
Enzymatic Hydrolysis of Tenebrio molitor (Mealworm) Using Nuruk Extract Concentrate and an Evaluation of Its Nutritional, Functional, and Sensory Properties
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials, Chemicals, and Reagents
2.2. Preparation of Nuruk Extract Concentrate and Protease Activity
2.3. Enzymatic Hydrolysis of MW and Hydrolytic Properties of NEC
2.3.1. Degree of Hydrolysis
2.3.2. Total Soluble Solid and Hydrolysis Yield
2.3.3. Molecular Weight Distribution
2.4. Nutritional Composition of MW and Its Hydrolysates
2.4.1. Proximate and Total Amino Acid Composition
2.4.2. Free Amino Acid Analysis
2.5. Biological Activities of MW Hydrolysate
2.5.1. Antioxidant Activity
2.5.2. Angiotensin Converting Enzyme Inhibitory Activity
2.6. Sensory Properties of MW Hydrolysate Using E-Tongue
2.7. Statistical Analysis
3. Results and Discussion
3.1. Hydrolytic Properties of Nuruk Extract Concentrate
3.1.1. Protease Activity
3.1.2. Degree of Hydrolysis (DH), Hydrolysate Yield, and Total Soluble Solid
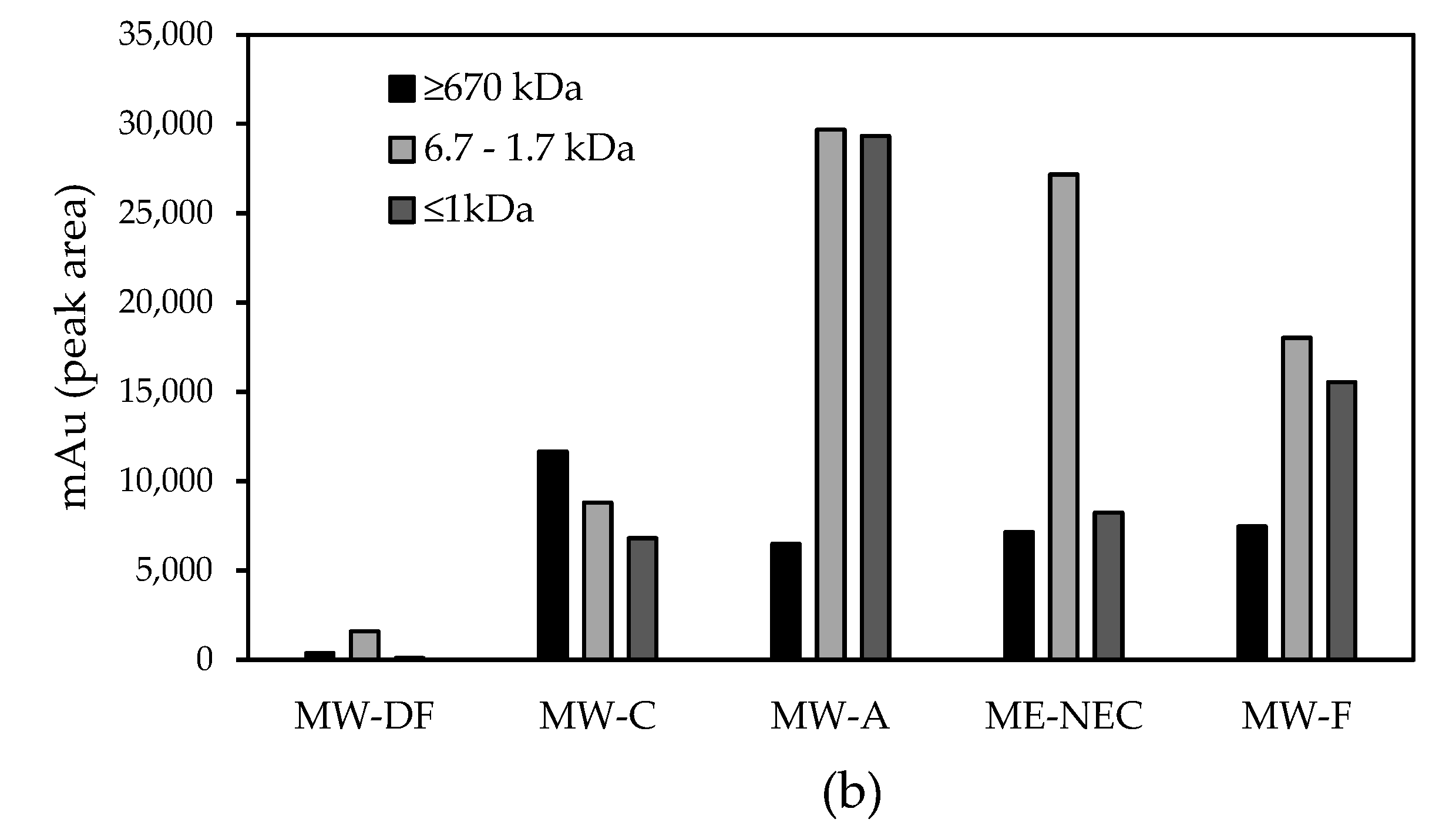
3.1.3. Molecular Weight Distribution
3.2. Nutritional Composition
3.2.1. Proximate and Total Amino Acid Composition of MW
3.2.2. Free Amino Acid Composition of MW Hydrolysate
3.3. Biological Activities of MW Hydrolysate
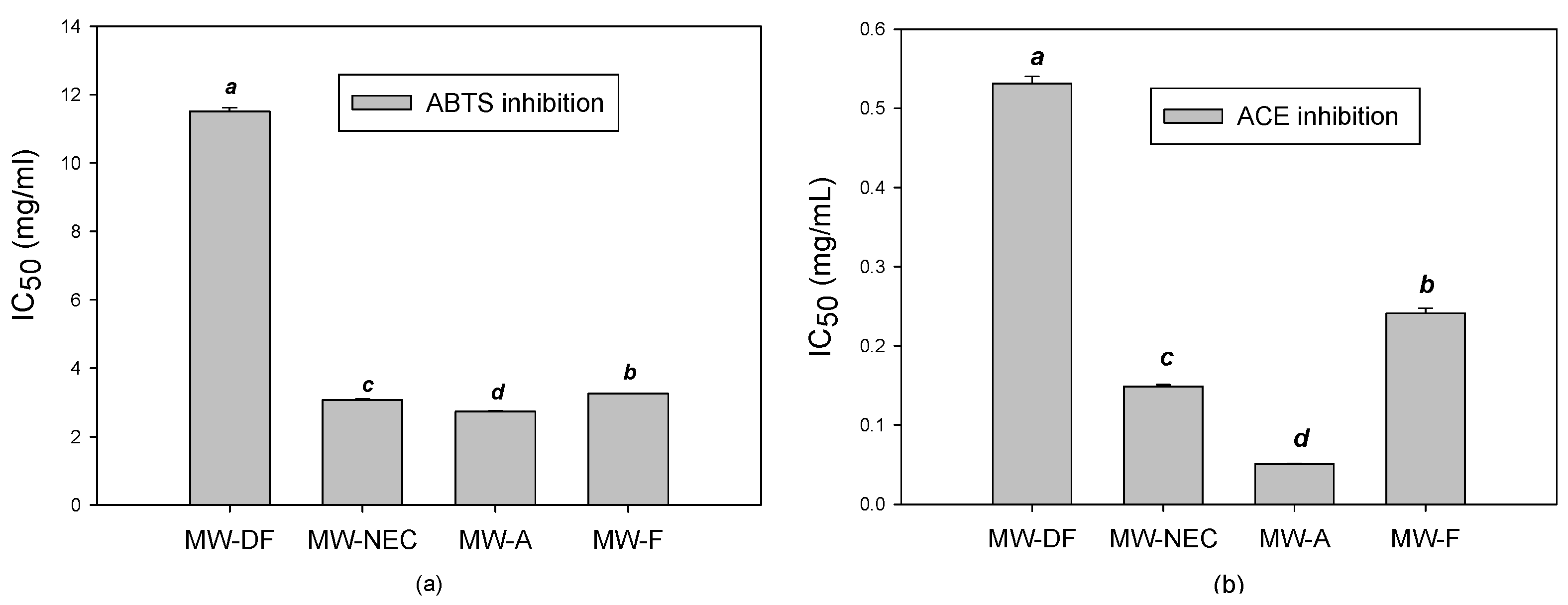
3.3.1. Antioxidant Activity
3.3.2. ACE Inhibitory Activity
3.4. Sensory Properties of MW Hydrolysate
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- FAO. Sustainable and Circular Bioeconomy for Food Systems Transformation. Alternative Proteins Top the Bill for the Latest FAO–International Sustainable Bioeconomy. 2022. Available online: https://www.fao.org/in-action/sustainable-and-circular-bioeconomy/resources/news/details/en/c/1507553/ (accessed on 5 December 2022).
- Yoon, S.; Wong, N.A.K.; Chae, M.; Auh, J.-H. Comparative Characterization of Protein Hydrolysates from Three Edible Insects: Mealworm Larvae, Adult Crickets, and Silkworm Pupae. Foods 2019, 8, 563. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Vázquez-Gutiérrez, J.L.; Johansson, D.; Landberg, R.; Langton, M. Yellow Mealworm Protein for Food Purposes—Extraction and Functional Properties. PLoS ONE 2016, 11, e0147791. [Google Scholar] [CrossRef] [PubMed]
- Bordiean, A.; Krzyżaniak, M.; Stolarski, M.J.; Czachorowski, S.; Peni, D. Will Yellow Mealworm Become a Source of Safe Proteins for Europe? Agriculture 2020, 10, 233. [Google Scholar] [CrossRef]
- Zielińska, E.; Baraniak, B.; Karaś, M.; Rybczyńska-Tkaczyk, K.; Jakubczyk, A. Selected species of edible insects as a source of nutrient composition. Food Res. Int. 2015, 77, 460–466. [Google Scholar] [CrossRef]
- Borremans, A.; Bußler, S.; Sagu, S.T.; Rawel, H.; Schlüter, O.K.; Leen, V.C. Effect of Blanching Plus Fermentation on Selected Functional Properties of Mealworm (Tenebrio molitor) Powders. Foods 2020, 9, 917. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.-J.; Choi, S.Y.; Hwang, I.-K.; Nho, C.W.; Kim, S.H. Could Defatted Mealworm (Tenebrio molitor) and Mealworm Oil Be Used as Food Ingredients? Foods 2020, 9, 40. [Google Scholar] [CrossRef]
- Janssen, R.H.; Vincken, J.-P.; van den Broek, L.A.M.; Fogliano, V.; Lakemond, C.M.M. Nitrogen-to-Protein Conversion Factors for Three Edible Insects: Tenebrio molitor, Alphitobius diaperinus, and Hermetia illucens. J. Agric. Food Chem. 2017, 65, 2275–2278. [Google Scholar] [CrossRef]
- Muthukrishnan, S.; Mun, S.; Noh, M.Y.; Geisbrecht, E.R.; Arakane, Y. Insect Cuticular Chitin Contributes to Form and Function. Curr. Pharm. Des. 2020, 26, 3530–3545. [Google Scholar] [CrossRef]
- Segura-Campos, M.; Espinosa-García, L.; Chel-Guerrero, L.A.; Betancur-Ancona, D.A. Effect of Enzymatic Hydrolysis on Solubility, Hydrophobicity, and In Vivo Digestibility in Cowpea (Vigna unguiculata). Int. J. Food Prop. 2012, 15, 770–780. [Google Scholar] [CrossRef]
- Purschke, B.; Meinlschmidt, P.; Horn, C.; Rieder, O.; Jäger, H. Improvement of techno-functional properties of edible insect protein from migratory locust by enzymatic hydrolysis. Eur. Food Res. Technol. 2018, 244, 999–1013. [Google Scholar] [CrossRef]
- Tapal, A.; Tiku, P.K. Nutritional and nutraceutical improvement by enzymatic modification of food proteins. In Enzymes in Food Biotechnology: Production, Applications, and Future Prospects; Elsevier Inc.: Amsterdam, The Netherlands, 2019; pp. 471–481. [Google Scholar] [CrossRef]
- Hall, F.G.; Jones, O.G.; O’Haire, M.E.; Liceaga, A.M. Functional properties of tropical banded cricket (Gryllodes sigillatus) protein hydrolysates. Food Chem. 2017, 224, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Nongonierma, A.B.; FitzGerald, R.J. Unlocking the biological potential of proteins from edible insects through enzymatic hydrolysis: A review. Innov. Food Sci. Emerg. Technol. 2017, 43, 239–252. [Google Scholar] [CrossRef]
- Sousa, P.; Borges, S.; Pintado, M. Enzymatic hydrolysis of insect Alphitobius diaperinus towards the development of bioactive peptide hydrolysates. Food Funct. 2020, 11, 3539–3548. [Google Scholar] [CrossRef] [PubMed]
- Leni, G.; Soetemans, L.; Jacobs, J.; Depraetere, S.; Gianotten, N.; Bastiaens, L.; Caligiani, A.; Sforza, S. Protein hydrolysates from Alphitobius diaperinus and Hermetia illucens larvae treated with commercial proteases. J. Insects Food Feed. 2020, 6, 393–404. [Google Scholar] [CrossRef]
- Su, G.; Ren, J.; Yang, B.; Cui, C.; Zhao, M. Comparison of hydrolysis characteristics on defatted peanut meal proteins between a protease extract from Aspergillus oryzae and commercial proteases. Food Chem. 2011, 126, 1306–1311. [Google Scholar] [CrossRef]
- Tang, Y.; Debnath, T.; Choi, E.-J.; Kim, Y.W.; Ryu, J.P.; Jang, S.; Chung, S.; Choi, Y.J.; Kim, E.-K. Changes in amino acid profile and free radical scavenging activties of Tenebrio molitor larvae following enzymatic hydrolysis. PLoS ONE 2018, 13, e0196218. [Google Scholar] [CrossRef]
- Hong, H.H.; Kim, M.K. Physiochemical Quality and Sensory Characteristics of koji Made with Soybean, Rice, and Wheat for Commercial doenjang Production. Foods 2020, 9, 975. [Google Scholar] [CrossRef]
- Lee, J.-E.; Lee, A.R.; Kim, H.; Lee, E.; Kim, T.W.; Shin, W.C.; Kim, J.H. Restoration of Traditional Korean Nuruk and Analysis of the Brewing Characteristics. J. Microbiol. Biotechnol. 2017, 27, 896–908. [Google Scholar] [CrossRef]
- Bal, J.; Yun, S.-H.; Song, H.-Y.; Yeo, S.-H.; Kim, J.H.; Kim, J.-M.; Kim, D.-H. Mycoflora dynamics analysis of Korean traditional wheat-based nuruk. J. Microbiol. 2014, 52, 1025–1029. [Google Scholar] [CrossRef]
- Hoang, N.X.; Ting, C.-H.; Hsu, C.-K. Optimization of Protease and Amylase Activities Derived from Soy Sauce Koji. J. Food Sci. Nutr. Res. 2020, 3, 153–170. [Google Scholar]
- Yu, K.-W.; Lee, S.-E.; Choi, H.-S.; Suh, H.J.; Ra, K.S.; Choi, J.W.; Hwang, J.-H. Optimization for rice koji preparation using aspergillus oryzae CJCM-4 isolated from a korean traditional meju. Food Sci. Biotechnol. 2012, 21, 129–135. [Google Scholar] [CrossRef]
- Chancharoonpong, C.; Hsieh, P.-C.; Sheu, S.-C. Enzyme Production and Growth of Aspergillus oryzae S. on Soybean Koji Fermentation. APCBEE Procedia 2012, 2, 57–61. [Google Scholar] [CrossRef]
- Bechman, A.; Phillips, R.D.; Chen, J. Changes in Selected Physical Property and Enzyme Activity of Rice and Barley Koji during Fermentation and Storage. J. Food Sci. 2012, 77, M318–M322. [Google Scholar] [CrossRef]
- Spellman, D.; McEvoy, E.; O’cuinn, G.; FitzGerald, R. Proteinase and exopeptidase hydrolysis of whey protein: Comparison of the TNBS, OPA and pH stat methods for quantification of degree of hydrolysis. Int. Dairy J. 2003, 13, 447–453. [Google Scholar] [CrossRef]
- AOAC. Official Method of Analysis, 18th ed.; Association of Official Analytical Chemist: Washington, DC, USA, 2005. [Google Scholar]
- Waters Corporations. UPLC Amino Acid Analysis Solution System Guide; 71500129702/Revision B; Waters Corporation: Milford, MA, USA, 2007. [Google Scholar]
- Wang, M.; Li, C.; Li, H.; Wu, Z.; Chen, B.; Lei, Y.; Shen, Y. In Vitro and In Silico Antioxidant Activity of Novel Peptides Prepared from Paeonia Ostii ‘Feng Dan’ Hydrolysate. Antioxidants 2019, 8, 433. [Google Scholar] [CrossRef]
- Wang, Z.; Li, J.; Hong, X.; Han, X.; Liu, B.; Li, X.; Zhang, H.; Gao, J.; Liu, N.; Gao, X.; et al. Taste Masking Study Based on an Electronic Tongue: The Formulation Design of 3D Printed Levetiracetam Instant-Dissolving Tablets. Pharm. Res. 2021, 38, 831–842. [Google Scholar] [CrossRef]
- Alpha, M.O.S. Astree E-Tongue Hardware User Manual; Alpha MOS: Toulouse, France, 2020; pp. 47–90. [Google Scholar]
- Bal, J.; Yun, S.-H.; Yeo, S.-H.; Kim, J.-M.; Kim, D.-H. Metagenomic analysis of fungal diversity in Korean traditional wheat-based fermentation starter nuruk. Food Microbiol. 2016, 60, 73–83. [Google Scholar] [CrossRef]
- Carroll, E.; Trinh, T.N.; Son, H.; Lee, Y.-W.; Seo, J.-A. Comprehensive analysis of fungal diversity and enzyme activity in nuruk, a Korean fermenting starter, for acquiring useful fungi. J. Microbiol. 2017, 55, 357–365. [Google Scholar] [CrossRef]
- Charles, C.W.; Von, W.; Andrew, W.N. Introduction to Enzymes; Worthington Bichemical Corporation: Washington, DC, USA, 2019. [Google Scholar]
- Hunsakul, K.; Laokuldilok, T.; Sakdatorn, V.; Klangpetch, W.; Brennan, C.S.; Utama-Ang, N. Optimization of enzymatic hydrolysis by alcalase and flavourzyme to enhance the antioxidant properties of jasmine rice bran protein hydrolysate. Sci. Rep. 2022, 12, 12582. [Google Scholar] [CrossRef]
- Leni, G.; Soetemans, L.; Caligiani, A.; Sforza, S.; Bastiaens, L. Degree of Hydrolysis Affects the Techno-Functional Properties of Lesser Mealworm Protein Hydrolysates. Foods 2020, 9, 381. [Google Scholar] [CrossRef]
- Yu, M.-H.; Lee, H.-S.; Cho, H.-R.; Lee, S.-O. Enzymatic Preparation and Antioxidant Activities of Protein Hydrolysates from Tenebrio molitor Larvae (Mealworm). J. Korean Soc. Food Sci. Nutr. 2017, 46, 435–441. [Google Scholar] [CrossRef]
- Chen, F.; Jiang, H.; Lu, Y.; Chen, W.; Huang, G. Identification and in silico analysis of antithrombotic peptides from the enzymatic hydrolysates of Tenebrio molitor larvae. Eur. Food Res. Technol. 2019, 245, 2687–2695. [Google Scholar] [CrossRef]
- Dai, C.; Ma, H.; Luo, L.; Yin, X. Angiotensin I-converting enzyme (ACE) inhibitory peptide derived from Tenebrio molitor (L.) larva protein hydrolysate. Eur. Food Res. Technol. 2013, 236, 681–689. [Google Scholar] [CrossRef]
- Mirzapour-Kouhdasht, A.; Garcia-Vaquero, M.; Eun, J.-B.; Simal-Gandara, J. Influence of Enzymatic Hydrolysis and Molecular Weight Fractionation on the Antioxidant and Lipase/α-Amylase Inhibitory Activities In Vitro of Watermelon Seed Protein Hydrolysates. Molecules 2022, 27, 7897. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.-R.; Lee, S.-O. Novel hepatoprotective peptides derived from protein hydrolysates of mealworm (Tenebrio molitor). Food Res. Int. 2020, 133, 109194. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.-J.; Lee, J.-C.; Hwang, I.-K.; Nho, C.W.; Kim, S.-H. Physicochemical properties of mealworm (Tenebrio molitor) powders manufactured by different industrial processes. LWT 2019, 116, 108514. [Google Scholar] [CrossRef]
- Kröncke, N.; Böschen, V.; Woyzichovski, J.; Demtröder, S.; Benning, R. Comparison of suitable drying processes for mealworms (Tenebrio molitor). Innov. Food Sci. Emerg. Technol. 2018, 50, 20–25. [Google Scholar] [CrossRef]
- Azagoh, C.; Ducept, F.; Garcia, R.; Rakotozafy, L.; Cuvelier, M.-E.; Keller, S.; Lewandowski, R.; Mezdour, S. Extraction and physicochemical characterization of Tenebrio molitor proteins. Food Res. Int. 2016, 88, 24–31. [Google Scholar] [CrossRef]
- Ghosh, S.; Lee, S.; Jung, C.; Meyer-Rochow, V.B. Nutritional composition of five commercial edible insects in South Korea. J. Asia-Pac. Entomol. 2017, 20, 686–694. [Google Scholar] [CrossRef]
- Koopman, R.; Crombach, N.; Gijsen, A.P.; Walrand, S.; Fauquant, J.; Kies, A.K.; Lemosquet, S.; Saris, W.H.M.; Boirie, Y.; van Loon, L.J.C. Ingestion of a protein hydrolysate is accompanied by an accelerated in vivo digestion and absorption rate when compared with its intact protein. Am. J. Clin. Nutr. 2009, 90, 106–115. [Google Scholar] [CrossRef]
- Esfandi, R.; Walters, M.E.; Tsopmo, A. Antioxidant properties and potential mechanisms of hydrolyzed proteins and peptides from cereals. Heliyon 2019, 5, e01538. [Google Scholar] [CrossRef] [PubMed]
- Mine, Y.; Li-Chan, E.; Jiang, B. Bioactive Protein and Peptides as a Functional Foods and Nutraciticals; Ames, I.A., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2010; 420p. [Google Scholar] [CrossRef]
- Vercruysse, L.; Smagghe, G.; Herregods, G.; Van Camp, J. ACE Inhibitory Activity in Enzymatic Hydrolysates of Insect Protein. J. Agric. Food Chem. 2005, 53, 5207–5211. [Google Scholar] [CrossRef] [PubMed]
- Hall, F.; Johnson, P.E.; Liceaga, A. Effect of enzymatic hydrolysis on bioactive properties and allergenicity of cricket (Gryllodes sigillatus) protein. Food Chem. 2017, 262, 39–47. [Google Scholar] [CrossRef]
- Badal, C.S.; Hayashi, K. Debittering of protein hydrolysates. Biotechnol. Adv. 2001, 19, 355–370. [Google Scholar]
- Kang, J.-E.; Kim, C.-W.; Yeo, S.-H.; Jeong, S.-T.; Kim, Y.-S.; Choi, H.-S. Effect of heat-treated Nuruk on the quality characteristics of aged Yakju. Food Sci. Biotechnol. 2018, 27, 715–724. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Choi, S.J.; Kwak, J.; Kim, K.; Seo, M.; Moon, T.W.; Lee, Y.-W. Aspergillus oryzae strains isolated from traditional Korean Nuruk: Fermentation properties and influence on rice wine quality. Food Sci. Biotechnol. 2013, 22, 425–432. [Google Scholar] [CrossRef]
- Minagawa, E.; Kaminokawa, S.; Tsukasaki, F.; Yamauchi, K. Debittering mechanism in bitter peptides of enzymatic hydrolysates from milk casein by aminopeptidase. J. Food Sci. 1989, 54, 1225–1229. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Description | E/S Ratio (w/w) |
|---|---|---|
| MW-C | Mealworm control | No enzyme |
| MW-NEC | Mealworm hydrolyzed by nuruk extract concentrate | 10/100 |
| MW-F | Mealworm hydrolyzed by flavourzyme | 1/100 |
| MW-A | Mealworm hydrolyzed by alcalase | 1/100 |
| Enzyme | Protease Activity (EU/mL) | Protease Activity (EU/g) |
|---|---|---|
| Nuruk crude extract | 6.78 ± 0.07 c | 67.84 ± 0.73 d |
| Nuruk extract concentrate (>30 kDa) | 12.71 ± 0.33 a | 127.08 ± 3.30 c |
| Alcalase 2.4 L (1%) | 11.07 ± 0.07 b | 1107.27 ± 7.60 b |
| Flavourzyme 1000 L (1%) | 12.45 ± 0.02 a | 1245.37 ± 2.90 a |
| Product | DH (%) | TSS (%) | Yield (%) |
|---|---|---|---|
| MW-C | 0.51 ± 0.01 d | 3.60 ± 0.00 d | 18.62 ± 0.68 d |
| MW-NEC | 15.10 ± 0.08 a | 5.97 ± 0.06 b | 35.92 ± 0.78 b |
| MW-A | 9.56 ± 0.22 c | 7.97 ± 0.06 a | 42.88 ± 1.01 a |
| MW-F | 11.03 ± 0.46 b | 5.70 ± 0.10 c | 28.99 ± 0.71 c |
| Parameter | MW-Whole | MW-Defatted |
|---|---|---|
| Crude protein | 49.16 ± 0.17 b | 71.41 ± 0.09 a |
| Crude fat | 27.81 ± 0.73 a | 8.47 ± 0.02 b |
| Moisture | 3.41 ± 0.03 a | 0.29 ± 0.03 b |
| Ash | 3.09 ± 0.03 b | 4.63 ± 0.01 a |
| Carbohydrate | 16.52 ± 0.90 a | 15.19 ± 0.08 a |
| Free amino acid | 0.55 ± 0.01 b | 0.64 ± 0.03 a |
| Total amino acid | 39.62 ± 0.23 b | 53.48 ± 0.15 a |
| Amino Acids | MW-DF | MW-NEC | MW-A | MW-F |
|---|---|---|---|---|
| His | 0.47 ± 0.01 d | 3.42 ± 0.09 b | 1.86 ± 0.07 c | 3.35 ± 0.21 a |
| Thr | 0.05 ± 0.00 d | 1.12 ± 0.02 b | 0.49 ± 0.02 c | 2.28 ± 0.11 a |
| Lys | 0.19 ± 0.02 d | 3.35 ± 0.13 b | 1.36 ± 0.10 c | 5.49 ± 0.05 a |
| Met | 0.18 ± 0.00 b | 0.57 ± 0.01 a | 0.57 ± 0.02 a | 0.60 ± 0.05 a |
| Val | 0.39 ± 0.02 d | 4.55 ± 0.10 b | 2.56 ± 0.11 c | 5.58 ± 0.19 a |
| Ilu | 0.19 ± 0.01 d | 2.60 ± 0.07 b | 1.09 ± 0.06 c | 3.26 ± 0.10 a |
| Leu | 0.10 ± 0.01 d | 3.63 ± 0.10 b | 1.06 ± 0.05 c | 6.23 ± 0.18 a |
| Phe | 0.09 ± 0.00 d | 1.63 ± 0.04 b | 0.48 ± 0.02 c | 2.18 ± 0.13 a |
| EAA | 1.66 ± 0.08 d | 20.87 ± 0.54 b | 9.48 ±0.40 c | 28.97 ± 1.01 a |
| Ser | 0.57 ± 0.02 c | 1.09 ± 0.02 b | 0.33 ± 0.02 d | 2.15 ± 0.10 a |
| Arg | 0.26± 0.01 d | 6.37 ± 0.14 b | 3.30 ± 0.11 c | 7.76 ± 0.40 a |
| Gly | 0.08 ±0.00 d | 1.19 ± 0.03 b | 0.57 ± 0.03 c | 1.40 ± 0.07 a |
| Asp | 0.30 ± 0.00 d | 1.49 ± 0.04 a | 0.93 ± 0.03 b | 0.78 ± 0.03 c |
| Glu | 0.02 ± 0.01 d | 29.02 ± 1.20 a | 21.22 ± 1.62 b | 13.87 ± 0.52 c |
| Ala | 0.40 ± 0.03 c | 4.84 ± 0.13 a | 2.01 ± 0.09 b | 4.97 ± 0.12 a |
| Pro | 2.30 ± 0.11 d | 18.53 ± 0.47 a | 10.74 ± 0.57 d | 12.77 ± 0.41 b |
| Cys | 0.09 ± 0.01 d | 0.33 ± 0.02 c | 0.57 ± 0.02 a | 0.40 ± 0.01 b |
| Tyr | 0.75 ± 0.02 c | 6.62 ± 0.18 a | 3.86 ± 0.15 b | 6.57 ± 0.39 a |
| NEAA | 4.78 ± 0.21 d | 69.50 ± 1.01 a | 43.53 ± 0.57 c | 50.67 ± 2.01 b |
| Total AA | 6.44 ± 0.28 d | 90.37 ± 1.22 a | 53.01 ± 0.93 c | 79.64 ± 3.02 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chewaka, L.S.; Park, C.S.; Cha, Y.-S.; Desta, K.T.; Park, B.-R. Enzymatic Hydrolysis of Tenebrio molitor (Mealworm) Using Nuruk Extract Concentrate and an Evaluation of Its Nutritional, Functional, and Sensory Properties. Foods 2023, 12, 2188. https://doi.org/10.3390/foods12112188
Chewaka LS, Park CS, Cha Y-S, Desta KT, Park B-R. Enzymatic Hydrolysis of Tenebrio molitor (Mealworm) Using Nuruk Extract Concentrate and an Evaluation of Its Nutritional, Functional, and Sensory Properties. Foods. 2023; 12(11):2188. https://doi.org/10.3390/foods12112188
Chicago/Turabian StyleChewaka, Legesse Shiferaw, Chan Soon Park, Youn-Soo Cha, Kebede Taye Desta, and Bo-Ram Park. 2023. "Enzymatic Hydrolysis of Tenebrio molitor (Mealworm) Using Nuruk Extract Concentrate and an Evaluation of Its Nutritional, Functional, and Sensory Properties" Foods 12, no. 11: 2188. https://doi.org/10.3390/foods12112188