
Uncovering the Nutritive Profiles of Adult Male Chinese Mitten Crab (E. sinensis) Harvested from the Pond and Natural Water Area of Qin Lake Based on Metabolomics


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation and Sampling
2.2. Proximate Composition Analysis
2.3. Amino Acid Determination
2.4. Fatty Acid Composition
2.5. Wide Targeted Metabolomics Analysis
2.6. Statistical Analysis
3. Results
3.1. Proximate Composition of the E. sinensis
3.2. Amino Acid Determination
3.3. Fatty Acid Composition
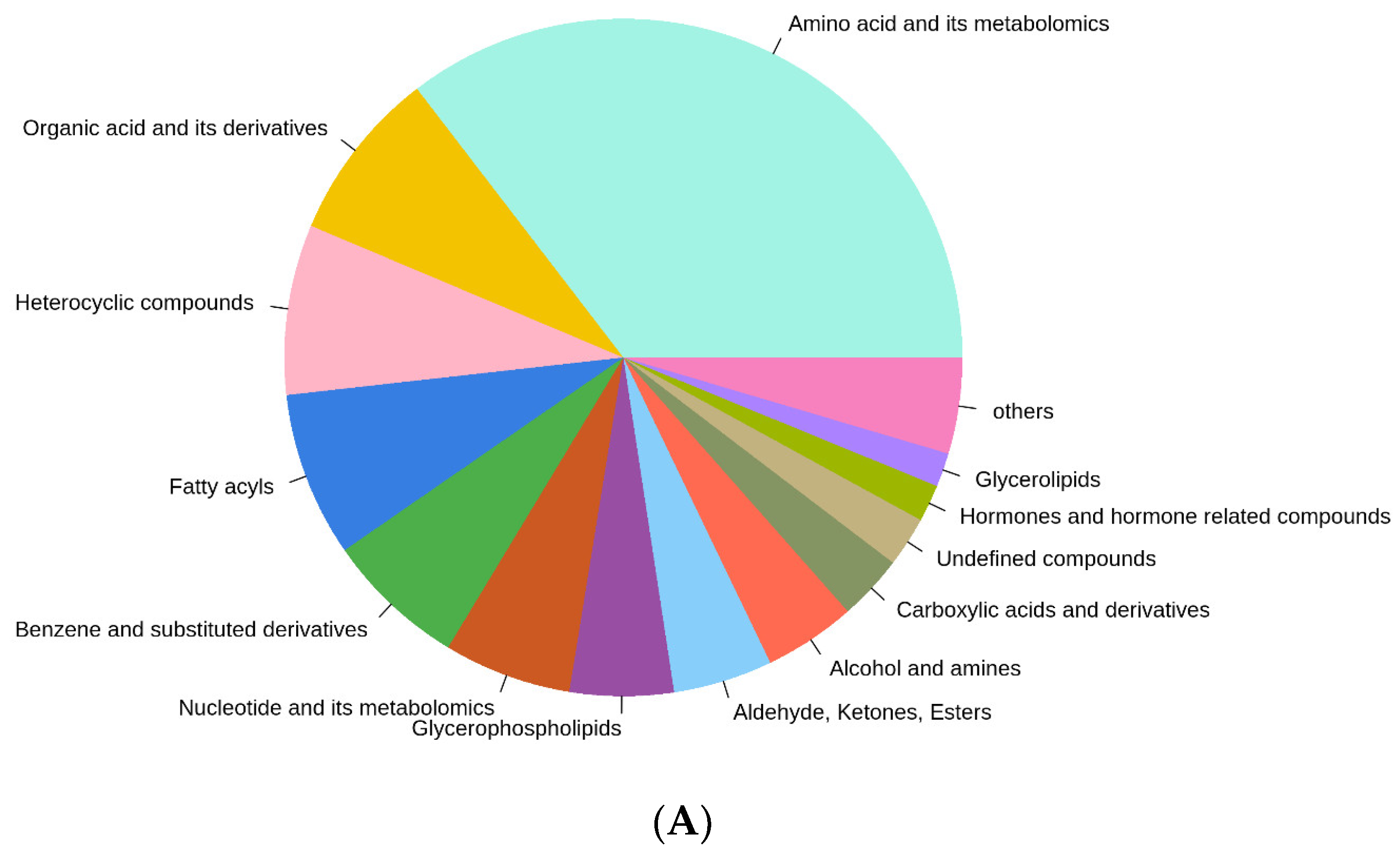
3.4. Overview of the Metabolic Profile of E. sinensis
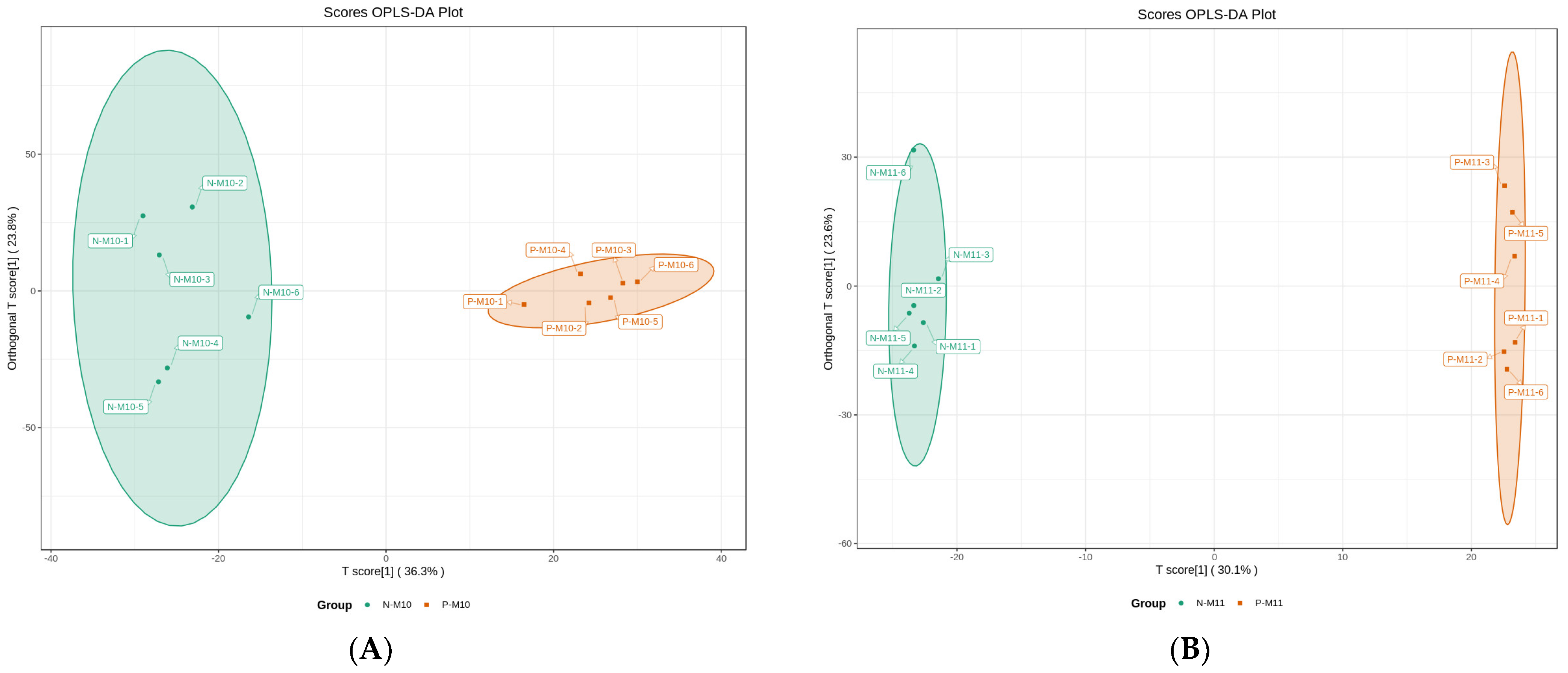
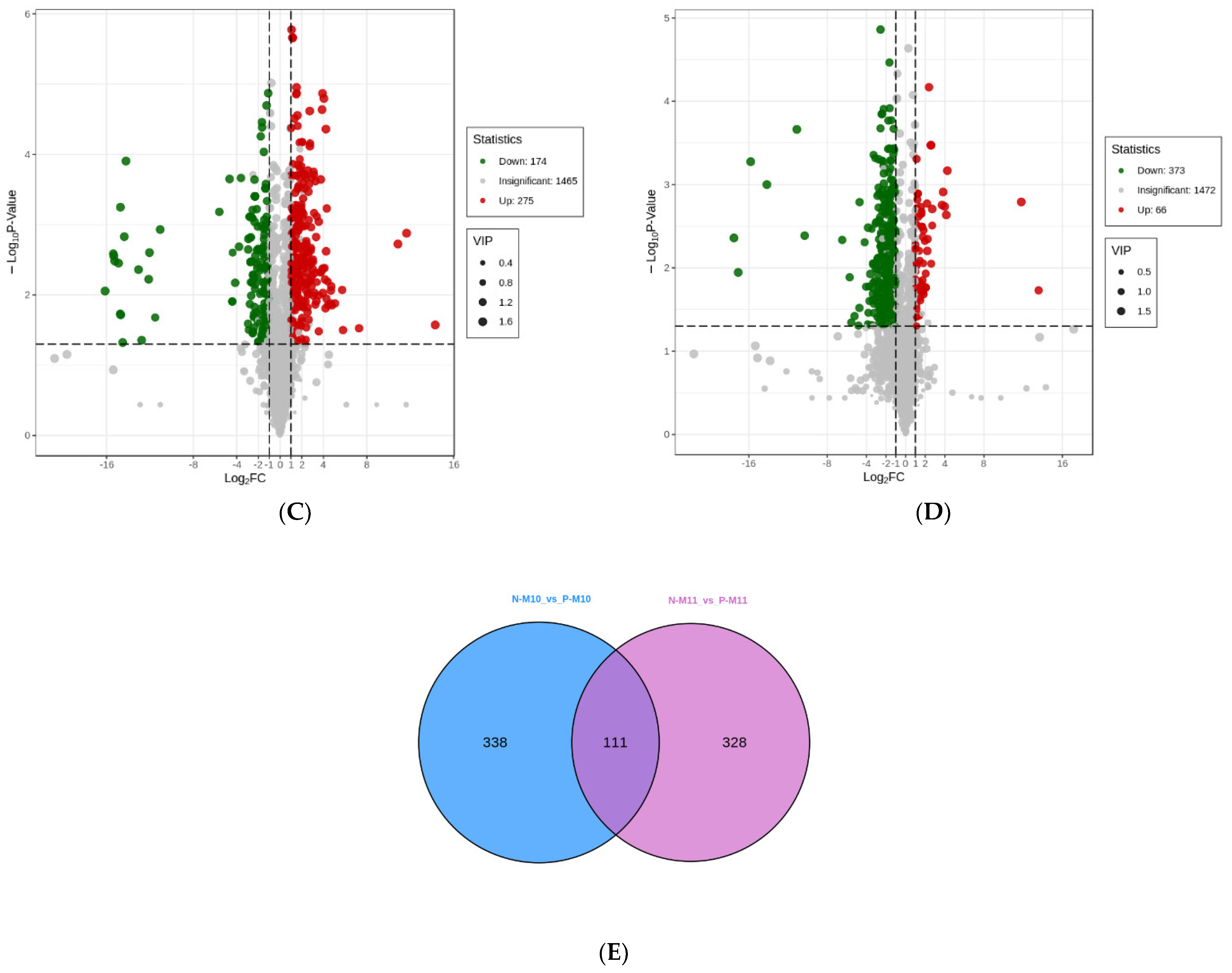
3.5. Differential Metabolites Analysis
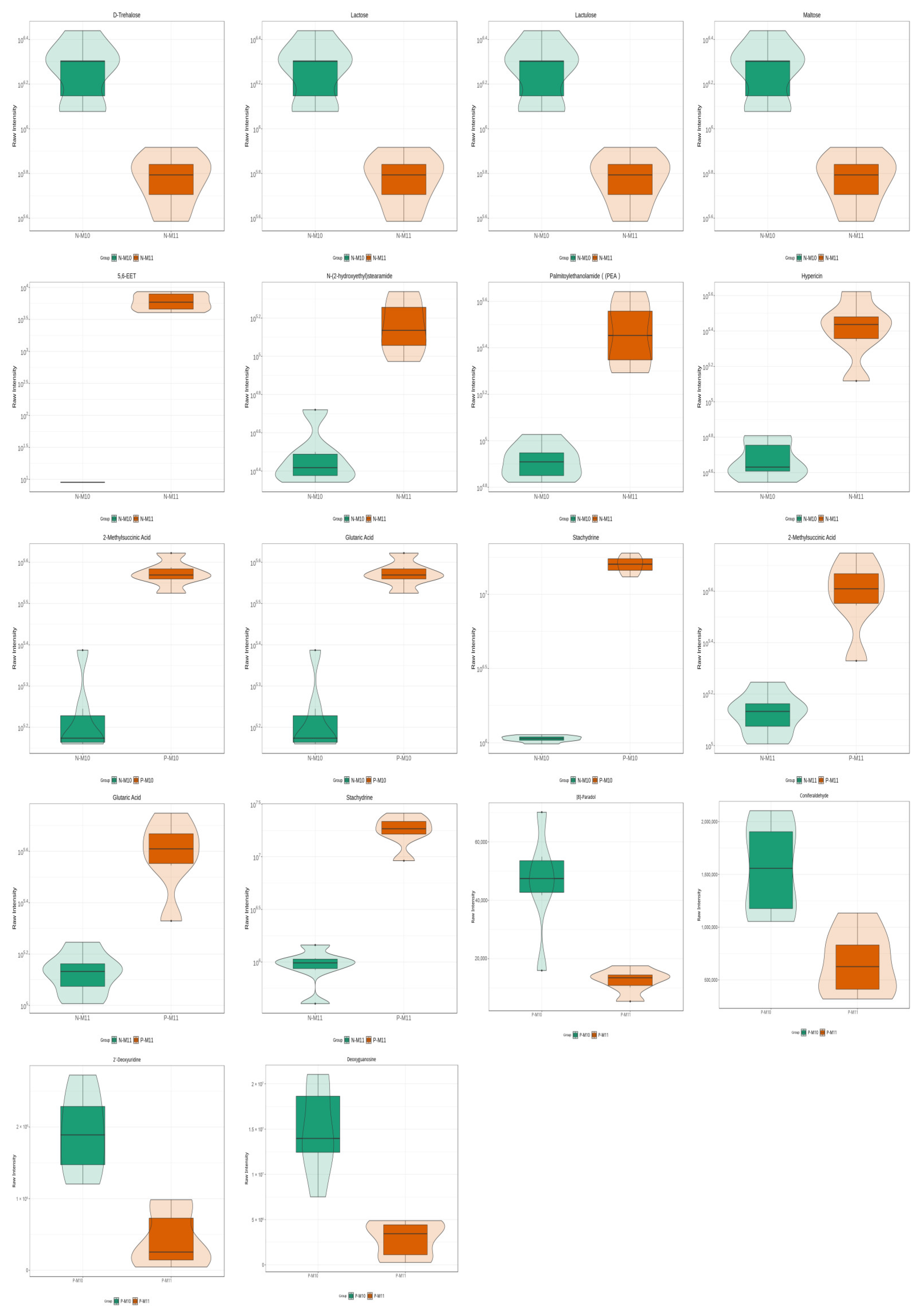
3.5.1. Effect of Pond Culture on the Metabolites of E. sinensis
3.5.2. Effect of Harvest Time (Maturity) on the Metabolites of E. sinensis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xue, J.; Jiang, T.; Chen, X.; Liu, H.; Yang, J. Multi-mineral fingerprinting analysis of the Chinese mitten crab (Eriocheir sinensis) in Yangcheng Lake during the year-round culture period. Food Chem. 2022, 390, 133167. [Google Scholar] [CrossRef] [PubMed]
- FAO. Fishery and Aquaculture Statistics. Global Aquaculture Production 1950–2019. Available online: https://www.fao.org/fishery/statistics-query/en/global_production (accessed on 23 March 2023).
- MOAC. China Fisheries Yearbook, 2021; Ministry of Agriculture of China: Beijing, China, 2021.
- Dittel, A.I.; Epifanio, C.E. Invasion biology of the Chinese mitten crab Eriochier sinensis: A brief review. J. Exp. Mar. Biol. Ecol. 2009, 374, 79–92. [Google Scholar] [CrossRef]
- GIAP. Geographical Indication Agricultural Product: Qin Lake Bamboo Weir Crab. Available online: http://120.78.129.247/Home/Product/28879 (accessed on 23 March 2023).
- QLTA. Qin Lake Tourist Attraction-Food Cultural 2-Qin Lake Bamboo Weir Crab. Available online: http://qinlake.com/en/show_04_03.html (accessed on 23 March 2023).
- Chen, D.; Zhang, M.; Shrestha, S. Compositional characteristics and nutritional quality of Chinese mitten crab (Eriocheir sinensis). Food Chem. 2007, 103, 1343–1349. [Google Scholar] [CrossRef]
- Hunter, J.E.; Zhang, J.; Kris-Etherton, P.M. Cardiovascular disease risk of dietary stearic acid compared with trans, other saturated, and unsaturated fatty acids: A systematic review. Am. J. Clin. Nutr. 2009, 91, 46–63. [Google Scholar] [CrossRef]
- Nicholls, S.J.; Lundman, P.; Harmer, J.A.; Cutri, B.; Griffiths, K.A.; Rye, K.-A.; Barter, P.J.; Celermajer, D.S. Consumption of Saturated Fat Impairs the Anti-Inflammatory Properties of High-Density Lipoproteins and Endothelial Function. J. Am. Coll. Cardiol. 2006, 48, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.; Summerbell, C.D.; Thompson, R.L.; Sills, D.; Roberts, F.; Moore, H.J.; Smith, G.D. Effects of reducing or modifying dietary fat on CVD: A systematic review and meta-analysis of randomised controlled trials. Proc. Nutr. Soc. 2011, 70, E222. [Google Scholar] [CrossRef]
- Mozaffarian, D.; Wu, J.H.Y. Omega-3 Fatty Acids and Cardiovascular Disease: Effects on Risk Factors, Molecular Pathways, and Clinical Events. J. Am. Coll. Cardiol. 2011, 58, 2047–2067. [Google Scholar] [CrossRef]
- Du, H.; Lv, H.; Xu, Z.; Zhao, S.; Huang, T.; Manyande, A.; Xiong, S. The mechanism for improving the flesh quality of grass carp (Ctenopharyngodon idella) following the micro-flowing water treatment using a UPLC-QTOF/MS based metabolomics method. Food Chem. 2020, 327, 126777. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Jiang, H.; Alhamoud, Y.; Chen, Y.; Zhuang, J.; Liu, T.; Cai, L.; Shen, W.; Wu, X.; Zheng, W.; et al. Integrated metabolomic and gene expression analyses to study the effects of glycerol monolaurate on flesh quality in large yellow croaker (Larimichthys crocea). Food Chem. 2022, 367, 130749. [Google Scholar] [CrossRef]
- Zhang, L.; Tao, N.-P.; Wu, X.; Wang, X. Metabolomics of the hepatopancreas in Chinese mitten crabs (Eriocheir sinensis). Food Res. Int. 2022, 152, 110914. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of Official Analytical Chemists International, 16th ed.; AOAC (Association of Official Analytical Chemists): Arlington, VA, USA, 1995. [Google Scholar]
- Cheng, Y.; Wu, X.; Li, J. Chinese mitten Crab Culture: Current Status and Recent Progress Towards Sustainable Development. In Aquaculture in China; John Wiley & Sons Ltd: Hoboken, NJ, USA, 2018; pp. 197–217. [Google Scholar]
- He, J.; Wu, X.; Li, J.; Huang, Q.; Huang, Z.; Cheng, Y. Comparison of the culture performance and profitability of wild-caught and captive pond-reared Chinese mitten crab (Eriocheir sinensis) juveniles reared in grow-out ponds: Implications for seed selection and genetic selection programs. Aquaculture 2014, 434, 48–56. [Google Scholar] [CrossRef]
- Kong, L.; Cai, C.; Ye, Y.; Chen, D.; Wu, P.; Li, E.; Chen, L.; Song, L. Comparison of non-volatile compounds and sensory characteristics of Chinese mitten crabs (Eriocheir sinensis) reared in lakes and ponds: Potential environmental factors. Aquaculture 2012, 364–365, 96–102. [Google Scholar] [CrossRef]
- Long, X.; Pan, J.; Wade, N.M.; Sun, Y.; Liu, Y.; Cheng, Y.; Wu, X. Effects of three feeding modes on the metabolism, antioxidant capacity, and metabolome of the adult male Chinese mitten crab Eriocheir sinensis. Aquac. Int. 2022, 30, 1101–1119. [Google Scholar] [CrossRef]
- Chen, X.; Chen, H.; Liu, Q.; Ni, K.; Wang, C. High Plasticity of the Gut Microbiome and Muscle Metabolome of Chinese Mitten Crab (Eriocheir sinensis) in Diverse Environments. J. Microbiol. Biotechnol. 2021, 31, 240–249. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Xuan, F.; Shi, H.; Xie, J.; Wang, W.; Wang, G.; Xu, W. Comparison of nutritional quality of three edible tissues of the wild-caught and pond-reared swimming crab (Portunus trituberculatus) females. LWT 2017, 75, 624–630. [Google Scholar] [CrossRef]
- Blanco, A.; Blanco, G. Chapter 16—Amino Acid Metabolism. In Medical Biochemistry; Blanco, A., Blanco, G., Eds.; Academic Press: Boston, MA, USA, 2017; pp. 367–399. [Google Scholar]
- Hu, Y.; Feng, L.; Jiang, W.; Wu, P.; Liu, Y.; Kuang, S.; Tang, L.; Zhou, X. Lysine deficiency impaired growth performance and immune response and aggravated inflammatory response of the skin, spleen and head kidney in grown-up grass carp (Ctenopharyngodon idella). Anim. Nutr. 2021, 7, 556–568. [Google Scholar] [CrossRef]
- Pekala, J.; Patkowska-Sokoła, B.; Bodkowski, R.; Jamroz, D.; Nowakowski, P.; Lochyński, S.; Librowski, T. L-carnitine-metabolic functions and meaning in humans life. Curr. Drug Metab. 2011, 12, 667–678. [Google Scholar] [CrossRef]
- Brosnan, M.E.; Brosnan, J.T. Histidine Metabolism and Function. J. Nutr. 2020, 150, 2570s–2575s. [Google Scholar] [CrossRef]
- Turner, N.; Cooney, G.J.; Kraegen, E.W.; Bruce, C.R. Fatty acid metabolism, energy expenditure and insulin resistance in muscle. J. Endocrinol. 2014, 220, T61–T79. [Google Scholar] [CrossRef]
- Rogowski, M.P.; Flowers, M.T.; Stamatikos, A.D.; Ntambi, J.M.; Paton, C.M. SCD1 activity in muscle increases triglyceride PUFA content, exercise capacity, and PPARδ expression in mice. J. Lipid Res. 2013, 54, 2636–2646. [Google Scholar] [CrossRef]
- Kumar, A.; Butt, N.A.; Levenson, A.S. Chapter 39—Natural Epigenetic-Modifying Molecules in Medical Therapy. In Medical Epigenetics; Tollefsbol, T.O., Ed.; Academic Press: Boston, MA, USA, 2016; pp. 747–798. [Google Scholar]
- Cheng, F.; Zhou, Y.; Wang, M.; Guo, C.; Cao, Z.; Zhang, R.; Peng, C. A review of pharmacological and pharmacokinetic properties of stachydrine. Pharmacol. Res. 2020, 155, 104755. [Google Scholar] [CrossRef]
- Gowthaman, N.S.K.; Lim, H.N.; Gopi, S.; Amalraj, A. 17—Identification of toxicology biomarker and evaluation of toxicity of natural products by metabolomic applications. In Inflammation and Natural Products; Gopi, S., Amalraj, A., Kunnumakkara, A., Thomas, S., Eds.; Academic Press: Boston, MA, USA, 2021; pp. 407–436. [Google Scholar]
- Gerstner, B.; Gratopp, A.; Marcinkowski, M.; Sifringer, M.; Obladen, M.; Bührer, C. Glutaric Acid and Its Metabolites Cause Apoptosis in Immature Oligodendrocytes: A Novel Mechanism of White Matter Degeneration in Glutaryl-CoA Dehydrogenase Deficiency. Pediatr. Res. 2005, 57, 771–776. [Google Scholar] [CrossRef]
- Romashko, M.; Schragenheim, J.; Abraham, N.G.; McClung, J.A. Epoxyeicosatrienoic Acid as Therapy for Diabetic and Ischemic Cardiomyopathy. Trends Pharmacol. Sci. 2016, 37, 945–962. [Google Scholar] [CrossRef] [PubMed]
- Sisignano, M.; Park, C.-K.; Angioni, C.; Zhang, D.D.; von Hehn, C.; Cobos, E.J.; Ghasemlou, N.; Xu, Z.-Z.; Kumaran, V.; Lu, R.; et al. 5,6-EET Is Released upon Neuronal Activity and Induces Mechanical Pain Hypersensitivity via TRPA1 on Central Afferent Terminals. J. Neurosci. 2012, 32, 6364. [Google Scholar] [CrossRef] [PubMed]
- Lian, J.S.; Liu, W.; Hao, S.R.; Guo, Y.Z.; Huang, H.J.; Chen, D.Y.; Xie, Q.; Pan, X.P.; Xu, W.; Yuan, W.X.; et al. A serum metabonomic study on the difference between alcohol- and HBV-induced liver cirrhosis by ultraperformance liquid chromatography coupled to mass spectrometry plus quadrupole time-of-flight mass spectrometry. Chin. Med. J. 2011, 124, 1367–1373. [Google Scholar] [CrossRef] [PubMed]
- Keppel Hesselink, J.M. Chapter 4—Palmitoylethanolamid and Other Lipid Autacoids Against Neuroinflammation, Pain, and Spasms in Multiple Sclerosis. In Nutrition and Lifestyle in Neurological Autoimmune Diseases; Watson, R.R., Killgore, W.D.S., Eds.; Academic Press: Boston, MA, USA, 2017; pp. 29–37. [Google Scholar]
- McIlwain, M.E.; Minassian, A.; Perry, W. Chapter 9—The Cannabinoid System in Nicotine Dependence and Withdrawal. In Negative Affective States and Cognitive Impairments in Nicotine Dependence; Hall, F.S., Young, J.W., Der-Avakian, A., Eds.; Academic Press: San Diego, CA, USA, 2017; pp. 153–176. [Google Scholar]
- Kubin, A.; Wierrani, F.; Burner, U.; Alth, G.; Grünberger, W. Hypericin—The facts about a controversial agent. Curr. Pharm. Des. 2005, 11, 233–253. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Meat | Cream | |||||||
|---|---|---|---|---|---|---|---|---|
| N-M10 | P-M10 | N-M11 | P-M11 | N-M10 | P-M10 | N-M11 | P-M11 | |
| Asp 1 | N.D. | 0.02 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 1.08 ± 0.13 C | 1.41 ± 0.07 B | 1.72 ± 0.18 A | 1.50 ± 0.11 AB |
| Thr | 0.63 ± 0.07 a | 0.59 ± 0.06 ab | N.D. | 0.47 ± 0.05 b | 2.49 ± 0.22 A | 2.13 ± 0.16 A | 2.58 ± 0.32 A | 2.49 ± 0.33 A |
| Ser | 0.18 ± 0.02 b | 0.32 ± 0.07 a | 0.15 ± 0.04 b | 0.24 ± 0.03 ab | 1.34 ± 0.12 A | 1.41 ± 0.32 A | 1.25 ± 0.23 A | 0.98 ± 0.15 B |
| Glu | 0.17 ± 0.01 b | 0.32 ± 0.09 a | 0.12 ± 0.02 b | 0.31 ± 0.07 a | 2.95 ± 0.15 B | 2.91 ± 0.34 B | 3.80 ± 0.29 A | 3.88 ± 0.35 A |
| Gly | 1.69 ± 0.31 b | 3.59 ± 0.24 a | 1.50 ± 0.24 b | 3.80 ± 0.23 a | 1.32 ± 0.16 B | 1.09 ± 0.14 B | 1.67 ± 0.18 A | 1.89 ± 0.38 A |
| Ala | 2.20 ± 0.07 b | 4.87 ± 0.74 a | 1.78 ± 0.34 b | 5.07 ± 0.70 a | 2.81 ± 0.36 C | 2.72 ± 0.22 C | 4.86 ± 0.12 B | 5.50 ± 0.22 A |
| Cys | 0.04 ± 0.01 b | 0.08 ± 0.02 a | 0.02 ± 0.00 c | 0.07 ± 0.02 a | 0.26 ± 0.03 B | 0.32 ± 0.03 A | 0.25 ± 0.01 B | 0.28 ± 0.07 AB |
| Val | 0.14 ± 0.03 b | 0.27 ± 0.10 a | 0.11 ± 0.04 b | 0.29 ± 0.07 a | 1.68 ± 0.18 A | 1.92 ± 0.18 A | 1.99 ± 0.47 A | 2.04 ± 0.23 A |
| Met | 0.14 ± 0.01 bc | 0.31 ± 0.11 a | 0.10 ± 0.03 c | 0.23 ± 0.02 ab | 0.83 ± 0.03 A | 0.85 ± 0.12 A | 0.81 ± 0.06 A | 0.86 ± 0.04 A |
| Ile | 0.07 ± 0.02 bc | 0.15 ± 0.05 a | 0.05 ± 0.01 c | 0.13 ± 0.03 ab | 1.23 ± 0.19 A | 1.25 ± 0.35 A | 1.49 ± 0.09 A | 1.54 ± 0.22 A |
| Leu | 0.16 ± 0.01 ab | 0.29 ± 0.12 a | 0.12 ± 0.02 b | 0.22 ± 0.09 ab | 2.36 ± 0.29 A | 2.55 ± 0.44 A | 2.70 ± 0.28 A | 2.75 ± 0.12 A |
| Tyr | 0.07 ± 0.02 b | 0.27 ± 0.09 a | 0.09 ± 0.02 b | 0.14 ± 0.03 b | 1.36 ± 0.29 A | 1.32 ± 0.14 A | 1.22 ± 0.25 A | 1.16 ± 0.20 A |
| Phe | 0.10 ± 0.04 b | 0.29 ± 0.08 a | 0.11 ± 0.03 b | 0.23 ± 0.07 a | 1.42 ± 0.05 A | 1.57 ± 0.15 A | 1.42 ± 0.15 A | 1.45 ± 0.22 A |
| Lys | 0.11 ± 0.02 b | 0.46 ± 0.19 a | 0.13 ± 0.01 b | 0.51 ± 0.03 a | 2.12 ± 0.27 A | 2.34 ± 0.38 A | 2.74 ± 0.24 A | 2.09 ± 0.48 A |
| His | 0.14 ± 0.03 b | 0.31 ± 0.08 a | 0.12 ± 0.03 b | 0.25 ± 0.04 a | 0.76 ± 0.06 AB | 0.69 ± 0.17 B | 0.89 ± 0.09 A | 0.90 ± 0.07 A |
| Arg | 2.53 ± 0.19 b | 3.81 ± 0.18 a | 1.86 ± 0.33 c | 4.26 ± 0.17 a | 2.67 ± 0.27 A | 2.70 ± 0.29 A | 2.99 ± 0.14 A | 3.28 ± 0.52 A |
| Pro | 1.06 ± 0.06 b | 1.19 ± 0.22 b | 1.07 ± 0.05 b | 2.32 ± 0.17 a | 1.44 ± 0.11 B | 1.50 ± 0.24 B | 2.91 ± 0.40 A | 2.43 ± 0.32 A |
| Essential AA | 1.49 ± 0.09 c | 2.67 ± 0.37 a | 0.74 ± 0.13 d | 2.33 ± 0.13 b | 12.89 ± 0.45 C | 13.30 ± 0.37 BC | 14.62 ± 0.1 A | 14.12 ± 0.57 AB |
| Total umami AA | 0.17 ± 0.01 b | 0.34 ± 0.09 a | 0.13 ± 0.02 b | 0.32 ± 0.07 a | 4.03 ± 0.25 B | 4.32 ± 0.39 B | 5.52 ± 0.29 A | 5.38 ± 0.24 A |
| Total sweet AA | 5.76 ± 0.39 c | 10.56 ± 0.80 b | 4.50 ± 0.22 d | 11.90 ± 0.49 a | 9.40 ± 0.30 B | 8.85 ± 0.22 B | 13.27 ± 0.96 A | 13.29 ± 1.10 A |
| Total bitter AA | 3.39 ± 0.29 b | 5.89 ± 0.53 a | 2.60 ± 0.25 c | 6.12 ± 0.28 a | 13.07 ± 0.54 B | 13.87 ± 0.44 B | 15.03 ± 0.41 A | 14.91 ± 0.32 A |
| Total AA | 9.43 ± 0.31 c | 17.14 ± 0.55 b | 7.34 ± 0.04 d | 18.55 ± 0.64 a | 28.12 ± 0.15 B | 28.68 ± 0.99 B | 35.29 ± 0.67 A | 35.02 ± 1.20 A |
| Meat | Cream | |||||||
|---|---|---|---|---|---|---|---|---|
| N-M10 | P-M10 | N-M11 | P-M11 | N-M10 | P-M10 | N-M11 | P-M11 | |
| C14:0 | 0.35 ± 0.06 | 0.49 ± 0.03 | 0.38 ± 0.02 | 0.62 ± 0.03 | 0.94 ± 0.02 | 1.28 ± 0.05 | 0.77 ± 0.06 | 1.23 ± 0.03 |
| C15:0 | N.D. | 0.32 ± 0.02 | 0.21 ± 0.01 | 0.28 ± 0.02 | 0.45 ± 0.03 | 0.73 ± 0.04 | 0.43 ± 0.02 | 0.56 ± 0.01 |
| C16:0 | 14.50 ± 0.46 | 15.13 ± 0.11 | 14.15 ± 0.15 | 14.04 ± 0.40 | 20.17 ± 0.36 | 21.83 ± 0.42 | 19.26 ± 0.18 | 19.49 ± 0.12 |
| C17:0 | 0.57 ± 0.08 | 0.66 ± 0.05 | 0.45 ± 0.02 | 0.54 ± 0.02 | 0.35 ± 0.09 | 0.48 ± 0.05 | 0.24 ± 0.01 | 0.35 ± 0.05 |
| C18:0 | 7.56 ± 0.35 | 7.29 ± 0.40 | 6.71 ± 0.25 | 6.40 ± 0.01 | 2.49 ± 0.30 | 2.92 ± 0.24 | 2.49 ± 0.13 | 2.60 ± 0.06 |
| C20:0 | N.D. | N.D. | N.D. | N.D. | 0.23 ± 0.02 | N.D. | N.D. | 0.21 ± 0.02 |
| C24:0 | 12.47 ± 0.47 | 9.98 ± 0.14 | 12.48 ± 0.16 | 11.13 ± 0.13 | 1.71 ± 0.20 | 1.64 ± 0.10 | 1.42 ± 0.12 | 1.54 ± 0.16 |
| SFA 1 | 35.45 ± 1.00 a | 33.87 ± 0.43 bc | 34.38 ± 0.55 b | 33.01 ± 0.21 c | 26.34 ± 0.26 B | 28.88 ± 0.72 A | 24.61 ± 0.42 C | 25.98 ± 0.44 B |
| C16:1 n7 | 3.14 ± 0.17 | 3.20 ± 0.12 | 2.89 ± 0.02 | 2.79 ± 0.06 | 8.72 ± 0.15 | 7.81 ± 0.09 | 7.59 ± 0.10 | 7.71 ± 0.02 |
| C17:1 n7 | 0.47 ± 0.06 | 0.51 ± 0.03 | 0.45 ± 0.06 | 0.44 ± 0.02 | 0.54 ± 0.05 | 0.50 ± 0.04 | 0.48 ± 0.02 | 0.52 ± 0.02 |
| C18:1 n9 | 20.71 ± 0.51 | 22.74 ± 0.13 | 21.45 ± 0.11 | 21.75 ± 0.09 | 28.05 ± 0.17 | 27.45 ± 0.60 | 29.23 ± 0.16 | 29.06 ± 0.11 |
| C20:1 n9 | 0.42 ± 0.08 | 0.51 ± 0.02 | 0.44 ± 0.01 | 1.58 ± 0.03 | 0.53 ± 0.05 | 0.74 ± 0.09 | 0.61 ± 0.02 | 0.92 ± 0.02 |
| MUFA | 24.74 ± 0.67 b | 26.96 ± 0.05 a | 25.24 ± 0.15 b | 26.56 ± 0.21 a | 37.84 ± 0.29 A | 36.50 ± 0.53 B | 37.92 ± 0.28 A | 38.22 ± 0.13 A |
| C18:2 n6 | 17.27 ± 0.24 | 13.27 ± 0.22 | 18.11 ± 0.19 | 15.08 ± 0.09 | 25.43 ± 0.27 | 16.54 ± 0.43 | 26.47 ± 0.36 | 19.60 ± 0.19 |
| C18:3 n3 | 1.66 ± 0.06 | 1.39 ± 0.06 | 1.34 ± 0.04 | 0.21 ± 0.02 | 2.39 ± 0.08 | 2.35 ± 0.19 | 1.70 ± 0.03 | 2.07 ± 0.01 |
| C20:2 n6 | 1.31 ± 0.08 | 1.81 ± 0.11 | 1.37 ± 0.07 | 1.62 ± 0.10 | 0.90 ± 0.10 | 2.07 ± 0.29 | 1.09 ± 0.05 | 1.68 ± 0.10 |
| C20:3 n3 | 0.35 ± 0.03 | 0.29 ± 0.33 | 0.34 ± 0.01 | N.D. | 0.31 ± 0.05 | 0.35 ± 0.48 | 0.32 ± 0.01 | 0.28 ± 0.03 |
| C20:4 n6 | 4.62 ± 0.17 | 7.08 ± 0.23 | 4.56 ± 0.19 | 6.32 ± 0.17 | 1.08 ± 0.12 | 2.79 ± 0.20 | 1.11 ± 0.14 | 2.10 ± 0.09 |
| C22:6 n3 | 6.30 ± 0.40 | 5.48 ± 0.26 | 6.45 ± 0.27 | 7.13 ± 0.20 | 0.93 ± 0.22 | 1.08 ± 0.06 | 0.89 ± 0.11 | 2.01 ± 0.19 |
| PUFA | 31.50 ± 0.64 a | 29.31 ± 0.07 c | 32.17 ± 0.37 a | 30.36 ± 0.35 b | 31.04 ± 0.22 A | 25.18 ± 0.61 C | 31.58 ± 0.63 A | 27.74 ± 0.26 B |
| n3 PUFA | 8.30 ± 0.46 a | 7.15 ± 0.27 b | 8.13 ± 0.27 a | 7.34 ± 0.19 b | 3.64 ± 0.22 B | 3.78 ± 0.14 B | 2.91 ± 0.11 C | 4.36 ± 0.16 A |
| n6 PUFA | 23.20 ± 0.23 b | 22.16 ± 0.21 c | 24.04 ± 0.11 a | 23.02 ± 0.17 b | 27.40 ± 0.28 B | 21.40 ± 0.57 D | 28.67 ± 0.54 A | 23.39 ± 0.33 C |
| n3/n6 | 0.36 ± 0.02 a | 0.32 ± 0.02 b | 0.34 ± 0.01 ab | 0.32 ± 0.01 b | 0.13 ± 0.01 B | 0.18 ± 0.01 A | 0.10 ± 0.00 C | 0.18 ± 0.01 A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, Y.; Wang, Y.; Liu, P.; Chen, J.; Zhang, C. Uncovering the Nutritive Profiles of Adult Male Chinese Mitten Crab (E. sinensis) Harvested from the Pond and Natural Water Area of Qin Lake Based on Metabolomics. Foods 2023, 12, 2178. https://doi.org/10.3390/foods12112178
Ye Y, Wang Y, Liu P, Chen J, Zhang C. Uncovering the Nutritive Profiles of Adult Male Chinese Mitten Crab (E. sinensis) Harvested from the Pond and Natural Water Area of Qin Lake Based on Metabolomics. Foods. 2023; 12(11):2178. https://doi.org/10.3390/foods12112178
Chicago/Turabian StyleYe, Yuhui, Yulong Wang, Pengyan Liu, Jian Chen, and Cunzheng Zhang. 2023. "Uncovering the Nutritive Profiles of Adult Male Chinese Mitten Crab (E. sinensis) Harvested from the Pond and Natural Water Area of Qin Lake Based on Metabolomics" Foods 12, no. 11: 2178. https://doi.org/10.3390/foods12112178