
Analysis of Metabolic Components of JUNCAO Wine Based on GC-QTOF-MS
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Wine Samples
2.3. Untargeted Metabolomics Analysis Based on GC-QTOF-MS
2.4. Data Processing and Analysis
3. Results
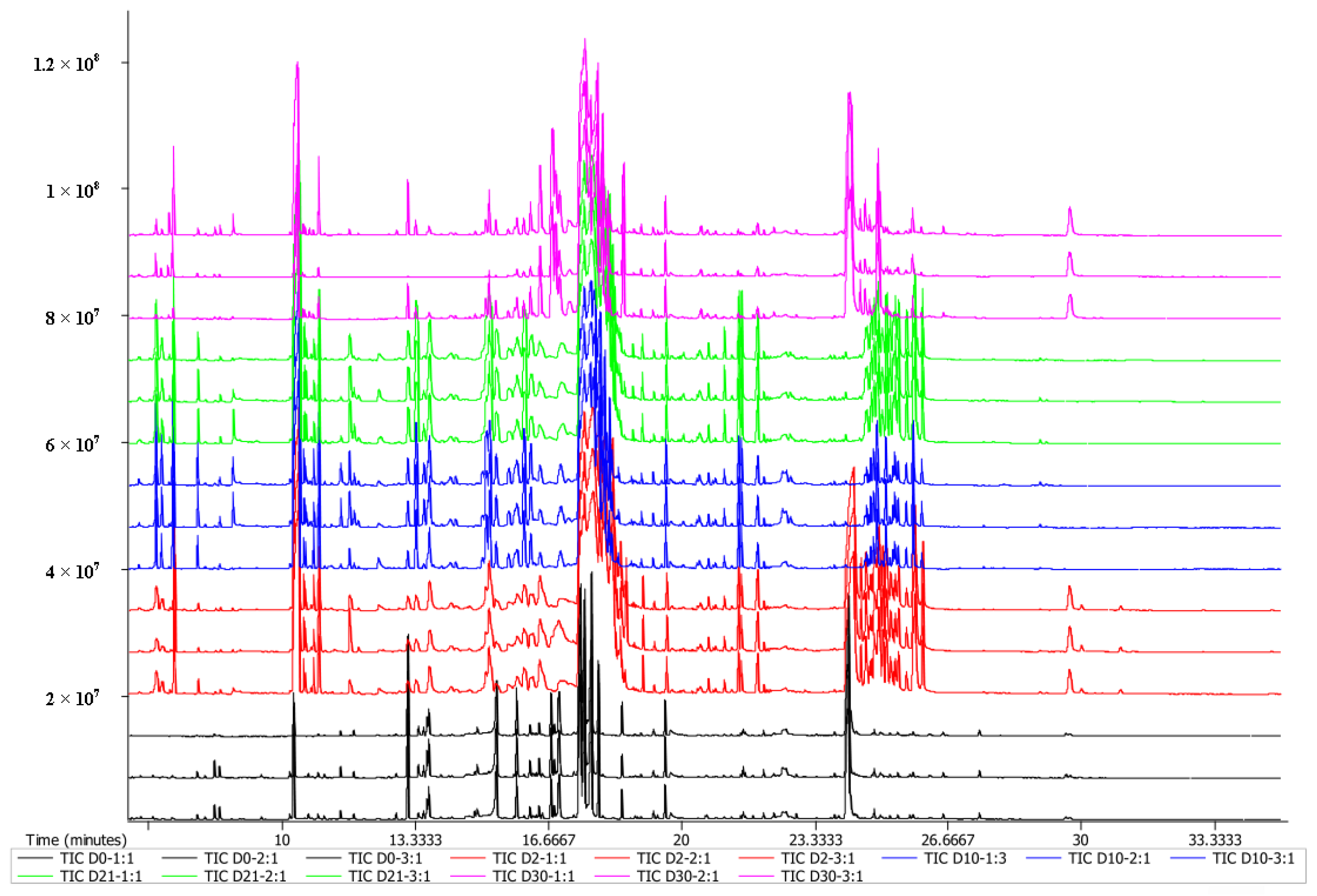
3.1. Sample Quality Control and Overall Analysis of Metabolites
3.2. PCA Analysis
3.3. VIP Diagram Analysis of OPLS-DA
3.4. Permutation Test
3.5. Heat Map Analysis of Different Compounds in the Fermentation Process of JUNCAO Wine
3.6. Metabolic Pathway Analysis of JUNCAO Wine during Fermentation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Meyer-Rochow, V.B. Can molecularly engineered plant galls help to ease the problem of world food shortage (and our de-pendence on pollinating insects). Foods 2022, 11, 4014. [Google Scholar] [CrossRef]
- Brankov, T.; Matkovski, B. Is a food shortage coming to the western Balkans. Foods 2022, 11, 3672. [Google Scholar] [CrossRef]
- Mehrabi, Z. How to buffer against an urban food shortage. Nature 2021, 595, 175–176. [Google Scholar] [CrossRef]
- Chen, X.H.; Zhang, X.; Liu, J.M.; Li, J.; Lin, B.S. High-throughput sequencing-based analysis of composition and growth-promoting effect of root-associated nitrogen-fixing bacteria in Pennisetum giganteum. J. Fujian Agric. For. Univ. 2022, 51, 570–576. [Google Scholar]
- Wei, D.D.; Wang, C.M.; Liu, J.F.; Zhao, X.L.; Wu, K.; Liang, C.Y.; Yang, B.; Zhang, W.D.; Yin, F. Study on production charac-teristics of biogas fermentation of giant Juncao in four growing environments. China Biogas 2022, 40, 29–36. [Google Scholar]
- Gustavo, C.; João, P.F.; Carla, V.; Maria, E.P.; João, J.S.; Isabel, V.F.; Maria, T.B. Polyphenols from Cymbopogon citratus leaves as topical anti-inflammory agents. J. Ethnopharmacol. 2016, 2, 222–228. [Google Scholar]
- Fan, J.L.; Zhang, L.L.; Zhang, Y.L.; Lin, H.; Yi, C.; Lin, Z.X. Optimization of brewing technology and quality analysis of JUNCAO wine. J. Fujian Agric. For. Univ. 2023, 52, 105. [Google Scholar]
- Wu, W.; Zhang, L.; Zheng, X.; Huang, Q.; Farag, M.A.; Zhu, R.; Zhao, C. Emerging applications of metabolomics in food science and future trends. Food Chem. X 2022, 16, 100500. [Google Scholar] [CrossRef]
- Chen, Y.; Wu, W.H.; Ni, X.; Farag, M.A.; Capanoglu, E.; Zhao, C. Regulatory mechanisms of the green alga Ulva lactuca oligosaccharide via the metabolomics and gut microbiome in diabetic mice. Curr. Res. Food Sci. 2022, 5, 1127–1139. [Google Scholar] [CrossRef]
- Xu, K.; Tian, C.Y.; Zhou, C.Z.; Zhu, C.; Weng, J.J.; Sun, Y.; Lin, Y.L.; Lai, Z.X.; Guo, Y.Q. Non-targeted metabolomics analysis revealed the characteristic non-volatile and volatile metabolites in the Rougui Wuyi rock tea (Camellia sinensis) from different culturing regions. Foods 2022, 11, 1694. [Google Scholar] [CrossRef] [PubMed]
- Li, N.Y.; Jiang, L.L.; Liu, Y.Y.; Zou, S.M.; Lu, M.; An, H.M. Metabolomics combined with transcriptomics analysis revealed the amino acids, phenolic acids, and flavonol derivatives biosynthesis network in developing Rosa roxburghii fruit. Foods 2022, 11, 1639. [Google Scholar] [CrossRef] [PubMed]
- Ling, X.W.; Yu, J.Q.; Peng, S.Y.; Chu, F.J. Combined application of intestinal microbial genomics metabolomic analysis, a review. Microbiol. China 2021, 48, 1292–1301. [Google Scholar]
- Onzo, A.; Acquavia, M.A.; Pascale, R.; Iannece, P.; Gaeta, C.; Lelario, F.; Ciriello, R.; Tesoro, C.; Bianco, G.; Capua, A.D. Untargeted metabolomic analysis by ultra-high-resolution mass spectrometry for the profiling of new Italian wine varieties. Anal. Bioanal. Chem. 2022, 414, 7805–7812. [Google Scholar] [CrossRef] [PubMed]
- Mayr, C.M.; De Rosso, M.; Dalla, V.A.; Flamini, R. High-resolution mass spectrometry identification of secondary metabolites in four red grape varieties potentially useful as traceability markers of wines. Beverages 2018, 4, 74. [Google Scholar] [CrossRef] [Green Version]
- Šukovíc, D.; Kneževíc, B.; Gašíc, U.; Sredojevíc, M.; Ćiríc, I.; Todíc, S.; Mutíc, J.; Tešíc, Ž. Phenolic profiles of leaves, grapes and wine of grapevine variety vranac (Vitis vinifera L.) from Montenegro. Foods 2020, 9, 138. [Google Scholar] [CrossRef] [Green Version]
- Baleiras-Couto, M.M.; Guedes, R.; Duarte, F.L.; Fortes, A.M.; Serralheiro, M. Untargeted metabolomics discriminates grapes and wines from two syrah vineyards located in the same wine region. Fermentation 2023, 9, 145. [Google Scholar] [CrossRef]
- Yan, H.G.; Zhang, J.Y.; Huo, Y.J.; Yuan, W. Volatile compounds analysis in blueberry wine fermented with different yeasts by gas chromatography-ion mobility spectrometry. Sci. Technol. Food Ind. 2021, 42, 234–241. [Google Scholar]
- O’Shea, K.; Misra, B.B. Software tools, databases and resources in metabolomics, updates from 2018 to 2019. Metabolomics 2020, 16, 36. [Google Scholar] [CrossRef]
- Wang, Z.X.; Chen, X.Y.; Liu, Q.Q.; Zhang, L.; Liu, S.; Su, Y.Y.; Ren, Y.M.; Yuan, C.L. Untargeted metabolomics analysis based on LC-IM-QTOF-MS for discriminating geographical origin and vintage of Chinese red wine. Food. Res. Int. 2023, 165, 112547. [Google Scholar] [CrossRef]
- Lopez, V.N.; Spelman, R.J.; Melis, J.; Davis, S.R.; Berry, S.D.; Lehnert, K.; Sneddon, N.W.; Holroyd, S.E.; Macgibbon, A.K.; Snell, R.G. Genetic correlations of milk fatty acid contents predicted from milk mid-infrared spectra in New Zealand dairy cattle. J. Dairy Sci. 2020, 103, 7238–7248. [Google Scholar] [CrossRef]
- Zeng, L.; Fu, Y.Q.; Huang, J.S.; Wang, J.R.; Jin, S.; Yin, J.F.; Xu, Y.Q. Comparative analysis of volatile compounds in Tieguanyin with different types based on HS–SPME–GC–MS. Foods 2022, 11, 1530. [Google Scholar] [CrossRef]
- Chen, M.; Gao, S.; Wang, R.Q.; Han, J.Q. Analysis of aroma components during cabernet Gernischt wine brewing process by HS-SPME-GC-MS. Food Ind. 2018, 39, 164–166. [Google Scholar]
- Zhu, J.; Hu, B.; Lu, J.; Xu, S. Analysis of metabolites in cabernet sauvignon and shiraz dry red wines from Shanxi by 1H NMR spectroscopy combined with pattern recognition analysis. Open Chem. 2018, 16, 446–452. [Google Scholar] [CrossRef] [Green Version]
- Aileen, P.; Huang, Y.L.; Vivian, G.R.M.; Kim-Huey, E.; Tan, L.P.; Maurin, C.; Liu, S.Q.; Benjamin, L.; Yu, B. Combination of solid phase microextraction and low energy electron ionisation gas chromatography-quadrupole time-of-flight mass spectrometry to meet the challenges of flavour analysis. Talanta 2021, 235, 122793. [Google Scholar]
- Yang, J.H.; Zhou, X.; Li, X.Q.; Li, X.M.; Gao, Y.; Zhang, Q.H. Simultaneous determination of 21 organophosphorus flame retardants in rice by gas chromatography quadrupole time-of-flight mass spectrometry. Talanta 2023, 253, 124101. [Google Scholar] [CrossRef]
- Chen, T.Y.; Wang, H.Y.; Su, W.; Mu, Y.C.; Tian, Y.X. Analysis of the formation mechanism of volatile and non-volatile flavor substances in corn wine fermentation based on high-throughput sequencing and metabolomics. Food Res. Int. 2023, 165, 112350. [Google Scholar] [CrossRef]
- Son, E.Y.; Lee, S.M.; Kim, M.; Seo, J.; Kim, Y. Comparison of volatile and non-volatile metabolites in rice wine fermented by Koji inoculated with Saccharomycopsis fibuligera and Aspergillus oryzae. Food Res. Int. 2018, 109, 596–605. [Google Scholar] [CrossRef]
- Han, D.M.; Chun, B.H.; Feng, T.; Kim, H.M.; Jeon, C.O. Dynamics of microbial communities and metabolites in ganjang, a traditional Korean fermented soy sauce, during fermentation bolites in ganjan. Food Microbiol. 2020, 92, 103591. [Google Scholar] [CrossRef]
- Chung, H.; Lee, N.; Seo, J.A.; Kim, Y.S. Comparative analysis of nonvolatile and volatile metabolites in Lichtheimia ramosa cultivated in different growth media. Biosci. Biotechnol. Biochem. 2017, 81, 565–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kind, T.; Wohlgemuth, G.; Lee, D.Y.; Lu, Y.; Palazoglu, M.; Shahbaz, S.; Fiehn, O. FiehnLib: Mass spectral and retention index libraries for metabolomics based on quadrupole and time-of-flight gas chromatography/mass spectrometry. Anal. Chem. 2009, 81, 10038–10048. [Google Scholar] [CrossRef] [Green Version]
- Kopka, J.; Schauer, N.; Krueger, S.; Birkemeyer, C.; Steinhauser, D. The Golm Metabolome Database. Bioinformatics 2005, 21, 1635–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasote, D.; Suleman, T.; Chen, W.; Sandasi, M.; Viljoen, A.; Vuuren, S. Chemical profiling and chemometric analysis of South African propolis. Biochem. Syst. Ecol. 2014, 55, 156–163. [Google Scholar] [CrossRef]
- Zhong, C.; Liu, L.P.; Li, Q.L.; Yang, P.F.; Hao, J.G.; Jia, S.R. Analyze the mechanism of flavor compounds formation using metabonomics method during industrial beer fermentation. China Biotechnol. 2016, 36, 49–58. [Google Scholar]
- Jiang, L.; Mu, Y.C.; Wei, S.; Mu, Y.; Zhao, C. Study on the dynamic changes and formation pathways of metabolites during the fermentation of black waxy rice wine. Food Sci. Nutr. 2020, 8, 2288–2298. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.B.; Yang, S.P. Effects of mannitol and sorbitol on salt tolerance of buckwheat seedlings. Hubei Agric. Sci. 2014, 53, 274–276. [Google Scholar]
- Wang, Y.f.; Chen, M.b.; Zhang, Y.; Li, Q.; Fang, S.L. Preliminary study on organic acids in Macheng old rice wine. Food Ind. 2020, 41, 199–202. [Google Scholar]
- Chun, A.R. Variation in quality and preference of sogokju (Korean traditional rice wine) from waxy rice varieties. Korean J. Crop Sci. 2010, 55, 177–186. [Google Scholar]
- Chen, Q.L.; Liu, S.P.; Tang, Y.F.; Han, X.; Zhou, Z.L.; Zou, H.J.; Wang, Z.M.; Ji, Z.W.; Mao, J. Changes in flavor components during fermentation process of mechanically produced Shaoxing rice wine. Food Sci. 2018, 39, 221–228. [Google Scholar]
- Wu, Z.W.; Sun, J.Y.; Wu, D.H.; Li, X.M.; Xie, G.F.; Lu, J. Preliminary study on organic acid and acid-producing bacterial during chinese rice wine fermentation. Food Ferment. Ind. 2016, 42, 12–18. [Google Scholar]
- Liu, X.; Liu, H.B.; Zeng, R.M.; Han, L.; Cai, N. Research Progress in Flavoring Characteristics of Dongjiu. Brew. Technol. 2016, 270, 91–93. [Google Scholar]
- Kosowski, G.; Mikulski, D.; Macko, D.; Miklaszewska, B.; Kotarska, K.; Czuprynski, B. Influence of various yeast strains and selected starchy raw materials on production of higher alcohols during the alcoholic fermentation process. Eur. Food. Res. Technol. 2015, 240, 233–242. [Google Scholar] [CrossRef]
- Wang, P.X.; Mao, J.; Meng, X.Y.; Li, Y.Y.; Feng, H. Changes in flavour characteristics and bacterial diversity during the traditional fermentation of chinese rice wines from Shaoxing region. Food Control 2014, 44, 58–63. [Google Scholar] [CrossRef]
- Kritzinger, E.C.; Bauer, F.F.; Toit, W.J. Role of glutathione in winemaking, a review. J. Agric. Food Chem. 2013, 61, 269–277. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Liu, L.L.; Jing, X.; Zhang, Y.F.; Xu, C.; Yan, Z.K.; Qi, Y.H. Analysis the difference of feng-flavour vintage base liquor based on the derivatization-GC-MS metabonomics method. Mod. Food Sci. Technol. 2021, 37, 295–303. [Google Scholar]
- Chen, J.; Kan, J.Q.; Zhang, R.; Tang, J.N. Study on fundamental components of honey-mulberry wine. Food Ferment. Ind. 2011, 37, 113–119. [Google Scholar]
- Zambrano, C.; Kotogan, A.; Bencsik, O.; Papp, T.; Vágvölgyi, C.; Keshab, C.M.; Krisch, J.; Takó, M. Mobilization of phenolic antioxidants from grape, apple and pitahaya residues via solid state fungal fermentation and carbohydrase treatment. LWT Food Sci. Technol. 2018, 89, 457–465. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.E.; Lee, S.; Jang, E.S.; Shin, W.; Moon, B.S.; Lee, C.H. Metabolomic profiles of Aspergillus oryzae and Bacillus amyloliq-uefaciens during rice koji fermentation. Molecules 2016, 21, 773. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, J.; Xiao, Z.; Qiu, W.; Zhao, C.; Yi, C.; Lin, D.; Lin, Z. Analysis of Metabolic Components of JUNCAO Wine Based on GC-QTOF-MS. Foods 2023, 12, 2254. https://doi.org/10.3390/foods12112254
Fan J, Xiao Z, Qiu W, Zhao C, Yi C, Lin D, Lin Z. Analysis of Metabolic Components of JUNCAO Wine Based on GC-QTOF-MS. Foods. 2023; 12(11):2254. https://doi.org/10.3390/foods12112254
Chicago/Turabian StyleFan, Jinlin, Zheng Xiao, Wanwei Qiu, Chao Zhao, Chao Yi, Dongmei Lin, and Zhanxi Lin. 2023. "Analysis of Metabolic Components of JUNCAO Wine Based on GC-QTOF-MS" Foods 12, no. 11: 2254. https://doi.org/10.3390/foods12112254