
NMR-Based Characterization of Wood Decay Fungi as Promising Novel Foods: Abortiporus biennis, Fomitopsis iberica and Stereum hirsutum Mycelia as Case Studies

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains
2.2. Mycelia Samples Preparation
2.3. Samples Extraction
2.4. NMR Analysis
3. Results and Discussion
3.1. NMR Assignment of Bligh–Dyer Extracts
3.2. Quantitative Metabolite Profile: Comparison between A. biennis, F. iberica and S. hirsutum Mycelia
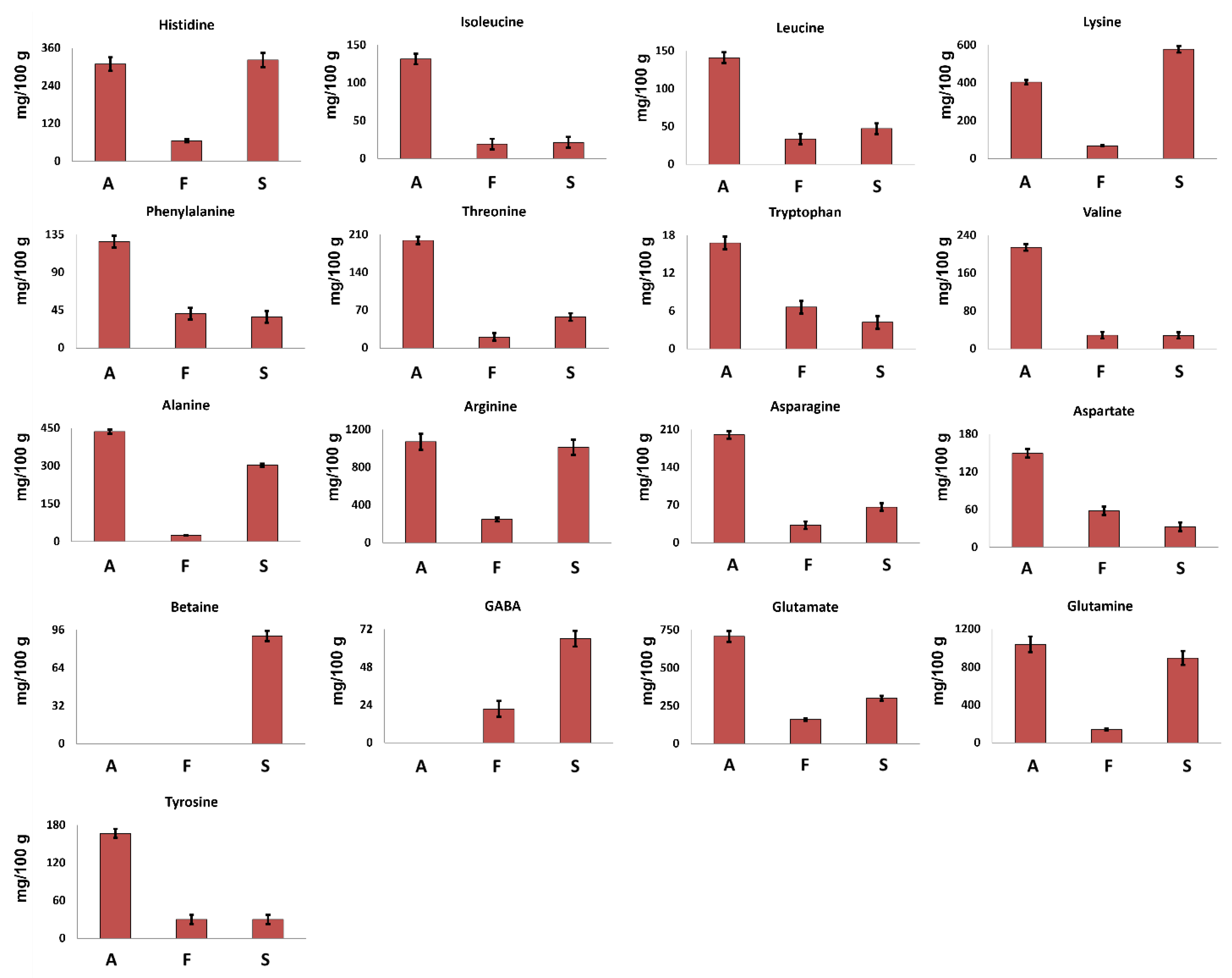
3.2.1. Amino Acids
3.2.2. Sugars
3.2.3. Organic Acids
3.2.4. Other Compounds
3.2.5. Apolar Fraction
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Li, T.; Cui, L.; Song, X.; Cui, X.; Wei, Y.; Tang, L.; Mu, Y.; Xu, Z. Wood decay fungi: An analysis of worldwide research. J. Soils Sediments 2022, 22, 1688–1702. [Google Scholar] [CrossRef]
- Girometta, C.; Dondi, D.; Baiguera, R.M.; Bracco, F.; Branciforti, D.S.; Buratti, S.; Lazzaroni, S.; Savino, E. Characterization of mycelia from wood-decay species by TGA and IR spectroscopy. Cellulose 2020, 27, 6133–6148. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the safety of ‘Chitin-glucan’ as a Novel Food ingredient. EFSA J. 2010, 8, 1687. [Google Scholar] [CrossRef]
- Turck, D.; Castenmiller, J.; de Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; Pelaez, C.; et al. Safety of vitamin D2 mushroom powder as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2020, 18, e05948. [Google Scholar] [CrossRef] [Green Version]
- European Commision Consultation Process on Novel Food Status. Available online: https://food.ec.europa.eu/safety/novel-food/consultation-process-novel-food-status_en (accessed on 26 April 2023).
- Swallah, M.S.; Bondzie-Quaye, P.; Wu, Y.; Acheampong, A.; Sossah, F.L.; Elsherbiny, S.M.; Huang, Q. Therapeutic potential and nutritional significance of Ganoderma lucidum—A comprehensive review from 2010 to 2022. Food Funct. 2023, 14, 1812–1838. [Google Scholar] [CrossRef]
- Wu, J.Y.; Siu, K.C.; Geng, P. Bioactive ingredients and medicinal values of Grifola frondosa (Maitake). Foods 2021, 10, 95. [Google Scholar] [CrossRef]
- Cartabia, M.; Girometta, C.E.; Milanese, C.; Baiguera, R.M.; Buratti, S.; Branciforti, D.S.; Vadivel, D.; Girella, A.; Babbini, S.; Savino, E.; et al. Collection and characterization of wood decay fungal strains for developing pure mycelium mats. J. Fungi 2021, 7, 1008. [Google Scholar] [CrossRef]
- Peiris, D.; Dunn, W.B.; Brown, M.; Kell, D.B.; Roy, I.; Hedger, J.N. Metabolite profiles of interacting mycelial fronts differ for pairings of the wood decay basidiomycete fungus, Stereum hirsutum with its competitors Coprinus micaceus and Coprinus disseminatus. Metabolomics 2008, 4, 52–62. [Google Scholar] [CrossRef]
- Mannina, L.; Sobolev, A.P.; Viel, S. Liquid state 1H high field NMR in food analysis. Prog. Nucl. Magn. Reson. Spectrosc. 2012, 66, 1–39. [Google Scholar] [CrossRef]
- Moco, S. Studying Metabolism by NMR-Based Metabolomics. Front. Mol. Biosci. 2022, 9, 372. [Google Scholar] [CrossRef]
- Marcone, M.F.; Wang, S.; Albabish, W.; Nie, S.; Somnarain, D.; Hill, A. Diverse food-based applications of nuclear magnetic resonance (NMR) technology. Food Res. Int. 2013, 51, 729–747. [Google Scholar] [CrossRef]
- Parlak, Y.; Guzeler, N. Nuclear magnetic resonance spectroscopy applications in foods. Curr. Res. Nutr. Food Sci. 2016, 4, 161–168. [Google Scholar] [CrossRef]
- Cartabia, M.; Girometta, C.E.; Baiguera, R.M.; Buratti, S.; Babbini, S.; Bernicchia, A.; Savino, E. Lignicolous Fungi Collected in Northern Italy: Identification and Morphological Description of Isolates. Diversity 2022, 14, 413. [Google Scholar] [CrossRef]
- Di Matteo, G.; Spano, M.; Esposito, C.; Santarcangelo, C.; Baldi, A.; Daglia, M.; Mannina, L.; Ingallina, C.; Sobolev, A.P. Nmr characterization of ten apple cultivars from the piedmont region. Foods 2021, 10, 289. [Google Scholar] [CrossRef]
- Ingallina, C.; Spano, M.; Sobolev, A.P.; Esposito, C.; Santarcangelo, C.; Baldi, A.; Daglia, M.; Mannina, L. Characterization of local products for their industrial use: The case of Italian potato cultivars analyzed by untargeted and targeted methodologies. Foods 2020, 9, 1216. [Google Scholar] [CrossRef] [PubMed]
- Spano, M.; Di Matteo, G.; Ingallina, C.; Botta, B.; Quaglio, D.; Ghirga, F.; Balducci, S.; Cammarone, S.; Campiglia, E.; Giusti, A.M.; et al. A multimethodological characterization of cannabis sativa L. Inflorescences from seven dioecious cultivars grown in Italy: The effect of different harvesting stages. Molecules 2021, 26, 2912. [Google Scholar] [CrossRef]
- Spano, M.; Maccelli, A.; Di Matteo, G.; Ingallina, C.; Biava, M.; Crestoni, M.E.; Bardaud, J.X.; Giusti, A.M.; Mariano, A.; D’abusco, A.S.; et al. Metabolomic profiling of fresh goji (Lycium barbarum L.) berries from two cultivars grown in central italy: A multi-methodological approach. Molecules 2021, 26, 5412. [Google Scholar] [CrossRef] [PubMed]
- Rahi, D.K.; Malik, D. Diversity of Mushrooms and Their Metabolites of Nutraceutical and Therapeutic Significance. J. Mycol. 2016, 2016, 7654123. [Google Scholar] [CrossRef]
- Sangtitanu, T.; Sangtanoo, P.; Srimongkol, P.; Saisavoey, T.; Reamtong, O.; Karnchanatat, A. Peptides obtained from edible mushrooms: Hericium erinaceus offers the ability to scavenge free radicals and induce apoptosis in lung cancer cells in humans. Food Funct. 2020, 11, 4927–4939. [Google Scholar] [CrossRef]
- Chang, H.; Chao, G.; Chen, C.; Mau, J. Non-volatile taste components of Agaricus blazei, Antrodia camphorata and Cordyceps militaris mycelia. Food Chem. 2001, 74, 203–207. [Google Scholar] [CrossRef]
- Tsai, S.Y.; Weng, C.C.; Huang, S.J.; Chen, C.C.; Mau, J.L. Nonvolatile taste components of Grifola frondosa, Morchella esculenta and Termitomyces albuminosus mycelia. LWT 2006, 39, 1066–1071. [Google Scholar] [CrossRef]
- Aminuddin, H.; Khan, A.M.; Abidin, H.; Madzlan, K.; Suri, R.; Kamal, M.K.; Hussin, A.; Ayob, K.; Hamid, A.; Kasran, M.; et al. Optimization of submerged culture for the production of Lentinula edodes mycelia biomass and amino acid composition by different temperatures (Pengoptimuman kultur ampaian dalam penghasilan biojisim miselium Lentinula edodes dan komposisi asid amino pada. J. Trop. Agric. Food Sci. 2007, 35, 131–138. [Google Scholar]
- FAO Part I. Amino-Acid Content of Foods. Available online: https://www.fao.org/3/AC854T/AC854T03.htm#chI.I (accessed on 22 June 2023).
- Jeong, S.J.; Stitham, J.; Evans, T.D.; Zhang, X.; Rodriguez-Velez, A.; Yeh, Y.S.; Tao, J.; Takabatake, K.; Epelman, S.; Lodhi, I.J.; et al. Trehalose causes low-grade lysosomal stress to activate TFEB and the autophagy-lysosome biogenesis response. Autophagy 2021, 17, 3740–3752. [Google Scholar] [CrossRef]
- Liaud, N.; Giniés, C.; Navarro, D.; Fabre, N.; Crapart, S.; Gimbert, I.H.; Levasseur, A.; Raouche, S.; Sigoillot, J.-C. Exploring fungal biodiversity: Organic acid production by 66 strains of filamentous fungi. Fungal Biol. Biotechnol. 2014, 1, 1. [Google Scholar] [CrossRef] [Green Version]
- Valentão, P.; Lopes, G.; Valente, M.; Barbosa, P.; Andrade, P.B.; Silva, B.M.; Baptista, P.; Seabra, R.M. Quantitation of nine organic acids in wild mushrooms. J. Agric. Food Chem. 2005, 53, 3626–3630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barros, L.; Pereira, C.; Ferreira, I.C.F.R. Optimized Analysis of Organic Acids in Edible Mushrooms from Portugal by Ultra Fast Liquid Chromatography and Photodiode Array Detection. Food Anal. Methods 2013, 6, 309–316. [Google Scholar] [CrossRef]
- Zeisel, S.H.; Da Costa, K.A. Choline: An essential nutrient for public health. Nutr. Rev. 2009, 67, 615–623. [Google Scholar] [CrossRef] [Green Version]
- Zeisel, S.H. Nutritional Importance of Choline for Brain Development. J. Am. Coll. Nutr. 2004, 23, 621S–626S. [Google Scholar] [CrossRef]
- Zeisel, S.H.; Klatt, K.C.; Caudill, M.A. Choline. Adv. Nutr. 2018, 9, 58–60. [Google Scholar] [CrossRef] [Green Version]
- Baur, A.C.; Kühn, J.; Brandsch, C.; Hirche, F.; Stangl, G.I. Intake of ergosterol increases the vitamin D concentrations in serum and liver of mice. J. Steroid Biochem. Mol. Biol. 2019, 194, 105435. [Google Scholar] [CrossRef]
- Ruess, L.; Häggblom, M.M.; García Zapata, E.J.; Dighton, J. Fatty acids of fungi and nematodes—Possible biomarkers in the soil food chain? Soil Biol. Biochem. 2002, 34, 745–756. [Google Scholar] [CrossRef]
- Zhang, Y.; Mills, G.L.; Nair, M.G. Cyclooxygenase inhibitory and antioxidant compounds from the mycelia of the edible mushroom Grifola frondosa. J. Agric. Food Chem. 2002, 50, 7581–7585. [Google Scholar] [CrossRef] [PubMed]
- Lv, G.-P.; Zhao, J.; Duan, J.-A.; Tang, Y.-P.; Li, S.-P. Comparison of sterols and fatty acids in two species of Ganoderma. Chem. Cent. J. 2012, 6, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stahl, P.D.; Klug, M.J. Characterization and differentiation of filamentous fungi based on fatty acid composition. Appl. Environ. Microbiol. 1996, 62, 4136–4146. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | Assignment | 1H (ppm) | Multiplicity [J (Hz)] | 13C (ppm) |
|---|---|---|---|---|
| Hydroalcoholic extract | ||||
| Amino acids and derivatives | ||||
| Alanine a,b,c | COO- | 177.0 | ||
| α-CH | 3.80 | 51.6 | ||
| β-CH3 | 1.49 * | d [7.3] | 17.2 | |
| Arginine a,b,c | α-CH | 3.76 | ||
| β-CH2 | 1.93 | |||
| γ-CH | 1.66 * | m | 25.0 | |
| γ′-CH | 1.74 | m | 25.0 | |
| δ-CH3 | 3.25 | |||
| Asparagine a,b,c | α-CH | 4.02 | ||
| β, β′-CH2 | 2.87; 2.95 * | dd [7.4; 16.9] | ||
| Aspartate a,b,c | α-CH | 3.92 | ||
| β, β′-CH2 | 2.72; 2.81 * | dd [17.4;3.8] | ||
| Betaine c | N(CH3)3+ | 3.27 * | s | |
| GABA b,c | α-CH2 | 2.30 * | t [7.4] | 35.4 |
| β-CH2 | 1.94 | 24.3 | ||
| γ-CH2 | 3.02 | 39.8 | ||
| Glutamate a,b,c | α-CH | 3.78 | 55.6 | |
| β, β′-CH2 | 2.07;2.14 | 28.0 | ||
| γ-CH2 | 2.36 * | m | 34.8 | |
| Glutamine a,b,c | α-CH | 3.78 | ||
| β, β′-CH2 | 2.15 | |||
| γ-CH2 | 2.46 * | m | ||
| Histidine a,b,c | CH-3, ring | 7.94 | s | |
| CH-5, ring | 7.12 * | s | ||
| Isoleucine a,b,c | α-CH | 3.69 | 60.7 | |
| β-CH | 1.99 | 37.1 | ||
| γ-CH3 | 1.02 * | d [7.1] | 15.8 | |
| γ′-CH | 1.27 | 25.9 | ||
| δ-CH3 | 0.94 | t [7.4] | 12.3 | |
| Leucine a,b,c | α-CH | 3.74 | ||
| β-CH2 | 1.72 | 40.8 | ||
| δ, δ′-CH3 | 0.96; 0.97 * | d [6.2] | 25.7 | |
| Lysine a,b,c | α-CH | 3.77 | ||
| β-CH2 | 1.93 | |||
| γ-CH2 | 1.49 | 22.7 | ||
| δ-CH2 | 1.74 | 27.7 | ||
| ε-CH2 | 3.03 * | t [7.3] | 40.2 | |
| Phenylalanine a,b,c | CH-2,6 ring | 7.34 | m | |
| CH-3,5 ring | 7.43 * | m | ||
| CH-4 ring | 7.38 | m | ||
| Threonine a,b,c | α-CH | 3.58 | 61.7 | |
| β-CH | 4.27 | 67.0 | ||
| γ-CH3 | 1.34 * | d [6.6] | 21.2 | |
| Tryptophan a,b,c | CH-4, ring | 7.74 | d [8.1] | |
| CH-5, ring | 7.21 | |||
| CH-6, ring | 7.29 | |||
| CH-7, ring | 7.55 * | d [8.1] | ||
| Tyrosine a,b,c | CH-2,6 ring | 6.90 * | d [8.6] | 116.9 |
| CH-3,5 ring | 7.20 | d [8.6] | 129.5 | |
| Valine a,b,c | α-CH | 3.63 | 61.8 | |
| β-CH | 2.29 | 30.3 | ||
| γ-CH3 | 0.99 | d [7.06] | 18.0 | |
| γ′-CH3 | 1.05 * | d [7.06] | 19.2 | |
| Sugars and polyols | ||||
| β-Galactose b | CH-1 | 4.60 * | d [7.9] | |
| CH-2 | 3.51 | |||
| CH-3 | 3.67 | |||
| α-Glucose a,b,c | CH-1 | 5.24 | d [3.8] | |
| CH-2 | 3.56 | 72.7 | ||
| CH-3 | 3.74 | 74.2 | ||
| CH-4 | 3.42 | 71.3 | ||
| CH-5 | 3.84 | |||
| β-Glucose a,b,c | CH-1 | 4.65 * | d [7.8] | 97.0 |
| CH-2 | 3.28 | 75.5 | ||
| CH-3 | 3.50 | 76.9 | ||
| CH-4 | 3.43 | |||
| α-Maltose a | CH-1 | 5.24 | d [3.6] | |
| CH-2 | 3.57 | 72.7 | ||
| CH-3 | 3.74 | 74.2 | ||
| CH-4 | 3.45 | 71.3 | ||
| CH-5 | 3.88 | 72.5 | ||
| CH2-6 | 3.98 | 60.1 | ||
| CH-1′ | 5.42 * | d [3.8] | ||
| CH-2′ | 3.62 | 72.7 | ||
| CH-3′ | 3.70 | 73.9 | ||
| CH-4′ | 78.0 | |||
| β-Maltose a | CH-1 | 4.67 | d [7.8] | 97.4 |
| CH-2 | 3.30 | 72.7 | ||
| CH-3 | 3.79 | 74.2 | ||
| CH-4 | 3.65 | 71.3 | ||
| CH-1′ | 5.42 * | d [3.8] | ||
| CH-2′ | 3.62 | 72.7 | ||
| CH-3′ | 3.70 | 73.9 | ||
| CH-4′ | 78.0 | |||
| Mannitol c | CH-1,6 | 3.68 | dd [6.2; 11.9] | |
| CH-1′,6′ | 3.87 | dd [2.9; 11.9] | ||
| CH-2,5 | 3.77 | m | ||
| CH-3,4 | 3.80 * | m | ||
| Trehalose a,b,c | CH-1 | 5.20 * | d [3.8] | |
| CH-2 | 3.64 | |||
| CH-3 | 3.86 | |||
| CH-4 | 3.46 | |||
| Organic acids | ||||
| Acetate a,c | COO- | 182.9 | ||
| α-CH3 | 1.92 * | s | 24.4 | |
| Citrate a,b,c | α, γ-CH | 2.55 * | d [15.9] | 46.9 |
| α′, γ′-CH | 2.66 | d [15.9] | 46.9 | |
| β-C | 76.5 | |||
| 1,5-COO- | 180.3 | |||
| 6-COO- | 183.2 | |||
| Formate a,b,c | HCOO- | 8.46 * | s | |
| Fumarate a,b,c | α, β-CH=CH | 6.53 * | s | |
| Lactate a,b,c | α-CH | 4.13 | ||
| β-CH3 | 1.33 * | d [6.6] | ||
| Malate a,b,c | α-CH | 4.30 * | dd [9.8; 3.2] | |
| β-CH | 2.67 | dd [15.6; 3.2] | ||
| β′-CH | 2.39 | dd [15.6; 9.8] | ||
| Succinate a,b,c | α, β-CH2 | 2.41 * | s | |
| Other metabolites | ||||
| Adenosine a,b,c | CH-2 | 8.36 * | s | |
| CH-8 | 8.27 | s | ||
| CH-1′ | 6.08 | d [6.2] | ||
| Choline a,b,c | N(CH3)3+ | 3.21 * | s | 55.2 |
| α-CH2 | 68.7 | |||
| Uridine a,b,c | CH-5 | 5.90 | d [8.1] | |
| CH-6 | 7.88 * | d [8.1] | ||
| CH-1′ | 5.92 | d [4.5] | ||
| Uracil a,c | CH-5 | 5.80 * | d [8.1] | |
| CH-6 | 7.54 | d [8.1] | ||
| Organic extract | ||||
| Mono-unsaturated fatty chain a,b,c | COO | 173.9 | ||
| (Cn:1 Δ9) | CH2-2 | 2.28 * | 34.6 | |
| CH2-3 | 1.57 | m | 25.4 | |
| CH2-4,7 | 1.30 | m | 29.5 | |
| CH2-8 | 2.01 | m | 27.6 | |
| CH=CH 9,10 | 5.33 * | m | 130.4 | |
| CH2-11 | 2.01 | m | 27.6 | |
| CH2 | 1.33–1.28 | m | 29.8–32.0 | |
| CH2-n-1 | 1.26 | m | 22.9 | |
| CH3-n | 0.87 | t | 14.2 | |
| Di-unsaturated fatty chain a,b,c | COO | 173.9 | ||
| (Cn:2 Δ9,12) | CH2-2 | 2.30 * | 34.2 | |
| CH2-3 | 1.57 | m | 25.3 | |
| CH2-4,7 | 1.32–1.28 | m | 29.8 | |
| CH2-8 | 2.09 | m | 27.2 | |
| CH=9 | 5.35 * | m | 130.4 | |
| CH=10 | 5.33 * | m | 128.2 | |
| CH2-11 | 2.77 * | t [6.7] | 26.0 | |
| CH=12 | 5.33 * | m | 128.2 | |
| CH=13 | 5.35 * | m | 130.4 | |
| CH2-14 | 2.09 | m | 27.2 | |
| CH2 | 1.26–1.27 | m | 29.8–32.0 | |
| CH2-n-1 | 1.23 | m | 22.5 | |
| CH3-n | 0.89 | t [6.4] | 14.2 | |
| Di-unsaturated fatty chain a,b,c | COO | 173.9 | ||
| (Cn:2 Δ9,12) | CH2-2 | 2.30 * | 34.2 | |
| CH2-3 | 1.57 | m | 25.3 | |
| CH2-4,7 | 1.32–1.28 | m | 29.8 | |
| CH2-8 | 2.09 | m | 27.2 | |
| CH=9 | 5.35 * | m | 130.4 | |
| CH=10 | 5.33 * | m | 128.2 | |
| CH2-11 | 2.77 * | t [6.7] | 26.0 | |
| CH=12 | 5.33 * | m | 128.2 | |
| CH=13 | 5.35 * | m | 130.4 | |
| CH2-14 | 2.09 | m | 27.2 | |
| CH2 | 1.26–1.27 | m | 29.8–32.0 | |
| CH2-n-1 | 1.23 | m | 22.5 | |
| CH3-n | 0.89 | t [6.4] | 14.2 | |
| Saturated fatty acids a,b,c | COO | 173.9 | ||
| CH2-2 | 2.28 * | 34.6 | ||
| CH2-3 | 1.57 | m | 25.4 | |
| CH2 | 1.28–1.22 | m | 29.8–32.0 | |
| CH2 n-1 | 1.25 | 22.5 | ||
| CH3 n | 0.87 | t | 14.2 | |
| Ergosterol a,b,c | CH=6 | 5.55 * | ||
| CH=7 | 5.43 | |||
| CH2-12 | 41.6 | |||
| C-13 | 45.7 | |||
| CH-14 | 58.6 | |||
| CH-17 | 57.0 | |||
| CH3-18 | 0.66 | s | 12.4 | |
| CH=22,23 | 5.27 | m | ||
| 1,2-Diacyl-sn-glycero-3-phosphatidylethanolamine a,b,c | CH2N | 3.10 * | t [5.0] | |
| CH2OP | 4.08 | |||
| CH2 sn1 | 4.45; 4.16 | 62.2 | ||
| CH sn2 | 5.28 | 69.1 | ||
| CH2 sn3 | 4.05 | 64.7 | ||
| 1,2-Diacyl-sn-glycero-3-phosphatidylcholine a,b,c | +N(CH3)3 | 3.21 * | s | 54.5 |
| CH2N+ | 3.64 | 66.7 | ||
| CH2OP | 4.31 | |||
| CH2 sn1 | 4.45; 4.16 | 62.2 | ||
| CH sn2 | 5.28 | 69.1 | ||
| CH2 sn3 | 4.05 | 64.7 | ||
| Amino Acid | A. biennis (mg/g) | A. blazei Chang et al., 2001 [21] (mg/g) | G. frondosa Tsai et al., 2006 [22] (mg/g) | L. edodes Aminuddin et al., 2007 [23] (mg/g) |
|---|---|---|---|---|
| Leucine | 1.41 ± 0.02 | 0.31 ± < 0.1 | 4.92 ± 0.04 | 0.83 ± 0.01 |
| Isoleucine | 1.32 ± 0.02 | 0.21 ± < 0.1 | 2.80 ± 0.02 | 0.66 ± 0.04 |
| Valine | 2.15 ± 0.03 | 1.76 ± 0.07 | 4.13 ± 0.11 | 0.62 ± 0.01 |
| Histidine | 3.08 ± 0.06 | 0.66 ± 0.28 | 4.1 ± 0.17 | 0.26 ± 0.02 |
| Lysine | 4.04 ± 0.45 | 0.61 ± < 0.1 | 0.22 ± 0.01 | 0.90 ± 0.03 |
| Methionine | ND 1 | 0.67 ± 0.27 | 2.67 ± 0.10 | 0.06 ± 0.01 |
| Phenylalanine | 1.27 ± 0.03 | 0.17 ± < 0.1 | 1.66 ± 0.42 | 0.61 ± 0.01 |
| Threonine | 1.99 ± 0.07 | 0.53 ± 0.03 | 8.23 ± 0.36 | 0.58 ± 0.02 |
| Tryptophan | 0.16 ± 0.03 | ND | ND | ND |
| Alanine | 4.37 ± 0.11 | 1.05 ± 0.09 | 3.26 ± 0.25 | 0.82 ± 0.01 |
| Arginine | ND | 0.45 ± 0.01 | 0.97 ± 0.04 | 0.88 ± 0.01 |
| Asparagine | 2.00 ± 0.04 | ND | ND | ND |
| Aspartate | 1.49 ± 0.04 | 0.50 ± 0.06 | 2.75 ± 0.12 | 1.16 ± 0.03 |
| Glutamate | 7.07 ± 0.03 | ND | 3.76 ± 0.26 | 2.02 ± 0.13 |
| Glutamine | 10.38 ± 0.44 | ND | ND | ND |
| Glycine | ND | ND | 1.93 ± 0.04 | 0.47 ± 0.04 |
| Proline | ND | ND | ND | 0.69 ± 0.02 |
| Serine | ND | 0.09 ± < 0.10 | 2.73 ± 0.20 | 0.69 ± 0.02 |
| Tyrosine | 1.67 ± 0.05 | ND | 2.15 ± 0.10 | 0.32 ± 0.01 |
| Amino Acid | A. biennis Mycelium (mg/g) | Buckwheat Fagopyrum sagittatum FAO (mg/g) | Potato Solanum tuberosum FAO (mg/g) | Lentil Lens culinaris FAO (mg/g) | Palm Kernel Elaeis guineensis FAO (mg/g) | Lettuce Lactuca sativa FAO (mg/g) | Avocado Persea armeniaca FAO (mg/g) |
|---|---|---|---|---|---|---|---|
| Leucine | 1.41 ± 0.02 | 7.2 | 1.21 | 18.47 | 4.21 | 0.83 | 0.76 |
| Isoleucine | 1.32 ± 0.02 | 4.15 | 0.76 | 10.45 | 2.4 | 0.5 | 0.47 |
| Valine | 2.15 ± 0.03 | 8.1 | 0.93 | 12.11 | 3.77 | 0.71 | 0.63 |
| Histidine | 3.08 ± 0.06 | 2.6 | 0.3 | 6.62 | 1.47 | 0.21 | 0.25 |
| Lysine | 4.04 ± 0.45 | 4.6 | 0.96 | 17.39 | 2.46 | 0.5 | 0.59 |
| Methionine | ND 1 | 1.8 | 0.26 | 1.94 | 1.64 | 0.24 | 0.29 |
| Phenylalanine | 1.27 ± 0.03 | 4.6 | 0.8 | 12.66 | 2.56 | 0.67 | 0.48 |
| Threonine | 1.99 ± 0.07 | 4.4 | 0.75 | 9.6 | 2.22 | 0.54 | 0.4 |
| Tryptophan | 0.16 ± 0.03 | ND | ND | ND | ND | ND | ND |
| Alanine | 4.37 ± 0.11 | 5.7 | 0.89 | 10.41 | 2.83 | 0.56 | 0.82 |
| Arginine | ND | 11.96 | 1 | 21.01 | 9.33 | 0.59 | 0.47 |
| Asparagine | 2 ± 0.04 | ND | ND | ND | ND | ND | ND |
| Aspartate | 1.49 ± 0.04 | 10.8 | 2.48 | 27.98 | 5.75 | 1.51 | 3.11 |
| Glutamate | 7.07 ± 0.03 | 2.9 | 0.12 | 2.21 | 1.23 | ND | ND |
| Glutamine | 10.38 ± 0.44 | 21.14 | 2.04 | 40.13 | 11.99 | 1.34 | 1.69 |
| Glycine | ND | ND | ND | ND | ND | ND | ND |
| Proline | ND | 7.8 | 0.76 | 10.22 | 3.13 | 0.54 | 0.55 |
| Serine | ND | 5.3 | 0.75 | 10.33 | 2.29 | 0.68 | 0.54 |
| Tyrosine | 1.67 ± 0.05 | 6.1 | 0.83 | 12.73 | 3.28 | 0.43 | 0.56 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goppa, L.; Spano, M.; Baiguera, R.M.; Cartabia, M.; Rossi, P.; Mannina, L.; Savino, E. NMR-Based Characterization of Wood Decay Fungi as Promising Novel Foods: Abortiporus biennis, Fomitopsis iberica and Stereum hirsutum Mycelia as Case Studies. Foods 2023, 12, 2507. https://doi.org/10.3390/foods12132507
Goppa L, Spano M, Baiguera RM, Cartabia M, Rossi P, Mannina L, Savino E. NMR-Based Characterization of Wood Decay Fungi as Promising Novel Foods: Abortiporus biennis, Fomitopsis iberica and Stereum hirsutum Mycelia as Case Studies. Foods. 2023; 12(13):2507. https://doi.org/10.3390/foods12132507
Chicago/Turabian StyleGoppa, Lorenzo, Mattia Spano, Rebecca Michela Baiguera, Marco Cartabia, Paola Rossi, Luisa Mannina, and Elena Savino. 2023. "NMR-Based Characterization of Wood Decay Fungi as Promising Novel Foods: Abortiporus biennis, Fomitopsis iberica and Stereum hirsutum Mycelia as Case Studies" Foods 12, no. 13: 2507. https://doi.org/10.3390/foods12132507