
Glucosinolate Diversity Analysis in Choy Sum (Brassica rapa subsp. chinensis var. parachinensis) Germplasms for Functional Food Breeding



Abstract
:1. Introduction
2. Materials and Methods
2.1. GSLs Standards Used in This Experiment
2.2. Choy sum Genetic Materials and Cultivation Condition
2.3. Sample Preparation: Pretreatment and Extraction
2.4. Identification and Quantification of GSLs Using UPLC-MS/MS
2.5. Statistical Analysis
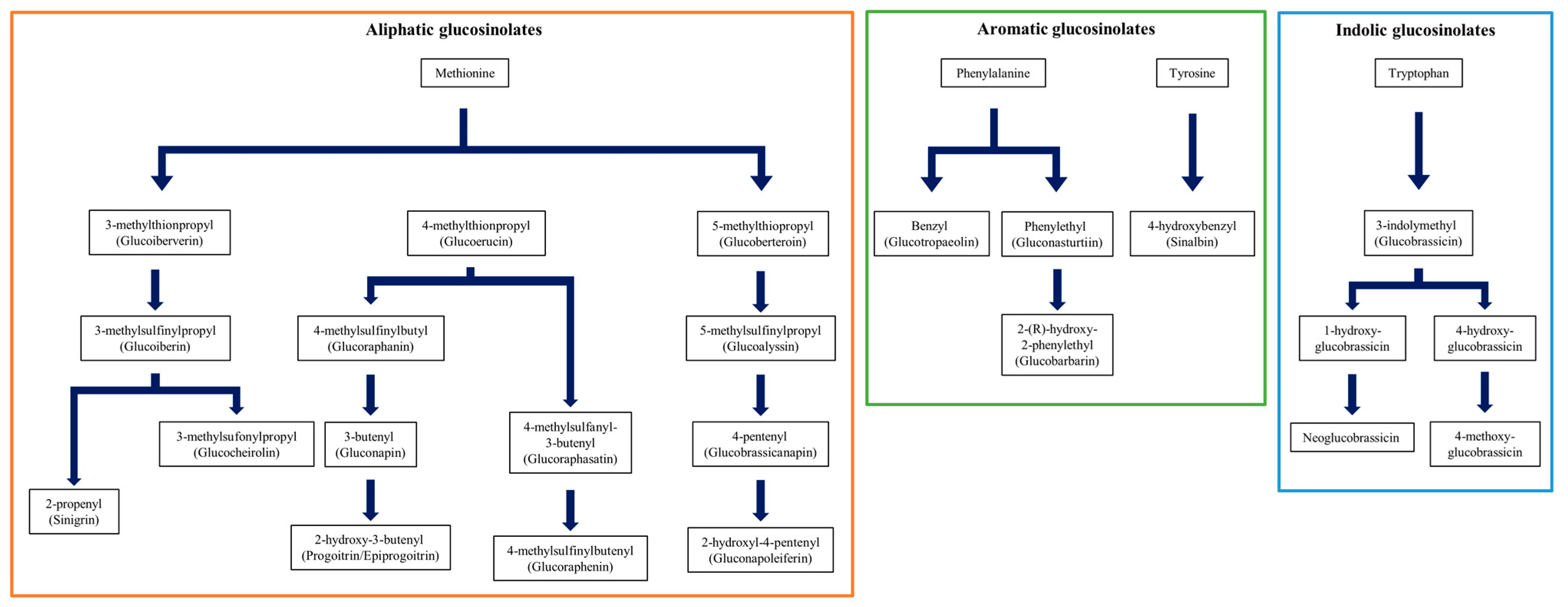
3. Results and Discussion
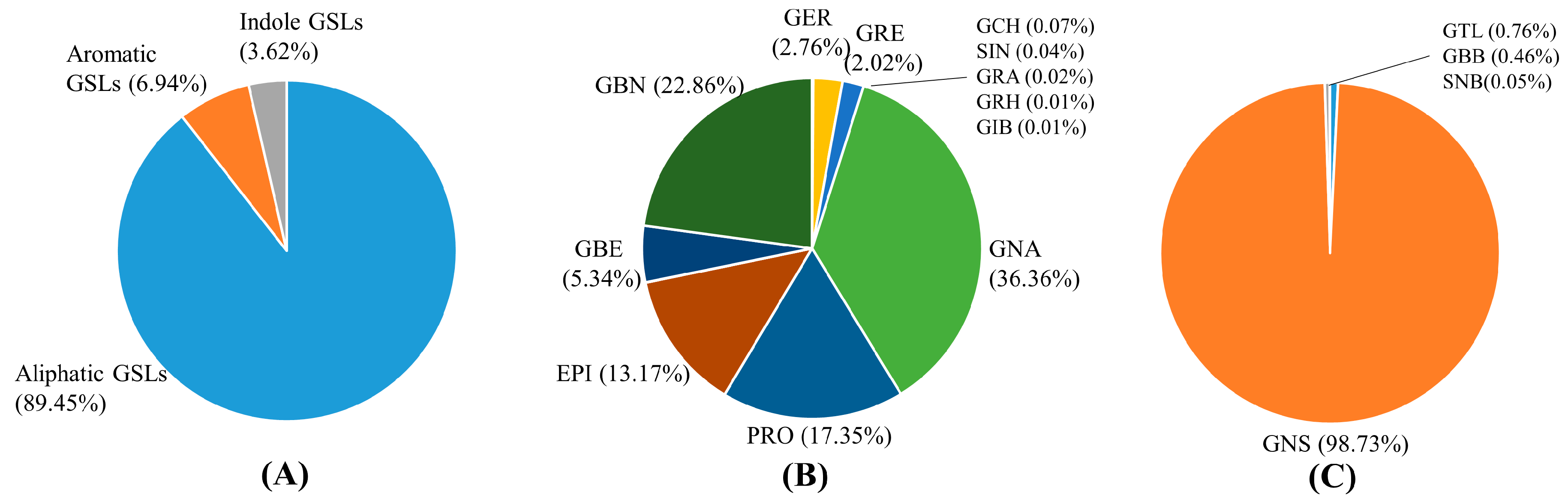
3.1. Quantification of GSLs and Selection of Candidate Germplasm for Breeding Materials
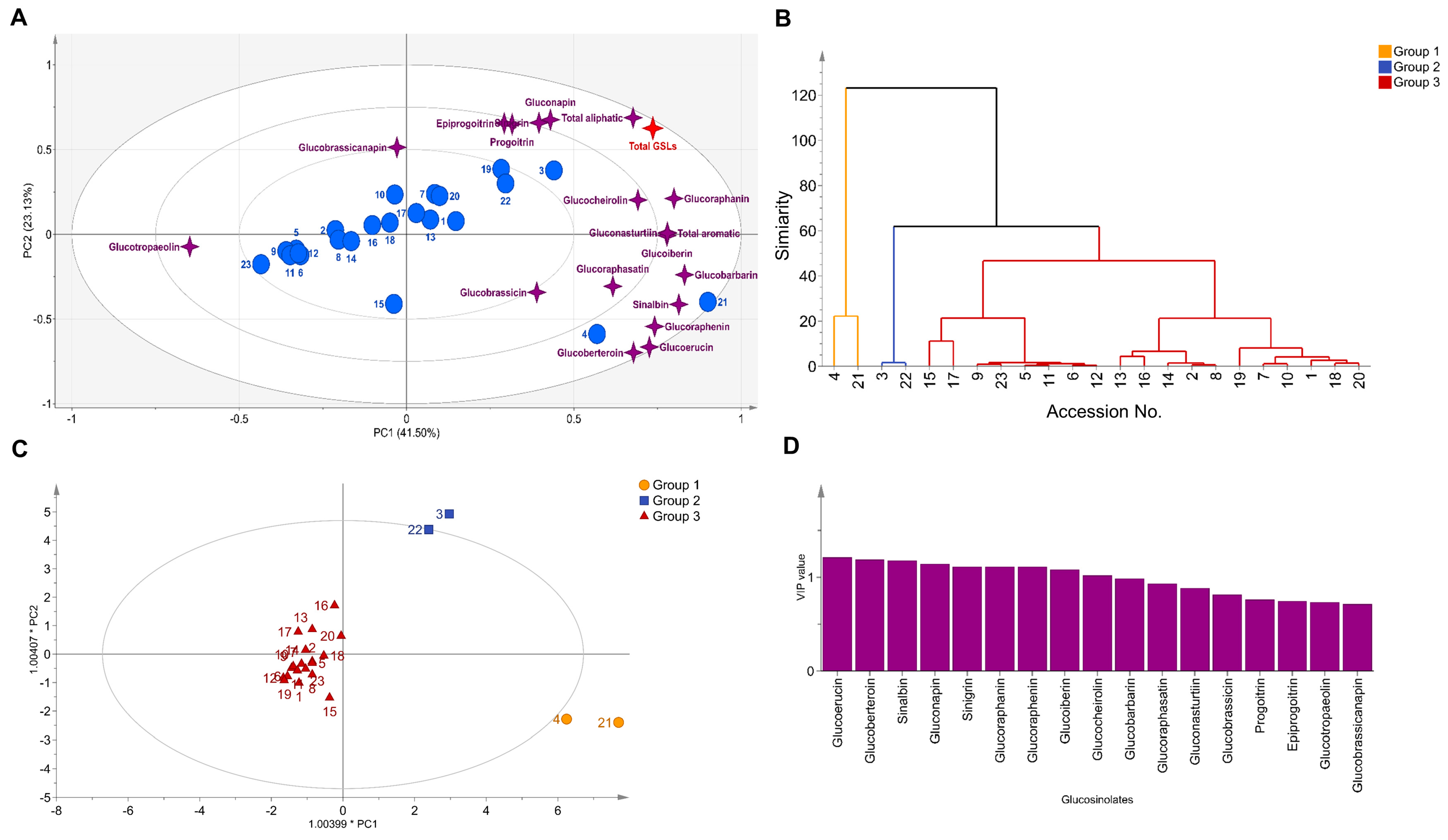
3.2. Correlation Analysis
3.3. Diversity Analysis and Clustering
3.4. Nutritional Value of Glucosinolates
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Foo, J.T.S. A Guide to Common Vegetables; Singapore Science Centre: Singapore, 1996. [Google Scholar]
- Edward, E. Modelling the Growth of Choy Sum (Brassica chinensis var. parachinensis), at Different Nitrogen Fertilizer Rates; Universiti Putra Malaysia: Serdang, Malaysia, 2008. [Google Scholar]
- Huang, J.J.; D’Souza, C.; Tan, M.Q.; Zhou, W. Light intensity plays contrasting roles in regulating metabolite compositions in choy sum (Brassica rapa var. parachinensis). J. Agric. Food Chem. 2021, 69, 5318–5331. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Tan, W.K.; Du, Y.; Lee, H.W.; Liang, X.; Lei, J.; Striegel, L.; Weber, N.; Rychlik, M.; Ong, C.N. Nutritional metabolites in Brassica rapa subsp. chinensis var. parachinensis (choy sum) at three different growth stages: Microgreen, seedling and adult plant. Food Chem. 2021, 357, 129535. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Shi, Q.; Wang, X.; Wei, M.; Hu, J.; Liu, J.; Yang, F. Influence of cow manure vermicompost on the growth, metabolite contents, and antioxidant activities of Chinese cabbage (Brassica campestris ssp. chinensis). Biol. Fertil. Soils 2010, 46, 689–696. [Google Scholar] [CrossRef]
- Domínguez-Perles, R.; Mena, P.; Garcia-Viguera, C.; Moreno, D. Brassica foods as a dietary source of vitamin C: A review. Crit. Rev. Food Sci. Nutr. 2014, 54, 1076–1091. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, J.; Wei, Q.; Li, B.; Zhong, X.; Hu, T.; Hu, H.; Bao, C. Transcriptome-wide identification and characterization of circular RNAs in leaves of Chinese cabbage (Brassica rapa L. ssp. pekinensis) in response to calcium deficiency-induced tip-burn. Sci. Rep. 2019, 9, 14544. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Zhou, Q.-h.; Xu, K. Are isothiocyanates potential anti-cancer drugs? Acta Pharmacol. Sin. 2009, 30, 501–512. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Wang, H.; Xie, J.; Lv, J.; Zhang, G.; Hu, L.; Luo, S.; Li, L.; Yu, J. The roles of cruciferae glucosinolates in disease and pest resistance. Plants 2021, 10, 1097. [Google Scholar] [CrossRef]
- Wittstock, U.; Halkier, B.A. Glucosinolate research in the Arabidopsis era. Trends Plant Sci. 2002, 7, 263–270. [Google Scholar] [CrossRef]
- Halkier, B.A.; Du, L. The biosynthesis of glucosinolates. Trends Plant Sci. 1997, 2, 425–431. [Google Scholar] [CrossRef]
- Bouranis, J.A.; Beaver, L.M.; Choi, J.; Wong, C.P.; Jiang, D.; Sharpton, T.J.; Stevens, J.F.; Ho, E. Composition of the gut microbiome influences production of sulforaphane-nitrile and iberin-nitrile from glucosinolates in broccoli sprouts. Nutrients 2021, 13, 3013. [Google Scholar] [CrossRef]
- Park, S.; Son, H.-K.; Chang, H.-C.; Lee, J.-J. Effects of cabbage-apple juice fermented by Lactobacillus plantarum EM on lipid profile improvement and obesity amelioration in rats. Nutrients 2020, 12, 1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandrich, L.; Caputo, E. Brassicaceae-derived anticancer agents: Towards a green approach to beat cancer. Nutrients 2020, 12, 868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takasugi, M.; Katsui, N.; Shirata, A. Isolation of three novel sulphur-containing phytoalexins from the chinese cabbage Brassica campestris L. ssp. pekinensis (cruciferae). J. Chem. Soc. Chem. Commun. 1986, 14, 1077–1078. [Google Scholar] [CrossRef]
- Le, T.N.; Chiu, C.-H.; Hsieh, P.-C. Bioactive compounds and bioactivities of Brassica oleracea L. var. italica sprouts and microgreens: An updated overview from a nutraceutical perspective. Plants 2020, 9, 946. [Google Scholar] [CrossRef] [PubMed]
- Lim, W.F.; Mohamad Yusof, M.I.; Teh, L.K.; Salleh, M.Z. Significant decreased expressions of CaN, VEGF, SLC39A6 and SFRP1 in MDA-MB-231 xenograft breast tumor mice treated with Moringa oleifera leaves and seed residue (MOLSr) extracts. Nutrients 2020, 12, 2993. [Google Scholar] [CrossRef]
- He, H.; Liu, L.; Song, S.; Tang, X.; Wang, Y. Evaluation of glucosinolate composition and contents in Chinese Brassica vegetables. In Proceedings of the XXVI International Horticultural Congress: Asian Plants with Unique Horticultural Potential: Genetic Resources, Cultural 620, Toronto, ON, Canada, 1 December 2002; pp. 85–92. [Google Scholar]
- Tan, W.K.; Goenadie, V.; Lee, H.W.; Liang, X.; Loh, C.S.; Ong, C.N.; Tan, H.T.W. Growth and glucosinolate profiles of a common Asian green leafy vegetable, Brassica rapa subsp. chinensis var. parachinensis (choy sum), under LED lighting. Sci. Hortic. 2020, 261, 108922. [Google Scholar] [CrossRef]
- RDA-Genebank. RDA-Genebank Passport Data. Available online: http://genebank.rda.go.kr/ (accessed on 23 March 2023).
- Kim, S.-H.; Lee, G.-A.; Subramanian, P.; Hahn, B.-S. Quantification and Diversity Analyses of Major Glucosinolates in Conserved Chinese Cabbage (Brassica rapa L. ssp. pekinensis) Germplasms. Foods 2023, 12, 1243. [Google Scholar] [CrossRef]
- Tobias, S.; Carlson, J.E. Brief report: Bartlett’s test of sphericity and chance findings in factor analysis. Multivar. Behav. Res. 1969, 4, 375–377. [Google Scholar] [CrossRef]
- Kim, S.-H.; Subramanian, P.; Hahn, B.-S.; Ha, B.-K. High-Throughput Phenotypic Characterization and Diversity Analysis of Soybean Roots (Glycine max L.). Plants 2022, 11, 2017. [Google Scholar] [CrossRef]
- Kim, S.-H.; Jo, J.W.; Wang, X.; Shin, M.-J.; Hur, O.S.; Ha, B.-K.; Hahn, B.-S. Diversity characterization of soybean germplasm seeds using image analysis. Agronomy 2022, 12, 1004. [Google Scholar] [CrossRef]
- He, H.; Ping, L.; Bonnema, G.; Dekker, M.; Verkerk, R. Genetic variation in glucosinolate content within Brassica rapa vegetables. Acta Hortic. 2012, 944, 129–140. [Google Scholar]
- Wiesner, M.; Zrenner, R.; Krumbein, A.; Glatt, H.; Schreiner, M. Genotypic variation of the glucosinolate profile in pak choi (Brassica rapa ssp. chinensis). J. Agric. Food Chem. 2013, 61, 1943–1953. [Google Scholar] [CrossRef] [PubMed]
- Rhee, J.-H.; Choi, S.; Lee, J.-E.; Hur, O.-S.; Ro, N.-Y.; Hwang, A.-J.; Ko, H.-C.; Chung, Y.-J.; Noh, J.-J.; Assefa, A.D. Glucosinolate content in Brassica genetic resources and their distribution pattern within and between inner, middle, and outer leaves. Plants 2020, 9, 1421. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.; Hong, E.; Kim, G.H. Evaluation of antibacterial activity of 3-butenyl, 4-pentenyl, 2-phenylethyl, and benzyl isothiocyanate in Brassica vegetables. J. Food Sci. 2010, 75, M412–M416. [Google Scholar] [CrossRef]
- Yamada-Kato, T.; Nagai, M.; Ohnishi, M.; Yoshida, K. Inhibitory effects of wasabi isothiocyanates on chemical mediator release in RBL-2H3 rat basophilic leukemia cells. J. Nutr. Sci. Vitaminol. 2012, 58, 303–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, J.D.; Kelleher, M.O.; Eggleston, I.M. The cancer chemopreventive actions of phytochemicals derived from glucosinolates. Eur. J. Nutr. 2008, 47, 73–88. [Google Scholar] [CrossRef]
- Wang, J.; Gu, H.; Yu, H.; Zhao, Z.; Sheng, X.; Zhang, X. Genotypic variation of glucosinolates in broccoli (Brassica oleracea var. italica) florets from China. Food Chem. 2012, 133, 735–741. [Google Scholar] [CrossRef]
- Singh, R.R.; Reindl, K.M. Glutathione S-transferases in cancer. Antioxidants 2021, 10, 701. [Google Scholar] [CrossRef]
- Talalay, P. Chemoprotection against cancer by induction of phase 2 enzymes. Biofactors 2000, 12, 5–11. [Google Scholar] [CrossRef]
- Allain, E.P.; Rouleau, M.; Lévesque, E.; Guillemette, C. Emerging roles for UDP-glucuronosyltransferases in drug resistance and cancer progression. Br. J. Cancer 2020, 122, 1277–1287. [Google Scholar] [CrossRef] [Green Version]
- Soundararajan, P.; Kim, J.S. Anti-carcinogenic glucosinolates in cruciferous vegetables and their antagonistic effects on prevention of cancers. Molecules 2018, 23, 2983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Washida, K.; Miyata, M.; Koyama, T.; Yazawa, K.; Nomoto, K. Suppressive effect of Yamato-mana (Brassica rapa L. Oleifera Group) constituent 3-butenyl glucosinolate (gluconapin) on postprandial hypertriglyceridemia in mice. Biosci. Biotechnol. Biochem. 2010, 74, 1286–1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Hansen, P.E.; Wang, G.; Qiu, L.; Dong, J.; Yin, H.; Qian, Z.; Yang, M.; Miao, J. Pharmacological profile of xanthohumol, a prenylated flavonoid from hops (Humulus lupulus). Molecules 2015, 20, 754–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keck, A.-S.; Staack, R.; Jeffery, E.H. The cruciferous nitrile crambene has bioactivity similar to sulforaphane when administered to Fischer 344 rats but is far less potent in cell culture. Nutr. Cancer 2002, 42, 233–240. [Google Scholar] [CrossRef]
- Cao, Y.; Adhikari, S.; Clément, M.V.; Wallig, M.; Bhatia, M. Induction of apoptosis by crambene protects mice against acute pancreatitis via anti-inflammatory pathways. Am. J. Pathol. 2007, 170, 1521–1534. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Gao, X.; Chen, G.; Sun, J.; Chu, J.; Jing, K.; Li, P.; Zeng, R.; Wei, B. Indole-3-carbinol as inhibitors of glucocorticoid-induced apoptosis in osteoblastic cells through blocking ROS-mediated Nrf2 pathway. Biochem. Biophys. Res. Commun. 2015, 460, 422–427. [Google Scholar] [CrossRef]
- Traka, M.H.; Melchini, A.; Coode-Bate, J.; Al Kadhi, O.; Saha, S.; Defernez, M.; Troncoso-Rey, P.; Kibblewhite, H.; O’Neill, C.M.; Bernuzzi, F. Transcriptional changes in prostate of men on active surveillance after a 12-mo glucoraphanin-rich broccoli intervention—Results from the Effect of Sulforaphane on prostate CAncer PrEvention (ESCAPE) randomized controlled trial. Am. J. Clin. Nutr. 2019, 109, 1133–1144. [Google Scholar] [CrossRef] [Green Version]
- Jeffery, E.H.; Araya, M. Physiological effects of broccoli consumption. Phytochem. Rev. 2009, 8, 283–298. [Google Scholar] [CrossRef]
- Zasada, I.; Ferris, H. Sensitivity of Meloidogyne javanica and Tylenchulus semipenetrans to isothiocyanates in laboratory assays. Phytopathology 2003, 93, 747–750. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Talalay, P. Anticarcinogenic activities of organic isothiocyanates: Chemistry and mechanisms. Cancer Res. 1994, 54, 1976s–1981s. [Google Scholar]
- Staack, R.; Kingston, S.; Wallig, M.; Jeffery, E. A comparison of the individual and collective effects of four glucosinolate breakdown products from brussels sprouts on induction of detoxification enzymes. Toxicol. Appl. Pharmacol. 1998, 149, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Kelleher, M.O.; McMahon, M.; Eggleston, I.M.; Dixon, M.J.; Taguchi, K.; Yamamoto, M.; Hayes, J.D. 1-Cyano-2, 3-epithiopropane is a novel plant-derived chemopreventive agent which induces cytoprotective genes that afford resistance against the genotoxic α, β-unsaturated aldehyde acrolein. Carcinogenesis 2009, 30, 1754–1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoewsand, G. Bioactive organosulfur phytochemicals in Brassica oleracea vegetables—A review. Food Chem. Toxicol. 1995, 33, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.; Ho, H.-L.; Hsu, S.-J.; Chang, C.-C.; Tsai, M.-H.; Huo, T.-I.; Huang, H.-C.; Lee, F.-Y.; Hou, M.-C.; Lee, S.-D. Glucobrassicin metabolites ameliorate the development of portal hypertension and cirrhosis in bile duct-ligated rats. Int. J. Mol. Sci. 2019, 20, 4161. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; He, Q.; Yu, L.; Pham, Q.; Cheung, L.; Kim, Y.S.; Wang, T.T.; Smith, A.D. Indole-3-carbinol inhibits Citrobacter rodentium infection through multiple pathways including reduction of bacterial adhesion and enhancement of cytotoxic T cell activity. Nutrients 2020, 12, 917. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Malaysia | Thailand | Taiwan | Vietnam | China | Mauritius | India | Laos | Bangladesh | Total | |
|---|---|---|---|---|---|---|---|---|---|---|
| Cultivar | 2 | 1 | 1 | - | 1 | 1 | - | - | - | 6 |
| Landrace | 3 | 4 | 3 | 2 | 1 | 1 | 1 | 1 | 1 | 17 |
| Total | 5 | 5 | 4 | 2 | 2 | 2 | 1 | 1 | 1 | 23 |
| Name | Abbreviation | Class | RT (min) | MRM Transition | CID (ev) | Dwell Time (s) | Calibration Curve Parameters |
|---|---|---|---|---|---|---|---|
| Progoitrin | PRO | Aliphatic | 5.94 | 387.77 > 194.85 | 25 | 0.029 | Y = 8.2526X + 28.1501 (r2 = 0.961) |
| Sinigrin | SIN | Aliphatic | 6.56 | 357.75 > 161.84 | 25 | 0.029 | Y = 12.7878X − 11.1181 (r2 = 0.999) |
| Gluconapin | GNA | Aliphatic | 7.78 | 371.74 > 258.74 | 25 | 0.029 | Y = 8.36216X + 29.5397 (r2 = 0.994) |
| Glucoiberin | GIB | Aliphatic | 7.98 | 421.62 > 357.73 | 25 | 0.029 | Y = 33.6632X + 446.334 (r2 = 0.997) |
| Epiprogoitrin | EPI | Aliphatic | 8.06 | 387.7 > 258.74 | 25 | 0.029 | Y = 7.4939X − 6.76519 (r2 = 0.999) |
| Glucocheirolin | GCR | Aliphatic | 8.38 | 437.71 > 258.74 | 25 | 0.029 | Y = 20.7762X + 39.3608 (r2 = 0.986) |
| Glucoraphanin | GRA | Aliphatic | 8.39 | 435.59 > 177.78 | 25 | 0.029 | Y = 25.0808X + 60.584 (r2 = 0.983) |
| Glucoraphenin | GRE | Aliphatic | 8.53 | 433.66 > 258.81 | 25 | 0.029 | Y = 15.2565X + 3.62242 (r2 = 0.988) |
| Glucobrassicanapin | GBN | Aliphatic | 8.60 | 385.71 > 258.87 | 25 | 0.029 | Y = 7.2514X + 47.2841 (r2 = 0.992) |
| Glucobarbarin | GBB | Aromatic | 8.64 | 437.71 > 274.75 | 25 | 0.029 | Y = 9.29915X − 0.454779 (r2 = 0.999) |
| Glucoerucin | GER | Aliphatic | 8.73 | 419.69 > 258.74 | 25 | 0.029 | Y = 6.77393X + 73.6679 (r2 = 0.984) |
| Glucotropaeolin | GTL | Aromatic | 8.88 | 407.72 > 258.87 | 25 | 0.029 | Y = 18.2122X − 3.93949 (r2 = 0.999) |
| Sinalbin | SNB | Aromatic | 9.10 | 423.62 > 258.74 | 25 | 0.029 | Y = 49.7228X − 33.0636 (r2 = 0.999) |
| Glucoberteroin | GBE | Aliphatic | 9.18 | 433.72 > 275.06 | 25 | 0.029 | Y = 6.09397X + 63.1212 (r2 = 0.997) |
| Glucobrassicin | GBC | Indolyl | 9.31 | 446.69 > 204.94 | 25 | 0.029 | Y = 6.39827X + 2.6232 (r2 = 0.997) |
| Gluconasturtiin | GNS | Aromatic | 9.34 | 421.69 > 274.87 | 25 | 0.029 | Y = 4.36109X − 90.233 (r2 = 0.961) |
| Glucoraphasatin | GRH | Aromatic | 9.62 | 417.63 > 258.81 | 25 | 0.029 | Y = 15.5149X − 5.95281 (r2 = 0.997) |
| Variable | Range | Mean | Std. Deviation | |
|---|---|---|---|---|
| Aliphatic GSLs | Glucoiberin | 0~1.48 | 0.39 | 0.46 |
| Sinigrin | 0.16~17.69 | 3.69 | 4.21 | |
| Glucocheirolin | 0.08~19.91 | 5.48 | 6.07 | |
| Glucoerucin | 0.64~1983.01 | 227.83 | 562.36 | |
| Glucoraphanin | 2.29~569.16 | 166.77 | 179.00 | |
| Gluconapin | 117.38~13,111.41 | 2997.62 | 3406.77 | |
| Progoitrin | 120.20~3172.65 | 1430.06 | 899.82 | |
| Epiprogoitrin | 72.56~2728.20 | 1085.29 | 711.35 | |
| Glucoraphasatin | 0.03~9.89 | 0.70 | 2.02 | |
| Glucoraphenin | 0.11~9.18 | 1.35 | 2.10 | |
| Glucoberteroin | 6.02~3491.34 | 440.22 | 899.73 | |
| Glucobrassicanapin | 148.87~6830.64 | 1884.15 | 1457.35 | |
| Total aliphatic | 8243~18,110.85 | 8243.54 | 4557.95 | |
| Aromatic GSLs | Glucotropaeolin | 1.83~9.58 | 4.87 | 2.08 |
| Gluconasturtiin | 74.28~2379.24 | 631.28 | 575.41 | |
| Glucobarbarin | 0.97~8.04 | 2.94 | 1.71 | |
| Sinalbin | 0.04~2.96 | 0.34 | 0.69 | |
| Total aromatic | 84.98~2389.73 | 639.42 | 576.14 | |
| Indolic GSLs | Glucobrassicin | 85.15~908.09 | 333.30 | 203.01 |
| Total GSLs | 9216~20,023.79 | 9216.26 | 4905.73 | |
| Principal Component (Eigenvectors) | ||||
|---|---|---|---|---|
| PC1 | PC2 | PC3 | PC4 | |
| GIB | 0.3 | 0.11 | −0.27 | 0.19 |
| GCR | 0.25 | 0.25 | −0.21 | −0.22 |
| SIN | 0.09 | 0.39 | −0.29 | 0.2 |
| GER | 0.33 | −0.24 | 0.04 | −0.03 |
| GRA | 0.28 | 0.25 | −0.19 | −0.17 |
| GNA | 0.1 | 0.4 | −0.31 | 0.19 |
| GRH | 0.25 | −0.06 | 0.18 | −0.38 |
| PRO | 0.04 | 0.37 | 0.43 | −0.03 |
| EPI | 0.03 | 0.37 | 0.41 | −0.06 |
| GRE | 0.32 | −0.2 | 0.01 | 0.18 |
| GBE | 0.31 | −0.27 | 0.07 | 0.06 |
| GBN | −0.09 | 0.17 | 0.36 | 0.44 |
| GNL | −0.24 | −0.17 | −0.09 | 0.16 |
| GNS | 0.26 | 0.08 | 0.35 | 0.06 |
| GBB | 0.32 | −0.03 | 0.16 | 0.23 |
| SNB | 0.34 | −0.09 | 0.01 | −0.19 |
| GBC | 0.17 | −0.16 | −0.02 | 0.55 |
| Eigenvalue | 2.65 | 1.89 | 1.55 | 1.21 |
| Variability (%) | 41.50 | 23.13 | 14.11 | 8.62 |
| Cumulative (%) | 41.28 | 64.63 | 78.74 | 87.36 |
| Chemical Compounds | Class | Hydrolysis Products | Functions |
|---|---|---|---|
| Gluconapin | Aliphatic | 1-cyano-3,4-etithiobutane | In Mice |
| |||
| In Human | |||
| |||
| Glucobrassicanapin | Aliphatic | 4-pentenyl isothiocyanate | In Gram-negative bacteria |
| |||
| In rats [29] | |||
| |||
| Progoitrin | Aliphatic | Nitrile Crambene (1-cyano-2-hydroxy-3-butene) | In human Hep Gsub2 cell; mouse Hepa 1c1c7 cells and mouse H4IIEC3 cells [38] |
| |||
| In Swiss mice | |||
| In Swiss mice protect against acute pancreatitis by inducing pancreatic acinar cell apoptosis by activating anti-inflammatory and mitochondrial pathways [38,39] | |||
| Gluconasturtiin | Aromatic | 2-phenylethyl isothiocyante | The anticancer activity of phenyl ethyl isothiocyanate, a hydrolyzed product obtained from gluconasturtiin, is excellent as it induces cytoprotective genes mediated by Nrf2 and AhR transcription factors, represses NF-κB, and inhibits both cytochrome P450 and histone deacetylase [30] |
| Glucobrassicin | Indolic | Indole-3-carbinol | In human |
| In rat | |||
| |||
| In mice | |||
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.-H.; Subramanian, P.; Hahn, B.-S. Glucosinolate Diversity Analysis in Choy Sum (Brassica rapa subsp. chinensis var. parachinensis) Germplasms for Functional Food Breeding. Foods 2023, 12, 2400. https://doi.org/10.3390/foods12122400
Kim S-H, Subramanian P, Hahn B-S. Glucosinolate Diversity Analysis in Choy Sum (Brassica rapa subsp. chinensis var. parachinensis) Germplasms for Functional Food Breeding. Foods. 2023; 12(12):2400. https://doi.org/10.3390/foods12122400
Chicago/Turabian StyleKim, Seong-Hoon, Parthiban Subramanian, and Bum-Soo Hahn. 2023. "Glucosinolate Diversity Analysis in Choy Sum (Brassica rapa subsp. chinensis var. parachinensis) Germplasms for Functional Food Breeding" Foods 12, no. 12: 2400. https://doi.org/10.3390/foods12122400