of sourdough and impact of its use on bread quality. Czech J. Food Sci. 2017, 35, 496–506. [Google Scholar] [CrossRef] [Green Version]
- Van Der Meulen, R.; Scheirlinck, I.; Van Schoor, A.; Huys, G.; Vancanneyt, M.; Vandamme, P.; De Vuyst, L. Population dynamics and metabolite target analysis of lactic acid bacteria during laboratory fermentations of wheat and spelt sourdoughs. Appl. Environ. Microbiol. 2007, 73, 4741–4750. [Google Scholar] [CrossRef] [Green Version]
- Pontonio, E.; Dingeo, C.; Di Cagno, R.; Blandino, M.; Gobbetti, M.; Rizzello, C.G. Brans from hull-less barley, emmer and pigmented wheat varieties: From by-products to bread nutritional improvers using selected lactic acid bacteria and xylanase. Int. J. Food Microbiol. 2020, 313, 108384. [Google Scholar] [CrossRef]
- Jekle, M.; Houben, A.; Mitzscherling, M.; Becker, T. Effects of selected lactic acid bacteria on the characteristics of amaranth sourdough. J. Sci. Food Agric. 2010, 90, 2326–2332. [Google Scholar] [CrossRef]
- Ognean, C.F. The effects of water content and weight of inoculum on the production of sourdough. J. Agroaliment. Process. Technol. 2015, 21, 351–357. [Google Scholar]
- Boreczek, J.; Litwinek, D.; Żylińska-Urban, J.; Izak, D.; Buksa, K.; Gawor, J.; Gromadka, R.; Bardowski, J.K.; Kowalczyk, M. Bacterial community dynamics in spontaneous sourdoughs made from wheat, spelt, and rye wholemeal flour. Microbiologyopen 2020, 9, e1009. [Google Scholar] [CrossRef]
- Sevgili, A.; Erkmen, O.; Koçaslan, S. Identification of lactic acid bacteria and yeasts from traditional sourdoughs and sourdough production by enrichment. Czech J. Food Sci. 2021, 39, 312–318. [Google Scholar] [CrossRef]
- Silva, J.D.R.; Rosa, G.C.; Neves, N.d.A.; Leoro, M.G.V.; Schmiele, M. Production of sourdough and gluten-free bread with brown rice and carioca and cowpea beans flours: Biochemical, nutritional and structural characteristics. Res. Soc. Dev. 2021, 10, e303101623992. [Google Scholar] [CrossRef]
- Czaja, T.; Sobota, A.; Szotak, R. Quantification of Ash and Moisture in Wheat Flour by. Foods 2020, 9, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vuyst, L.; Vrancken, G.; Ravyts, F.; Rimaux, T.; Weckx, S. Biodiversity, ecological determinants, and metabolic exploitation of sourdough microbiota. Food Microbiol. 2009, 26, 666–675. [Google Scholar] [CrossRef] [PubMed]
- Bo, S.; Seletto, M.; Choc, A.; Ponzo, V.; Lezo, A.; Demagistris, A.; Evangelista, A.; Ciccone, G.; Bertolino, M.; Cassader, M.; et al. The acute impact of the intake of four types of bread on satiety and blood concentrations of glucose, insulin, free fatty acids, triglyceride and acylated ghrelin. A randomized controlled cross-over trial. Food Res. Int. 2017, 92, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Zörb, C.; Betsche, T.; Langenkämper, G.; Zapp, J.; Seifert, M. Free sugars in spelt wholemeal and flour. J. Appl. Bot. Food Qual. 2007, 81, 172–174. [Google Scholar]
- Pozzo, L.; Alcántara, C.; Selma-Royo, M.; Garcia-Mantrana, I.; Bramanti, E.; Longo, V.; Collado, M.C.; Pucci, L. The impact of sourdough fermentation of spelt (Triticum dicoccum) from Garfagnana on gut microbiota composition and in vitro activity. J. Funct. Foods 2022, 91, 105007. [Google Scholar] [CrossRef]
- Saeed, M.; Randhawa, M.A.; Pasha, I.; Anjum Murtaza, M.; Afzal, J. Amino acids and organic acids production by single strain starter culture in sourdough fermentation. Br. J. Agric. Sci. 2014, 9, 121–127. [Google Scholar]
- Wang, Y.; Wu, J.; Lv, M.; Shao, Z.; Hungwe, M.; Wang, J.; Bai, X.; Xie, J.; Wang, Y.; Geng, W. Metabolism Characteristics of Lactic Acid Bacteria and the Expanding Applications in Food Industry. Front. Bioeng. Biotechnol. 2021, 9, 612285. [Google Scholar] [CrossRef]
- De Vuyst, L.; Van Kerrebroeck, S.; Leroy, F. Microbial Ecology and Process Technology of Sourdough Fermentation. In Advances in Applied Microbiology; Elsevier Ltd.: Amsterdam, The Netherlands, 2017; Volume 100, pp. 49–160. [Google Scholar]
- Pérez-Alvarado, O.; Zepeda-Hernández, A.; Garcia-Amezquita, L.E.; Requena, T.; Vinderola, G.; García-Cayuela, T. Role of lactic acid bacteria and yeasts in sourdough fermentation during breadmaking: Evaluation of postbiotic-like components and health benefits. Front. Microbiol. 2022, 13, 969460. [Google Scholar] [CrossRef]
- Novotni, D.; Ćurić, D.; Bituh, M.; Barić, I.C.; Škevin, D.; Čukelj, N. Glycemic index and phenolics of partially-baked frozen bread with sourdough. Int. J. Food Sci. Nutr. 2011, 62, 26–33. [Google Scholar] [CrossRef]
- Ventimiglia, G.; Alfonzo, A.; Galluzzo, P.; Corona, O.; Francesca, N.; Caracappa, S.; Moschetti, G.; Settanni, L. Codominance of Lactobacillus plantarum and obligate heterofermentative lactic acid bacteria during sourdough fermentation. Food Microbiol. 2015, 51, 57–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galli, V.; Venturi, M.; Pini, N.; Guerrini, S.; Granchi, L.; Vincenzini, M. Liquid and firm sourdough fermentation: Microbial robustness and interactions during consecutive backsloppings. LWT-Food Sci. Technol. 2019, 105, 9–15. [Google Scholar] [CrossRef]
- Lattanzi, A.; Minervini, F.; Di Cagno, R.; Diviccaro, A.; Antonielli, L.; Cardinali, G.; Cappelle, S.; De Angelis, M.; Gobbetti, M. The lactic acid bacteria and yeast microbiota of eighteen sourdoughs used for the manufacture of traditional Italian sweet leavened baked goods. Int. J. Food Microbiol. 2013, 163, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Peláez, J.; Paesani, C.; Gómez, M. Sourdough technology as a tool for the development of healthier grain-based products: An update. Agronomy 2020, 10, 1962. [Google Scholar] [CrossRef]
- Kulathunga, J.; Simsek, S. Dietary fiber variation in ancient and modern wheat species: Einkorn, emmer, spelt and hard red spring wheat. J. Cereal Sci. 2022, 104, 103420. [Google Scholar] [CrossRef]
- Saa, D.T.; Di Silvestro, R.; Dinelli, G.; Gianotti, A. Effect of sourdough fermentation and baking process severity on dietary fibre and phenolic compounds of immature wheat flour bread. LWT-Food Sci. Technol. 2017, 83, 26–32. [Google Scholar] [CrossRef]
- Fuckerer, K.; Hensel, O.; Schmitt, J.J. Rye Bread Fortified With Cellulose and Its Acceptance by Elderlies in Nursing Homes and Young Adults. J. Food Stud. 2016, 5, 1. [Google Scholar] [CrossRef]
- Lauková, M.; Kohajdová, Z.; Karovičová, J.; Kuchtová, V.; Minarovičová, L.; Tomášiková, L. Effects of cellulose fiber with different fiber length on rheological properties of wheat dough and quality of baked rolls. Food Sci. Technol. Int. 2017, 23, 490–499. [Google Scholar] [CrossRef]
- Ren, Y.; Linter, B.R.; Foster, T.J. Starch replacement in gluten free bread by cellulose and fibrillated cellulose. Food Hydrocoll. 2020, 107, 105957. [Google Scholar] [CrossRef]
- Atzler, J.J.; Sahin, A.W.; Gallagher, E.; Zannini, E.; Arendt, E.K. Investigation of different dietary-fibre-ingredients for the design of a fibre enriched bread formulation low in FODMAPs based on wheat starch and vital gluten. Eur. Food Res. Technol. 2021, 247, 1939–1957. [Google Scholar] [CrossRef]
- Correa, M.J.; Ferrero, C. A comparative study of commercial modified celluloses as bread making additives. Int. J. Food Prop. 2015, 18, 849–861. [Google Scholar] [CrossRef]
- Canja, C.M.; Măzărel, A.; Lupu, M.I.; Mărgean, A.; Pădureanu, V. Dietary fiber role and place in baking products. Bull. Transilv. Univ. Brasov 2016, 9, 91–96. [Google Scholar]
- McRae, M.P. Dietary Fiber Intake and Type 2 Diabetes Mellitus: An Umbrella Review of Meta-analyses. J. Chiropr. Med. 2018, 17, 44–53. [Google Scholar] [CrossRef]
- Nirmala Prasadi, V.P.; Joye, I.J. Dietary fibre from whole grains and their benefits on metabolic health. Nutrients 2020, 12, 3045. [Google Scholar] [CrossRef]
- Ioniță Mîndrican, C.B.; Ziani, K.; Mititelu, M.; Oprea, E.; Neacșu, S.M.; Moros, E.; Dumitrescu, D.-E.; Ros, A.C.; Drăgănescu, D.; Negrei, C. Therapeutic Benefits and Dietary Restrictions of Fiber Intake: A State of the Art Review. Nutrients 2022, 14, 2641. [Google Scholar] [CrossRef]
- Lattimer, J.M.; Haub, M.D. Effects of dietary fiber and its components on metabolic health. Nutrients 2010, 2, 1266–1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McRorie, J.W.; McKeown, N.M. Understanding the Physics of Functional Fibers in the Gastrointestinal Tract: An Evidence-Based Approach to Resolving Enduring Misconceptions about Insoluble and Soluble Fiber. J. Acad. Nutr. Diet. 2017, 117, 251–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gharibzahedi, S.M.T.; Jafari, S.M. The importance of minerals in human nutrition: Bioavailability, food fortification, processing effects and nanoencapsulation. Trends Food Sci. Technol. 2017, 62, 119–132. [Google Scholar] [CrossRef]
- Gupta, U.C.; Gupta, S.C. Sources and Deficiency Diseases of Mineral Nutrients in Human Health and Nutrition: A Review. Pedosphere 2014, 24, 13–38. [Google Scholar] [CrossRef]
- Tardy, A.L.; Pouteau, E.; Marquez, D.; Yilmaz, C.; Scholey, A. Vitamins and minerals for energy, fatigue and cognition: A narrative review of the biochemical and clinical evidence. Nutrients 2020, 12, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melse-Boonstra, A. Bioavailability of Micronutrients From Nutrient-Dense Whole Foods: Zooming in on Dairy, Vegetables, and Fruits. Front. Nutr. 2020, 7, 101. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.F.; Ellis, P.R.; Berry, S.E.; Latunde-Dada, G.O.; Sharp, P.A. Enhancing mineral bioavailability from cereals: Current strategies and future perspectives. Nutr. Bull. 2018, 43, 184–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samtiya, M.; Aluko, R.E.; Puniya, A.K.; Dhewa, T. Enhancing micronutrients bioavailability through fermentation of plant-based foods: A concise review. Fermentation 2021, 7, 63. [Google Scholar] [CrossRef]
- Fulea Fekensa, W. The Effect of Traditional Sourdough Starter Culture and Involved Microorganisms on Sensory and Nutritional Quality of Whole Wheat Bread. J. Food Nutr. Sci. 2021, 9, 178. [Google Scholar] [CrossRef]
- Bloot, A.P.M.; Kalschne, D.L.; Amaral, J.A.S.; Baraldi, I.J.; Canan, C. A Review of Phytic Acid Sources, Obtention, and Applications. Food Rev. Int. 2021, 1–20. [Google Scholar] [CrossRef]
- Kumar, A.; Sharma, E.; Marley, A.; Samaan, M.A.; Brookes, M.J. Iron deficiency anaemia: Pathophysiology, assessment, practical management. BMJ Open Gastroenterol. 2022, 9, e000759. [Google Scholar] [CrossRef]
- Suchowilska, E.; Wiwart, M.; Kandler, W.; Krska, R. A comparison of macro- and microelement concentrations in the whole grain of four Triticum species. Plant Soil Environ. 2012, 58, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Zahra, A.; Farooq, U.; Saeed, M.T.; Quddoos, M.Y.; Hameed, A.; Iftikhar, M.; Noreen, A.; Mahvish, S.; Bukhari, S.R.; Naqvi, S.N.; et al. Enhancement of sensory attributes and mineral content of Sourdough bread by means of microbial culture and yeast (Saccharomyces cerevisiae). Food Chem. Adv. 2022, 1, 100094. [Google Scholar] [CrossRef]
- Spisni, E.; Imbesi, V.; Giovanardi, E.; Petrocelli, G.; Alvisi, P.; Valerii, M.C. Differential physiological responses elicited by ancient and heritage wheat cultivars compared to modern ones. Nutrients 2019, 11, 2879. [Google Scholar] [CrossRef] [Green Version]
- Quintaes, K.D.; Diez-Garcia, R.W. The importance of minerals in the human diet. In Handbook of Mineral Elements in Food; de la Guardia, M., Garrigue, S., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2015; pp. 1–21. ISBN 9781118654316. [Google Scholar]
- Pétel, C.; Onno, B.; Prost, C. Sourdough volatile compounds and their contribution to bread: A review. Trends Food Sci. Technol. 2017, 59, 105–123. [Google Scholar] [CrossRef]
- Martin-Garcia, A.; Comas-bast, O.; Riu-Aumatell, M.; Latorre-Moratalla, M.; López-Tamames, E. Changes in the Volatile Profile of Wheat Sourdough Produced with the Addition of Cava Lees. Molecules 2022, 27, 3588. [Google Scholar] [CrossRef]
- De Luca, L.; Aiello, A.; Pizzolongo, F.; Blaiotta, G.; Aponte, M.; Romano, R. Volatile organic compounds in breads prepared with different sourdoughs. Appl. Sci. 2021, 11, 1330. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, W.; Adhikari, K.; Shi, Y.C. Determination of volatile compounds in heat-treated straight-grade flours from normal and waxy wheats. J. Cereal Sci. 2017, 75, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Chai, D.; Li, C.; Zhang, X.; Yang, J.; Liu, L.; Xu, X.; Du, M.; Wang, Y.; Chen, Y.; Dong, L. Analysis of volatile compounds from wheat flour in the heating process. Int. J. Food Eng. 2019, 15, 20190252. [Google Scholar] [CrossRef]
- Pizarro, F.; Franco, F. Volatile organic compounds at early stages of sourdough preparation via static headspace and GC/MS analysis. Curr. Res. Nutr. Food Sci. 2017, 5, 89–99. [Google Scholar] [CrossRef]
- Zhang, G.-H.; Wu, T.; Sadiq, F.A.; Yang, H.-Y.; Liu, T.J.; Ruan, H.; He, G. qing A study revealing the key aroma compounds of steamed bread made by Chinese traditional sourdough. J. Zhejiang Univ. Sci. B 2016, 17, 787–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arena, M.P.; Russo, P.; Spano, G.; Capozzi, V. From Microbial Ecology to Innovative Applications in Food Quality Improvements: The Case of Sourdough as a Model Matrix. J—Multidiscip. Sci. J. 2020, 3, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Yan, B.; Sadiq, F.A.; Cai, Y.; Fan, D.; Zhang, H.; Zhao, J.; Chen, W. Identification of key aroma compounds in type i sourdough-based chinese steamed bread: Application of untargeted metabolomics analysisp. Int. J. Mol. Sci. 2019, 20, 818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyaci Gunduz, C.P.; Agirman, B.; Gaglio, R.; Franciosi, E.; Francesca, N.; Settanni, L.; Erten, H. Evaluation of the variations in chemical and microbiological properties of the sourdoughs produced with selected lactic acid bacteria strains during fermentation. Food Chem. X 2022, 14, 100357. [Google Scholar] [CrossRef]
- The Good Scents Company Information System. Available online: http://www.thegoodscentscompany.com/ (accessed on 20 January 2023).
- Flavornet. Available online: http://www.flavornet.org/flavornet.html (accessed on 20 January 2023).
- Starr, G.; Petersen, M.A.; Jespersen, B.M.; Hansen, A.S. Variation of volatile compounds among wheat varieties and landraces. Food Chem. 2015, 174, 527–537. [Google Scholar] [CrossRef]
- Longoria, S.; Contreras, J.; Belmares, R.; Cruz, M.; Flores, M. Effect of short fermentation times with Lactobacillus paracasei in rheological, physical and chemical composition parameters in cassava dough and biscuits. Appl. Sci. 2020, 10, 1383. [Google Scholar] [CrossRef] [Green Version]
- Hadnađev, M.; Tomić, J.; Škrobot, D.; Dapčević-Hadnađev, T. Rheological behavior of emmer, spelt and khorasan flours. J. Food Process. Preserv. 2021, 46, e15873. [Google Scholar] [CrossRef]
- Olojede, A.O.; Sanni, A.I.; Banwo, K. Rheological, textural and nutritional properties of gluten-free sourdough made with functionally important lactic acid bacteria and yeast from Nigerian sorghum. LWT-Food Sci. Technol. 2020, 120, 108875. [Google Scholar] [CrossRef]
- Hardt, N.A.; Boom, R.M.; van der Goot, A.J. Wheat dough rheology at low water contents and the influence of xylanases. Food Res. Int. 2014, 66, 478–484. [Google Scholar] [CrossRef]
- Mohammed, I.; Ahmed, A.R.; Senge, B. Dynamic rheological properties of chickpea and wheat flour dough’s. J. Appl. Sci. 2011, 11, 3405–3412. [Google Scholar] [CrossRef]
- Meerts, M.; Cardinaels, R.; Oosterlinck, F.; Courtin, C.M.; Moldenaers, P. The Impact of Water Content and Mixing Time on the Linear and Non-Linear Rheology of Wheat Flour Dough. Food Biophys. 2017, 12, 151–163. [Google Scholar] [CrossRef] [Green Version]
- Kulathunga, J.; Reuhs, B.L.; Zwinger, S.; Simsek, S. Comparative study on kernel quality and chemical composition of ancient and modern wheat species: Einkorn, emmer, spelt and hard red spring wheat. Foods 2021, 10, 761. [Google Scholar] [CrossRef]
- Dhanavath, S.; Prasada Rao, U.J.S. Nutritional and Nutraceutical Properties of Triticum dicoccum Wheat and Its Health Benefits: An Overview. J. Food Sci. 2017, 82, 2243–2250. [Google Scholar] [CrossRef] [Green Version]
- Geisslitz, S.; Longin, C.F.H.; Scherf, K.A.; Koehler, P. Comparative Study on Gluten Protein Composition of Ancient (Einkorn, Emmer and Spelt) and Modern Wheat Species (Durum and Common Wheat). Foods 2019, 8, 409. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Materials | M0 | M1 | M2 | M3 |
|---|---|---|---|---|
| Wheat wholemeal four, g | 500 | - | - | - |
| Einkorn wholemeal flour, g | - | 500 | - | - |
| Spelt wholemeal flour, g | - | - | 500 | - |
| Emmer wholemeal four, g | - | - | - | 500 |
| Lactiplantibacillus plantarum ATCC 8014 suspension, mL | 100 | 100 | 100 | 100 |
| Water, mL | 400 | 400 | 400 | 400 |
| Sample | Hours of Fermentation [H] | Maltose [mg/g] | Glucose [mg/g] | Fructose [mg/g] |
|---|---|---|---|---|
| M0 | 0 | 5.807 ± 0.09 c | 3.689 ± 0.06 b c | 1.76 ± 0.09 a |
| M1 | 1.393 ± 0.02 a | 3.025 ± 0.03 b | 2.865 ± 0.07 b | |
| M2 | 3.552 ± 0.04 b | 4.251 ± 0.08 c | 2.774 ± 0.04 b | |
| M3 | 8.776 ± 0.08 d | 2.252 ± 0.05 a | 1.804 ± 0.09 a | |
| M0 | 12 | 6.465 ± 0.03 c | 3.489 ± 0.08 a | 2.861 ± 0.05 a b |
| M1 | 1.695 ± 0.04 a | 4.527 ± 0.07 a | 3.821 ± 0.06 c | |
| M2 | 3.347 ± 0.07 b | 4.63 ± 0.05 b | 3.183 ± 0.08 b | |
| M3 | 9.362 ± 0.06 d | 3.096 ± 0.03 a | 2.669 ± 0.06 a | |
| M0 | 24 | 1.389 ± 0.02 a | 1.466 ± 0.05 a | 1.056 ± 0.08 a |
| M1 | 1.113 ± 0.05 a | 4.986 ± 0.08 c | 2.065 ± 0.04 b | |
| M2 | 1.875 ± 0.01 a | 5.355 ± 0.09 c | 1.743 ± 0.06 b | |
| M3 | 11.339 ± 0.05 b | 2.709 ± 0.03 b | 0.513 ± 0.07 a |
| Sample | Hours of Fermentation [H] | Lactic Acid [mg/g] | Acetic Acid [mg/g] | Citric Acid [mg/g] | Ethanol [mg/g] | QF |
|---|---|---|---|---|---|---|
| M0 | 0 | 0.00 a | 0.00 a | 1.35 ± 0.03 a | 0.00 a | n.c. |
| M1 | 0.00 a | 0.00 a | 1.91 ± 0.03 b | 0.00 a | n.c. | |
| M2 | 0.00 a | 0.00 a | 2.09 ± 0.04 b | 0.00 a | n.c. | |
| M3 | 0.00 a | 0.00 a | 1.21 ± 0.02 a | 0.00 a | n.c. | |
| M0 | 12 | 0.40 ± 0.02 a | 0.36 ± 0.04 b | 1.72 ± 0.03 a | 0.14 ± 0.05 a | 1.11 |
| M1 | 1.85 ± 0.03 b | 0.14 ± 0.02 a | 2.50 ± 0.03 b | 0.16 ± 0.02 a | 13.21 | |
| M2 | 1.89 ± 0.04 b | 0.27 ± 0.02 b | 2.33 ± 0.05 b | 0.19 ± 0.01 a | 7.00 | |
| M3 | 0.41 ± 0.02 a | 0.00 c | 1.82 ± 0.03 a | 0.00 b | 0.00 | |
| M0 | 24 | 6.65 ± 0.06 c | 1.10 ± 0.03 b | 2.00 ± 0.04 a | 0.28 ± 0.03 a | 6.05 |
| M1 | 6.37 ± 0.02 c | 0.43 ± 0.05 a | 2.95 ± 0.03 b | 0.08 ± 0.02 a | 14.81 | |
| M2 | 5.43 ± 0.07 b | 0.41 ± 0.03 a | 2.66 ± 0.08 b | 0.08 ± 0.01 a | 13.24 | |
| M3 | 2.89 ± 0.03 a | 0.51 ± 0.02 a | 1.91 ± 0.07 a | 0.12 ± 0.03 a | 5.67 |
| Minerals | 0 h of Fermentation [H] | 12 h of Fermentation [H] | 24 h of Fermentation [H] | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| [mg/kg] | M0 | M1 | M2 | M3 | M0 | M1 | M2 | M3 | M0 | M1 | M2 | M3 |
| K | 346.93 ± 0.05 c | 347.3 ± 0.27 c | 334.22 ± 0.56 b | 332.86 ± 0.16 a | 353.76 ± 0.30 d | 350.63 ± 0.55 c | 345.87 ± 0.26 b | 338.730.34± a | 362.4 ± 0.14 b | 361.57 ± 0.15 b | 360.85 ± 0.15 b | 341.95 ± 0.15 a |
| Ca | 143.88 ± 0.04 b | 146.66 ± 0.33 c | 148.95 ± 0.11 d | 122.49 ± 0.34 a | 156.41 ± 0.23 c | 150.94 ± 0.21 b | 151.85 ± 0.30 b | 124.73 ± 0.20 a | 171.05 ± 0.17 c | 246.73 ± 0.30 d | 155.66 ± 0.20 b | 124.93 ± 0.12 a |
| Mg | 115.78 ± 0.08 ab | 114.99 ± 0.70 a | 115.84 ± 0.19 ab | 114.84 ± 0.21 a | 115.86 ± 0.37 a | 116.46 ± 0.20 a b | 116.26 ± 0.21 a b | 115.74 ± 0.20 a | 118.35 ± 0.35 c | 116.96 ± 0.17 b | 117.97 ± 0.17 c | 115.84 ± 0.19 a |
| Cd | 0.11 ± 0.07 a | 0.14 ± 0.03 a | 0.08 ± 0.02 a | 0.10 ± 0.03 a | 0.13 ± 0.05 a | 0.16 ± 0.05 a | 0.10 ± 0.03 a | 0.13 ± 0.11 a | 0.14 ± 0.05 a | 0.16 ± 0.05 a | 0.19 ± 0.04 a | 0.16 ± 0.05 a |
| Cu | 2.36 ± 0.09 a | 2.81 ± 0.15 a | 2.95 ± 0.14 a | 2.46 ± 0.14 a | 2.44 ± 0.26 a | 3.57 ± 0.29 b | 3.39 ± 0.17 b | 2.57 ± 0.12 a | 3.53 ± 0.11 b | 4.06 ± 0.10 bc | 3.59 ± 0.13 b | 2.79 ± 0.09 a |
| Zn | 14.89 ± 0.14 a | 24.85 ± 0.16 d | 18.74 ± 0.25 c | 16.84 ± 0.28 b | 14.90 ± 0.43 a | 34.20 ± 0.19 d | 18.96 ± 0.18 c | 16.98 ± 0.15 b | 15.36 ± 0.33 a | 36.07 ± 0.26 d | 19.85 ± 0.22 c | 17.23 ± 0.06 b |
| Cr | 0.18 ± 0.06 a | 0.38 ± 0.08 a | 0.25 ± 0.05 a | 0.35 ± 0.07 a | 0.27 ± 0.13 a | 0.39 ± 0.08 a | 0.29 ± 0.06 a | 0.4 ± 0.09 a | 0.69 ± 0.13 a | 0.63 ± 0.21 a | 0.32 ± 0.02 a | 0.43 ± 0.08 a |
| Mn | 21.75 ± 0.10 b | 34.61 ± 0.63 d | 25.83 ± 0.22 c | 10.20 ± 0.22 a | 22.34 ± 0.32 b | 43.99 ± 0.22 d | 27.62 ± 0.29 c | 12.69 ± 0.22 a | 22.86 ± 0.21 b | 46.70 ± 0.15 d | 28.95 ± 0.15 c | 13.85 ± 0.17 a |
| Ni | 0.35 ± 0.02 a | n.d. | 0.47 ± 0.10 a | n.d. | 0.75 ± 0.09 a | n.d. | 0.85 ± 0.09 a | n.d. | 0.90± a b | 0.57 ± 0.10 a | 0.94 ± 0.12 a b | 0.14 ± 0.04 a |
| Fe | 8.44 ± 0.03 a | 14.86 ± 0.19 c | 10.86 ± 0.23 b | 10.85 ± 0.23 b | 15.67 ± 0.19 b | 18.74 ± 0.16 c | 12.59 ± 0.19 a | 12.84 ± 0.17 a | 17.47± b | 19.29 ± 0.16 c | 13.96 ± 0.16 a | 13.46 ± 0.21 a |
| Pb | 5.46 ± 0.13 b | 4.14 ± 0.15 a | 6.49 ± 0.33 c | 7.50 ± 0.13 d | 6.77 ± 0.17 b | 4.9 ± 0.11 a | 7.48 ± 0.27 c | 8.24 ± 0.08 d | 7.29± a b | 6.70 ± 0.18 a | 8.93 ± 0.14 c | 8.94 ± 0.10 c |
| Volatile Compounds | 0 h of fermentation [H] | 12 h of Fermentation [H] | 24 h of Fermentation [H] | Odor Perception | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M0 | M1 | M2 | M3 | M0 | M1 | M2 | M3 | M0 | M1 | M2 | M3 | ||
| Alcohols | |||||||||||||
| 1-Butanol, 3-methyl- | 4.74 ± 0.07 c | 1.78 ± 0.11 a | 2.93 ± 0.09 b | n.d. | 12.33 ± 0.07 b | 3.82 ± 0.09 a | 3.42 ± 0.06 a | 23.52 ± 0.16 c | 7.04 ± 0.10 b | n.d. | n.d. | 2.77 ± 0.08 a | acid, spicy |
| 1-Pentanol | 17.85 ± 0.13 b | 8.69 ± 0.12 a | 30.21 ± 0.23 d | 23.79 ± 0.12 c | 5.74 ± 0.08 a | 8.00 ± 0.04 b | 19.74 ± 0.08 c | 7.90 ± 0.11 b | 5.83 ± 0.17 a | 8.26 ± 0.15 b | 20.00 ± 0.04 d | 12.09 ± 0.05 c | pungent |
| 1-Pentanol, 4-methyl- | 2.01 ± 0.07 | n.d. | n.d. | n.d. | 1.21 ± 0.09 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | nuts |
| 1-Hexanol | 15.70 ± 0.09 a | 29.14 ± 0.17 b | 35.16 ± 0.19 c | 51.96 ± 0.17 d | 9.75 ± 0.11 a | 24.88 ± 0.12 c | 20.45 ± 0.10 b | 51.50 ± 0.15 d | 11.74 ± 0.09 a | 47.60 ± 0.15 c | 29.94 ± 0.07 b | 69.5 ± 0.10 d | fruits |
| 1-Octen-3-ol | n.d. | n.d. | 2.27 ± 0.05 a | 3.08 ± 0.12 b | 0.93 ± 0.11 a | 1.48 ± 0.15 a b | n.d. | 2.22 ± 0.11 c | n.d. | n.d. | 1.91 ± 0.04 a | 2.93 ± 0.07 b | mushrooms |
| 1-Butanol, 2-methyl- | n.d. | n.d. | n.d. | n.d. | n.d. | 0.93 ± 0.13 | n.d. | n.d. | n.d. | 1.35 ± 0.08 | n.d. | n.d. | alcohol, wine |
| 1-Nonen-3-ol | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 1.37 ± 0.09 | n.d. | n.d. | n.d. | n.d. | n.d. | oil |
| 1-Heptanol | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 0.33 ± 0.04 | n.d. | n.d. | n.d. | n.d. | woody, fatty |
| 1-Penten-3-ol, 4-methyl- | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 1.16 ± 0.06 | n.d. | n.d. | n.d. | fresh |
| 2-Octen-1-ol | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 0.26 ± 0.08 a | n.d. | 0.92 ± 0.05 a | n.d. | citrus, green |
| 3-Buten-1-ol, 3-methyl- | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 6.31 ± 0.10 b | 4.62 ± 0.04 a | n.d. | sweet |
| 1-Butanol, 3-methyl- | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 6.31 ± 0.06 b | 5.05 ± 0.09 a | n.d. | pear, pungent |
| 2-Hexanol, 5-methyl- | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 0.99 ± 0.07 a | 0.30 ± 0.05 a | fruits |
| Aldehydes | |||||||||||||
| Hexanal | 40.37 ± 0.13 c | 46.09 ± 0.12 d | 18.23 ± 0.09 b | 13.19 ± 0.14 a | n.d. | 29.78 ± 0.12 b | 31.49 ± 0.15 c | 4.38 ± 0.11 a | n.d. | 4.19 ± 0.09 b | n.d. | 2.66 ± 0.08 a | grass, fatty |
| Heptanal | 5.07 ± 0.07 b | 2.69 ± 0.14 a | n.d. | n.d. | 0.65 ± 0.09 a | 2.07 ± 0.13 b | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | fruits |
| Benzaldehyde | 6.72 ± 0.10 c | 1.79 ± 0.06 ab | 1.13 ± 0.07 a | 0.71 ± 0.09 a | 1.13 ± 0.09 ab | 1.86 ± 0.09 b | 0.50 ± 0.11 a | 0.64 ± 0.11 a | 1.17 ± 0.05 ab | 2.80 ± 0.06 c | 0.85 ± 0.06 a | 0.30 ± 0.05 a | almonds |
| Octanal | n.d. | 1.15 ± 0.05 | n.d. | n.d. | n.d. | 0.45 ± 0.10 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | fatty, citrus |
| 2-Heptenal, (Z)- | n.d. | n.d. | n.d. | n.d. | n.d. | 0.91 ± 0.04 a | 0.71 ± 0.09 a | n.d. | 0.33 ± 0.07 | n.d. | n.d. | n.d. | green, fatty |
| Benzeneacetaldehyde | n.d. | n.d. | n.d. | n.d. | n.d. | 1.41 ± 0.11 ab | 0.87 ± 0.09 a | n.d. | 0.88 ± 0.10 a | 0.64 ± 0.09 a | 2.86 ± 0.08 b | n.d. | floral |
| 2-Octenal, (E)- | n.d. | n.d. | n.d. | n.d. | n.d. | 0.65 ± 0.08 a | 0.97 ± 0.12 a | n.d. | n.d. | n.d. | n.d. | n.d. | honey, nuts |
| Nonanal | n.d. | 1.08 ± 0.09 | n.d. | n.d. | n.d. | 0.35 ± 0.10 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | fatty, citrus |
| Decanal | n.d. | 0.58 ± 0.06 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | orange peel |
| Ketones | |||||||||||||
| Acetophenone | 7.53 ± 0.10 c | 1.63 ± 0.09 ab | 2.55 ± 0.05 b | 0.81 ± 0.09 a | 1.01 ± 0.10 ab | 0.49 ± 0.07 a | 0.32 ± 0.12 a | 0.18 ± 0.03 a | 0.74 ± 0.11 a | 1.07 ± 0.03 ab | 0.63 ± 0.050 a | 0.29 ± 0.03 a | floral |
| 2-Heptanone | n.d. | 0.58 ± 0.05 a | 1.79 ± 0.05 b | 1.89 ± 0.14 b | 0.30 ± 0.08 a | 2.12 ± 0.10 b | 1.73 ± 0.06 b | 0.53 ± 0.10 a | n.d. | 10.4 ± 0.04 b | 2.53 ± 0.06 a | n.d. | banana |
| 3-Octen-2-one | n.d. | n.d. | n.d. | 0.11 ± 0.02 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | cranberries |
| 2-Butanone, 3-hydroxy- | n.d. | n.d. | n.d. | n.d. | 1.24 ± 0.05 a | n.d. | n.d. | 4.83 ± 0.10 b | n.d. | 1.60 ± 0.04 | n.d. | n.d. | creamy |
| 2-Nonanone | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 0.26 ± 0.09 | n.d. | n.d. | 2.14 ± 0.04 b | 0.93 ± 0.09 a | n.d. | grass, fresh |
| Acids | |||||||||||||
| Butanoic acid | n.d. | n.d. | 0.93 ± 0.08 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 0.86 ± 0.08 | n.d. | n.d. | rancid cheese |
| Hexanoic acid | n.d. | n.d. | n.d. | n.d. | n.d. | 14.99 ± 0.13 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | urea |
| Benzoic acid | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 0.64 ± 0.10 a | n.d. | 0.91 ± 0.07 a | n.d. | balsamic |
| Others | |||||||||||||
| Butanoic acid, ethyl ester | n.d. | n.d. | n.d. | n.d. | 55.99 ± 0.13 b | n.d. | 11.31 ± 0.09 a | n.d. | 57.34 ± 0.12 c | n.d. | 22.24 ± 0.06 b | 1.32 ± 0.04 a | pineapple |
| Butanoic acid, propyl ester | n.d. | n.d. | n.d. | n.d. | 3.39 ± 0.12 | n.d. | n.d. | n.d. | 0.72 ± 0.08 | n.d. | n.d. | n.d. | apricots |
| Butanoic acid, butyl ester | n.d. | n.d. | n.d. | n.d. | 0.45 ± 0.12 | n.d. | n.d. | n.d. | 0.37 ± 0.07 | n.d. | n.d. | n.d. | pineapple |
| Hexanoic acid, ethyl ester | n.d. | n.d. | n.d. | n.d. | n.d. | 0.83 ± 0.07 a | n.d. | 0.46 ± 0.11 a | 0.86 ± 0.07 a | n.d. | n.d. | 1.22 ± 0.05 ab | fruits, sweet |
| Acetic acid, hexyl ester | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 0.30 ± 0.06 | n.d. | n.d. | n.d. | n.d. | 1.74 ± 0.10 | sweet, green |
| 1-Butanol, 3-methyl-, acetate | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 0.38 ± 0.06 | n.d. | n.d. | n.d. | pear, banana |
| Acetic acid, pentyl ester | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 0.23 ± 0.03 | fruits, banana |
| Furan, 2-pentyl- | n.d. | 2.35 ± 0.12 a | 2.42 ± 0.11 a | 3.14 ± 0.07 ab | 1.15 ± 0.05 a | 3.40 ± 0.10 b | 5.21 ± 0.09 c | 2.87 ± 0.08 b | 1.88 ± 0.08 a | 4.43 ± 0.04 bc | 4.66 ± 0.10 c | 3.59 ± 0.05 b | earth, green |
| Limonene | n.d. | n.d. | 2.39 ± 0.07 | n.d. | n.d. | 1.59 ± 0.07 b | n.d. | 0.66 ± 0.11 a | n.d. | 2.05 ± 0.08 b | n.d. | 0.85 ± 0.09 a | citrus |
| D-Limonene | n.d. | n.d. | n.d. | 1.30 ± 0.09 | n.d. | n.d. | 1.34 ± 0.08 | n.d. | n.d. | n.d. | 0.96 ± 0.08 | n.d. | mint, citrus |
| Disulfide, dimethyl | n.d. | n.d. | n.d. | n.d. | 4.72 ± 0.09 | n.d. | n.d. | n.d. | 7.41 ± 0.12 | n.d. | n.d. | n.d. | garlic |
| Dimethyl trisulfide | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 1.24 ± 0.06 | n.d. | n.d. | n.d. | sulfur |
| n.i. | n.d. | 2.44 ± 0.17 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Șerban, L.R.; Păucean, A.; Chiș, M.S.; Pop, C.R.; Man, S.M.; Pușcaș, A.; Ranga, F.; Socaci, S.A.; Alexa, E.; Berbecea, A.; et al. Metabolic Profile of Einkorn, Spelt, Emmer Ancient Wheat Species Sourdough Fermented with Strain of Lactiplantibacillus plantarum ATCC 8014. Foods 2023, 12, 1096. https://doi.org/10.3390/foods12051096
Șerban LR, Păucean A, Chiș MS, Pop CR, Man SM, Pușcaș A, Ranga F, Socaci SA, Alexa E, Berbecea A, et al. Metabolic Profile of Einkorn, Spelt, Emmer Ancient Wheat Species Sourdough Fermented with Strain of Lactiplantibacillus plantarum ATCC 8014. Foods. 2023; 12(5):1096. https://doi.org/10.3390/foods12051096
Chicago/Turabian StyleȘerban, Larisa Rebeca, Adriana Păucean, Maria Simona Chiș, Carmen Rodica Pop, Simona Maria Man, Andreea Pușcaș, Floricuța Ranga, Sonia Ancuța Socaci, Ersilia Alexa, Adina Berbecea, and et al. 2023. "Metabolic Profile of Einkorn, Spelt, Emmer Ancient Wheat Species Sourdough Fermented with Strain of Lactiplantibacillus plantarum ATCC 8014" Foods 12, no. 5: 1096. https://doi.org/10.3390/foods12051096