
Time Domain NMR Approach in the Chemical and Physical Characterization of Hazelnuts (Corylus avellana L.)

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Origin of the Samples
2.2. Sampling
2.3. TD NMR Spectroscopy
2.3.1. Analysis of the Liquid Content (X)
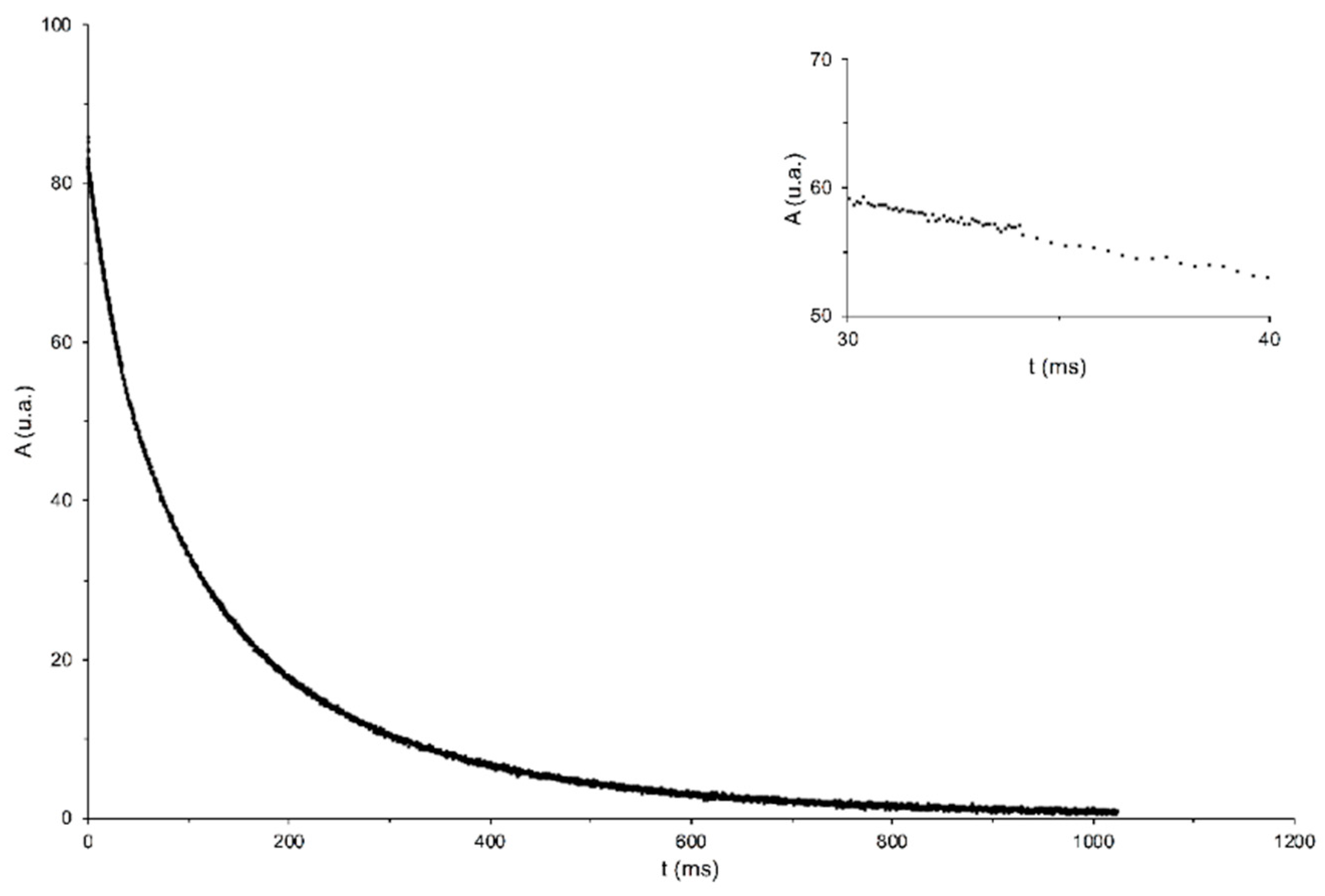
2.3.2. Spin Eco Sequence: Simultaneous Determination of Moisture and Oil Content
2.3.3. CPMG Sequence: Determination of Spin–Spin Relaxation Times
2.3.4. Cryo-SEM of Hazelnut Section
2.3.5. Statistical Analysis
3. Results and Discussion
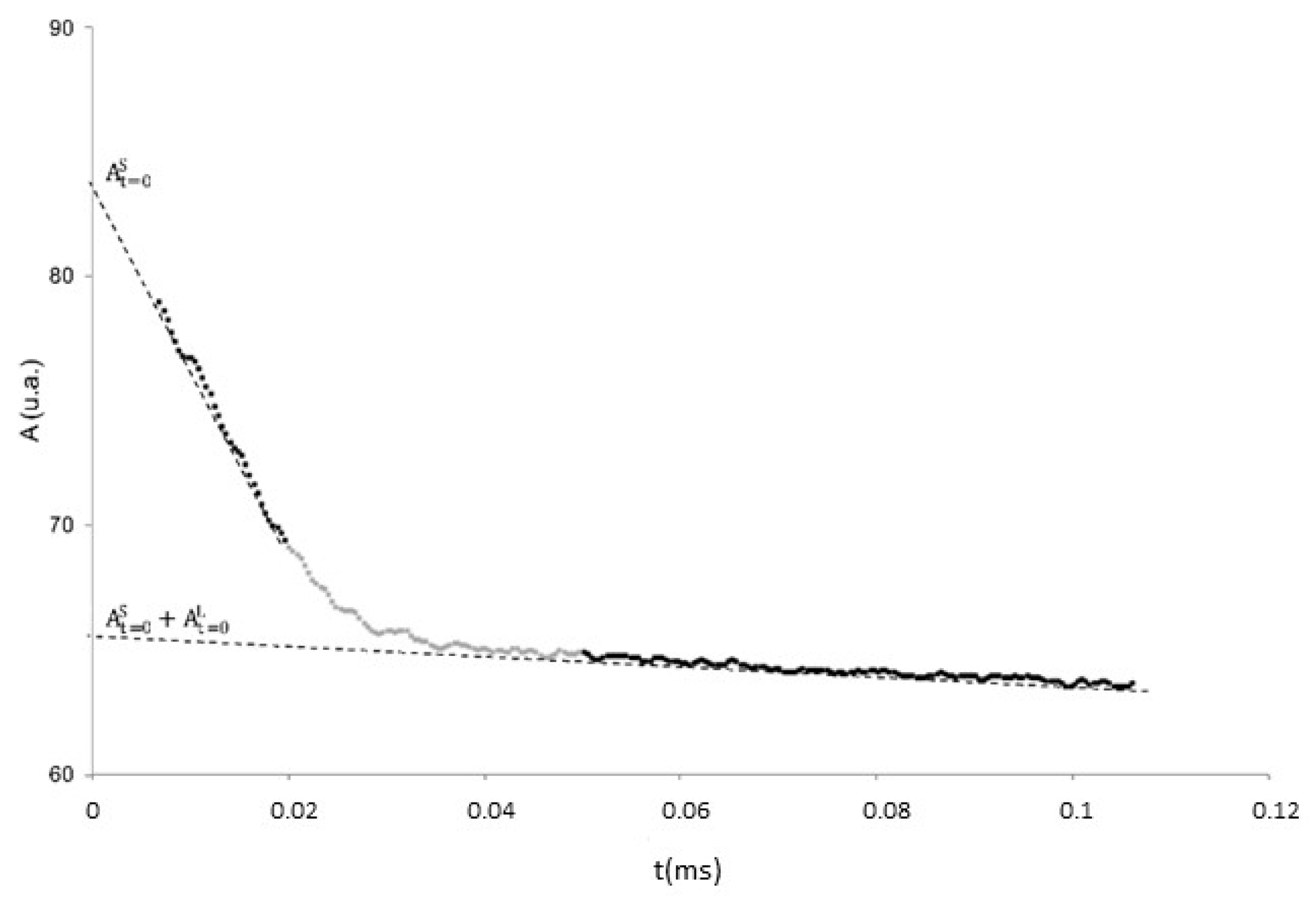
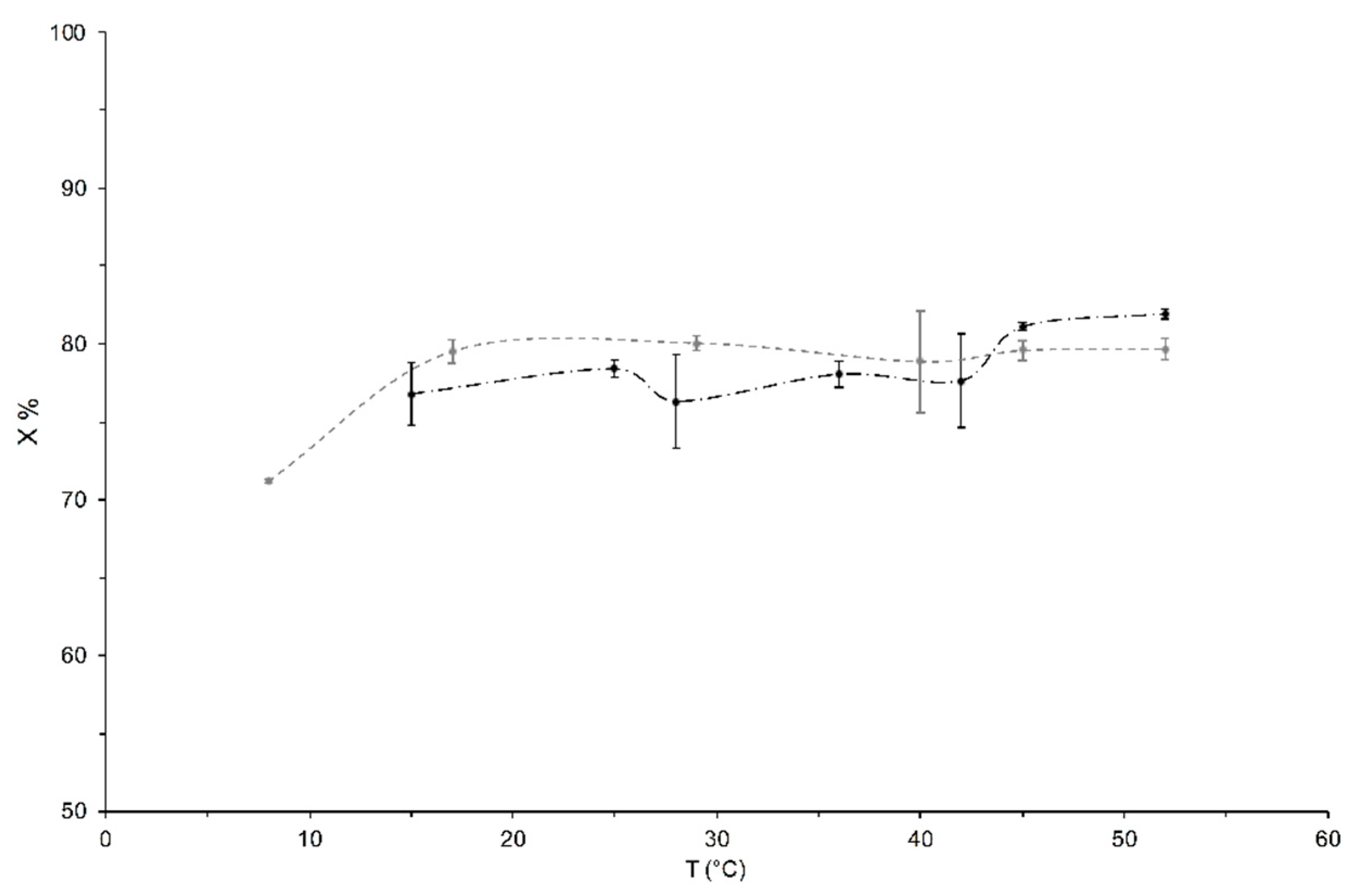
3.1. Measurements of the Liquid Phase Content, X, as a Function of Temperature
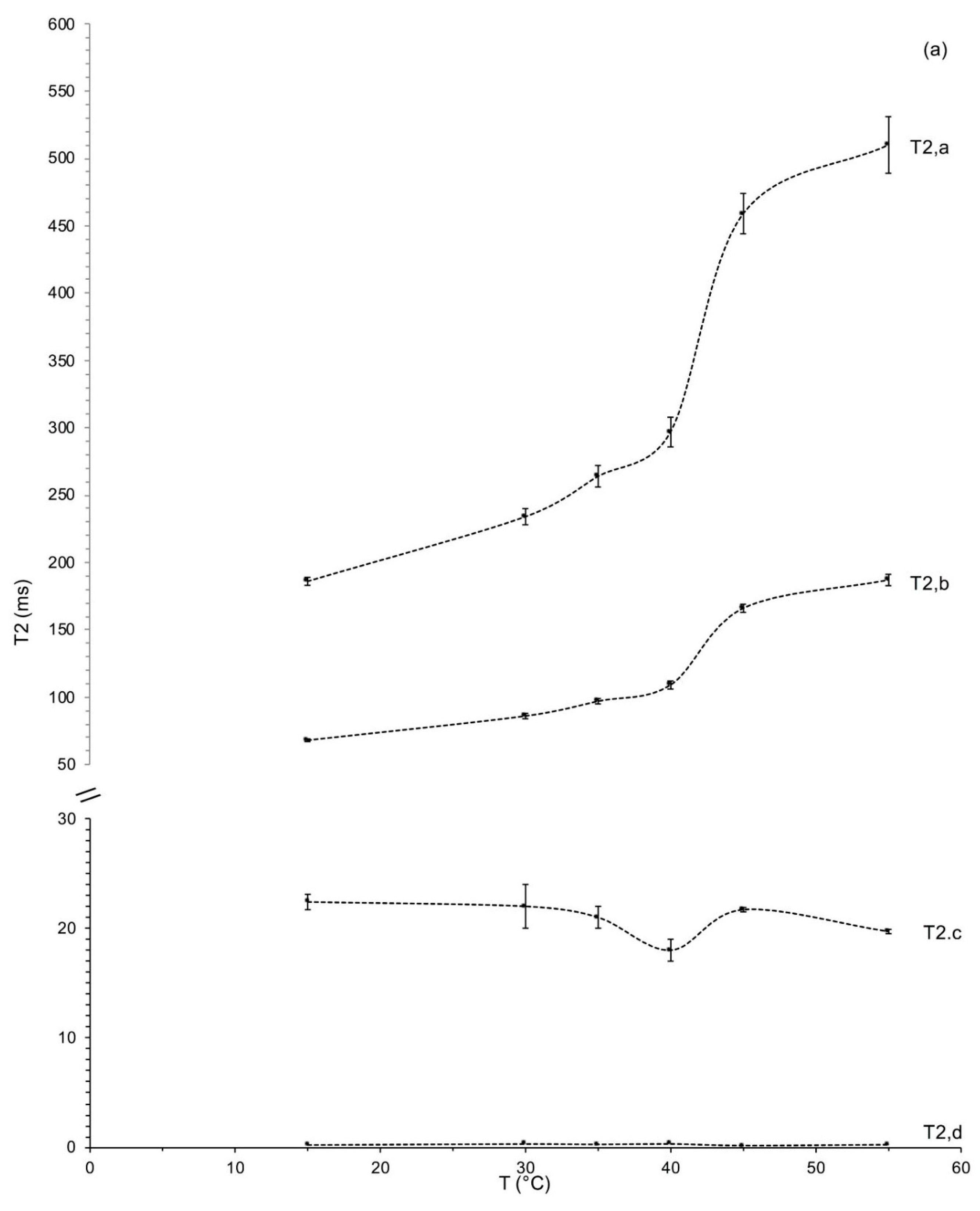
3.2. Measurements of the Spin–Spin Relaxation Times
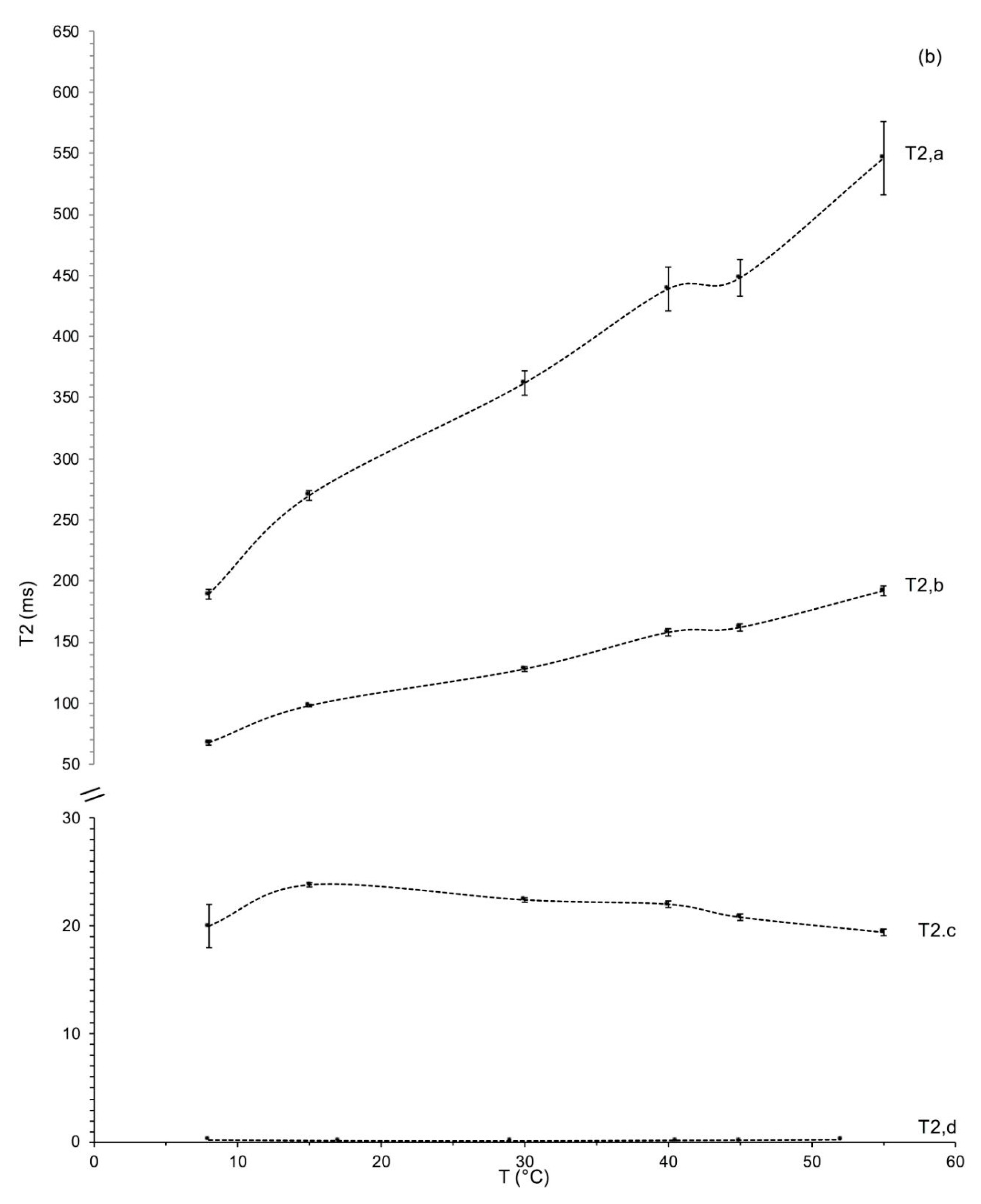
Measurements of the Relaxation Time Spin–Spin as a Function of Temperature
4. Conclusions and Future Trends
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. FAO Statistical Yearbook—World Food and Agriculture; FAO: Rome, Italy, 2022; ISBN 978-92-5-136930-2. [Google Scholar]
- Zinnanti, C.; Schimmenti, E.; Borsellino, V.P.; Severini, S. Economic performance and risk of farming systems specialized in perennial crops: An analysis of Italian hazelnut production. Agric. Syst. 2019, 176, 102645. [Google Scholar] [CrossRef]
- Manfredi, M.; Robotti, E.; Quasso, F.; Mazzucco, E.; Calabrese, G.; Marengo, M. Fast classification of hazelnut cultivars through portable infrared spectroscopy and chemometrics. Spectrochim. Acta A 2018, 189, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Król, K.; Gantner, M. Morphological Traits and Chemical Composition of Hazelnut from Different Geographical Origins: A Review. Agriculture 2020, 10, 375. [Google Scholar] [CrossRef]
- Müller, A.K.; Helms, U.; Rohrer, C.; Möhler, M.; Hellwig, F.; Glei, M.; Schwerdtle, T.; Lorkowski, S.; Dawczynski, C. Nutrient Composition of Different Hazelnut Cultivars Grown in Germany. Foods 2020, 9, 1596. [Google Scholar] [CrossRef]
- Dreher, M.L.; Maher, C.V.; Kearney, P. The traditional and emerging role of nuts in healthful diets. Nutr. Rev. 1996, 54, 241–245. [Google Scholar] [CrossRef]
- Bo, S.; Ponzo, V.; Goitre, I.; Fadda, M.; Pezzana, A.; Beccuti, G.; Gambino, R.; Cassader, M.; Soldati, L.; Broglio, F. Predictive role of the Mediterranean diet on mortality in individuals at low cardiovascular risk: A 12-year follow-up population-based cohort study. J. Transl. Med. 2016, 14, 91. [Google Scholar] [CrossRef]
- Becerra-Tomás, N.; Paz-Graniel, I.; Kendall, C.W.C.; Kahleova, H.; Rahelić, D.; Sievenpiper, J.L.; Salas-Salvadó, J. Nut consumption and incidence of cardiovascular diseases and cardiovascular disease mortality: A meta-analysis of prospective cohort studies. Nutr. Rev. 2019, 77, 691–709. [Google Scholar] [CrossRef]
- Ros, E.; Singh, A.; O’Keefe, J.H. Nuts: Natural Pleiotropic Nutraceuticals. Nutrients 2021, 13, 3269. [Google Scholar] [CrossRef]
- Zhang, D.; Dai, C.; Zhou, L.; Li, Y.; Liu, K.; Deng, Y.J.; Li, N.; Zheng, Y.; Hao, Q.; Yang, S.; et al. Meta-analysis of the association between nut consumption and the risks of cancer incidence and cancer-specific mortality. Aging 2020, 12, 10772–10794. [Google Scholar] [CrossRef]
- Available online: https://www.fda.gov/food/cfsan-constituent-updates/fda-completes-review-qualified-health-claim-petition-macadamia-nuts-and-risk-coronary-heart-disease (accessed on 8 February 2023).
- Ozdemir, M.; Açkurt, F.; Kaplan, M.; Yildiz, M.; Loker, M.; Gurcan, T.; Biringen, G.; Okay, A.; Seyhan, G.F. Evaluation of new Turkish hybrid hazelnut (Corylus avellana L.) varieties: Fatty acid composition, α-tocopherol content, mineral composition and stability. Food Chem. 2001, 73, 411–415. [Google Scholar] [CrossRef]
- Parcerisa, J.; Rafecas, M.; Codony, R.; Boatella, J. Triacylglycerol and Phospholipid Composition of Hazelnut (Corylus avellana L.) lipid fraction during Development. J. Agric. Food Chem. 1999, 47, 1410–1415. [Google Scholar] [CrossRef] [PubMed]
- Bignami, C.; Bertazza, G.; Cristofori, V.; Troso, D. Kernel quality and composition of hazelnut (Corylus avellana L.) cultivars. In Proceedings of the Sixth International Congress on Hazelnut, Tarragona-Reus, Spain, 14–18 June 2005; Volume 686, pp. 477–484. [Google Scholar]
- Bignami, C.; Bertazza, G.; Cristofori, V.; Scossa, A. Dinamica della composizione del seme di tre cultivar di nocciolo (Corylus avellana L.) durante lo sviluppo del frutto. In Proceedings of the II Convegno Nazionale Sul Nocciolo, Giffoni Valle Piana (SA), Italy, 5–6 October 2005; pp. 270–278. [Google Scholar]
- Granata, M.U.; Bracco, F.; Gratani, L.; Catoni, R.; Corana, F.; Mannucci, B.; Sartori, F.; Martino, E. Fatty acid content profile and main constituents of Corylus avellana kernel in wild type and cultivars growing in Italy. Nat. Prod. Res. 2017, 31, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, I.; Sousa, A.; Morais, J.S.; Ferreira, I.C.F.R.; Bento, A.; Estevinho, L.; Pereira, J.A. Chemical composition, and antioxidant and antimicrobial activities of three hazelnut (Corylus avellana L.) Cultivars. Food Chem. Toxicol. 2008, 46, 1801–1807. [Google Scholar] [CrossRef] [PubMed]
- Sciubba, F.; Di Cocco, M.E.; Gianferri, R.; Impellizzeri, D.; Mannina, L.; De Salvador, F.R.; Venditti, A.; Delfini, M. Metabolic profile of different Italian cultivars of hazelnut (Corylus avellana) by nuclear magnetic resonance spectroscopy. Nat. Prod. Res. 2014, 28, 1075–1081. [Google Scholar] [CrossRef]
- Guiné, R.P.F.; Almeida, C.F.F.; Correia, P.M.R.; Mendes, M. Modelling the Influence of Origin, Packing and Storage on Water Activity, Colour and Texture of Almonds, Hazelnuts and Walnuts Using Artificial Neural Networks. Food Bioprocess Technol. 2015, 8, 1113–1125. [Google Scholar] [CrossRef]
- Perna, S.; Giacosa, A.; Bonitta, G.; Bologna, C.; Isu, A.; Guido, D.; Rondanelli, M. Effects of Hazelnut Consumption on Blood Lipids and Body Weight: A Systematic Review and Bayesian Meta-Analysis. Nutrients 2016, 8, 747. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.; Ware, L.; Tey, S.L. Effects of hazelnut consumption on cardiometabolic risk factors and acceptance: A systematic review. Int. J. Environ. Res. Public Health 2022, 19, 2880. [Google Scholar] [CrossRef]
- Garcia-Lorda, P.; Megias-Rangil, I.; Salas-Salvado, J. Nut consumption, body weight and insulin resistance. Eur. J. Cin. Nutr. 2003, 57 (Suppl. S1), S8–S11. [Google Scholar] [CrossRef]
- Li, C.H.; Shelp, G.; Wright, A.J. Influence of nut structure and processing on lipid bioaccessibility and absorption. Curr. Opin. Food Sci. 2022, 49, 100966. [Google Scholar] [CrossRef]
- Lamberti, C.; Nebbia, S.; Balestrini, R.; Marengo, E.; Manfredi, M.; Pavese, V.; Cirrincione, S.; Giuffrida, M.G.; Cavallarin, L.; Acquadro, A.; et al. Identification of a caleosin associated with hazelnut (Corylus avellana L.) oil bodies. Plant Biol. 2020, 22, 404–409. [Google Scholar] [CrossRef]
- Brosio, E.; Belotti, M.; Gianferri, R. Diffusive and Chemical Exchange Model: Practical Hits to Investigate Water Dynamics in Foods, 1st ed.; Greco, L.V., Bruno, M.N., Eds.; Food Science and Technology; Nova Science Publishers: Hauppauge, NY, USA, 2008; pp. 323–340. [Google Scholar]
- Mannina, L.; Sobolev, A.P.; Aru, V.; Bellomaria, A.; Bertocchi, F.; Botta, B.; Cagliani, L.R.; Caligiani, A.; Capozzi, F.; Çela, D.; et al. NMR methodologies in food analysis. In Analytical Chemistry: Developments, Applications and Challenges in Food Analysis; Nova Science Publishers: Hauppauge, NY, USA, 2017; pp. 103–156. [Google Scholar]
- Tang, F.; Vasas, M.; Hatzakis, E.; Spyros, A. Chapter Five—Magnetic Resonance Applications in Food Analysis; Graham, A., Ed.; Webb, Annual Reports on NMR Spectroscopy; Academic Press: Cambridge, MA, USA, 2019; Volume 98, pp. 239–306. [Google Scholar] [CrossRef]
- Gianferri, R.; Maioli, M.; Delfini, M.; Brosio, E. A low-resolution and high resolution nuclear magnetic resonance integrated approach to investigate the physical structure and metabolic profile of Mozzarella di Bufala Campana cheese. Int. Dairy J. 2007, 17, 167–176. [Google Scholar] [CrossRef]
- Lucarini, M.; Durazzo, A.; Sciubba, F.; Di Cocco, M.E.; Gianferri, R.; Alise, M.; Santini, A.; Delfini, M.; Lombardi-Boccia, G. Stability of the Meat Protein Type I Collagen: Influence of pH, Ionic Strength, and Phenolic Antioxidant. Foods 2020, 9, 480. [Google Scholar] [CrossRef] [PubMed]
- Hills, B.P.; Takacs, S.F.; Belton, P.S. A new interpretation of proton NMR relaxation time measurements of water in food. Food Chem. 1990, 37, 95–111. [Google Scholar] [CrossRef]
- de Carvalho, L.C.; Manhas Verbi Pereira, F.; de Lelis Medeiros de Morais, C.; Gomes de Lima, K.M.; de Almeida Teixeira, G.H. Assessment of macadamia kernel quality defects by means of near infrared spectroscopy and nuclear magnetic resonance. Food Control. 2019, 106, 106695. [Google Scholar] [CrossRef]
- Ribeiro, Ú.A.; Queiroz, L.C.; Marassi, A.G.; Carvalho, A.S.; Barros, G.A.; Consalter, D.M.; Bezerra, J.A.; Santos, A.D.C.; Colnago, L.A.; Machado, M.B. Development of a TD-NMR Method to Monitor Brazil Nuts Oil Content: A Green and Low-Cost Based Approach. J. Braz. Chem. Soc. 2021, 32, 1405–1412. [Google Scholar] [CrossRef]
- Di Caro, D.; Liguori, C.; Pietrosanto, A.; Sommella, P. Quality Assessment of the Inshell Hazelnuts Based on TD-NMR Analysis, Trans. Instrum. Meas. 2020, 69, 3770–3779. [Google Scholar] [CrossRef]
- Di Caro, D.; Liguori, C.; Pietrosanto, A.; Sommella, P. Hazelnut oil classification by NMR techniques. Trans. Instrum. Meas. 2017, 66, 928–934. [Google Scholar] [CrossRef]
- N. 186 del 12-8-2009; Disciplinare di Produzione della Denominazione di Origine Protetta “nocciola romana”. Serie Generale. Gazzetta Ufficiale della Repubblica Italiana: Turin, Italy, 2009.
- Cristofori, V.; Pica, A.L.; Silvestri, C.; Bizzarri, S. Phenology and yield evaluation of hazelnut cultivars in Latium region. Acta Hortic. 2018, 1226, 123–130. [Google Scholar] [CrossRef]
- Commission Regulation (EC) No. 1284/2002; 15 July 2002 Laying Down the Marketing Standard for Hazelnuts in Shell. Publications Office of the EU: Luxembourg, 2002.
- Pedersen, H.T.; Bro, R.; Engelsen, S.B. Towards rapid and unique curve resolution of low-field NMR relaxation data: Trilinear SLICING versus Two-dimensional Curve Fitting. J. Magn. Reson. 2002, 157, 141–155. [Google Scholar] [CrossRef]
- AOCS Official Method Cd 16b-93: SFC by Low Resolution Magnetic Resonance; American Oil Chemist Society: Urbana, IL, USA, 1993.
- 2.150 (ex 2.323): Solid Content Determination in Fats by NMR (Low Resolution Nuclear Magnetic Resonance); IUPAC Norm Version; IUPAC: Research Triangle Park, NC, USA, 1987.
- ISO 8292; 1991(E)/International Standard: Animal and Vegetable Fats and Oils—Determination of Solid Fat Content—Pulsed Nuclear Magnetic Resonance Method/First Edition 1991-12-01. ISO: Geneva, Switzerland, 1991.
- SMeiboom, S.; Gill, D. Modified spin echo method for measuring nuclear relaxation times. Rev. Scient. Instrum. 1958, 29, 688–691. [Google Scholar] [CrossRef]
- Provencher, S.W. Inverse problems in polymer characterization: Direct analysis of polydispersity with photon correlation spectroscopy. Makromol. Chem. 1979, 180, 201–211. [Google Scholar] [CrossRef]
- Provencher, S.W. CONTIN: A general purpose constrained regularization program for inverting noisy linear algebraic and integral equations. Comput. Phys. Commun. 1982, 27, 229–242. [Google Scholar] [CrossRef]
- MATLAB Mathworks; The Mathworks Inc.: Natick, MA, USA. Available online: https://it.mathworks.com/ (accessed on 1 February 2023).
- Marino, I.G. Rilt. MATLAB Central File Exchange. 2023. Available online: https://www.mathworks.com/matlabcentral/fileexchange/6523-rilt (accessed on 10 March 2023).
- Marquardt, D.W. An Algorithm for least-squares estimation for nonlinear parameters. J. Soc. Ind. Appl. Math. 1963, 11, 431–441. [Google Scholar] [CrossRef]
- SigmaPlot, version 9; Systat Software Inc.: San Jose, CA, USA, 2008.
- Nikiforidis, C.V. Structure and functions of oleosomes (oil bodies). Adv. Colloid Interface Sci. 2019, 274, 102039. [Google Scholar] [CrossRef]
- Gianferri, R.; D’Aiuto, V.; Curini, R.; Delfini, M.; Brosio, E. Proton NMR transverse relaxation measurements to study water dynamics states and age-related changes in Mozzarella di Bufala Campana cheese. Food Chem. 2007, 105, 720–726. [Google Scholar] [CrossRef]
- Ritota, M.; Gianferri, R.; Bucci, R.; Brosio, E. Proton NMR relaxation study of swelling and gelatinization process in rice starch-water samples. Food Chem. 2008, 110, 14–22. [Google Scholar] [CrossRef]
- Song, W.; Qin, Y.; Zhu, Y.; Yin, G.; Wu, N.; Li, Y.; Hu, Y. Delineation of plant caleosin residues critical for functional divergence, positive selection and coevolution. BMC Evol. Biol. 2014, 14, 124. [Google Scholar] [CrossRef]
- Yatsu, L.Y.; Jacks, T.J.; Hensorling, T.P. Isolation of spherosomes (oleasomes) from onion, cabbage, and cottonseed tissues. Plant Physiol. 1971, 48, 675–682. [Google Scholar] [CrossRef]
- Salisbury, F.B.; Ross, C.W. Lipidi e composti aromatici. In Fisiologia Vegetale; Zanichelli, Ed: Bologna, Italy, 1984; Volume 14, pp. 218–224. [Google Scholar]
- Hills, B.P.; Belton, P.S.; Quantin, V.M. Water proton relaxation in heterogeneous systems. I. Saturated randomly packed suspensions of impenetrable particles. Mol. Phys. 1993, 78, 893–908. [Google Scholar] [CrossRef]
- Traoré, A.; Foucat, L.; Renou, J.-P. 1H NMR studies: Dynamics of water in gelatin. Eur. Biophys. J. 2000, 29, 159–164. [Google Scholar]
- Holtz, M.; Heil, S.R. Sacco: Temperature-dependent self-diffusion coefficients of water and six selected molecular liquid for calibration in accurate 1H NMR PFG measurements. PCCP 2000, 2, 4740–4742. [Google Scholar] [CrossRef]
- Woessner, D.E. Brownian motion and its effects in NMR chemical exchange and relaxation in liquids. Concept Magn. Reson. 1996, 6, 397–421. [Google Scholar] [CrossRef]
- Palmer, A.G.; Koss, H. Chapter Six—Chemical Exchange; Joshua Wand, A., Ed.; Methods in Enzymology; Academic Press: Cambridge, MA, USA, 2019; Volume 615, pp. 177–236. ISSN 0076-6879/9780128167625. [Google Scholar] [CrossRef]
- Alberts, B.; Hopkin, K.; Johnson, A.; Morgan, D.; Raff, M. Essential Cell Biology; W. W. Norton & Company: New York, NY, USA, 2019; 750p, ISBN 9780393680393. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tonda Gentile Romana | ||||||||
| T (°C) | T2,a (ms) | Aa (%) | T2,b (ms) | Ab (%) | T2,c (ms) | Ac (%) | T2,d (ms) | Ad (%) |
| 15 | 186 ± 3 f | 37.7 ± 0.1 d | 68 ± 1 f | 49.1 ± 0.5 a | 22.4 ± 0.7 a | 7.5 ± 0.3 c | 0.25 ± 0.01 c | 5.7 ± 0.1 d |
| 30 | 234 ± 6 e | 49.0 ± 0.2 a | 86 ± 2 e | 41.2 ± 0.1 f | 22.0 ± 2.0 a | 2.6 ± 0.2 d | 0.34 ± 0.01 a | 7.2 ± 0.1 c |
| 35 | 264 ± 8 d | 43.3 ± 0.1 c | 97 ± 2 d | 46.2 ± 0.0 c | 21.0 ± 1.0 a | 2.9 ± 0.1 d | 0.30 ± 0.01 b | 7.6 ± 0.1 b |
| 40 | 297 ± 9 c | 47.5 ± 0.1 b | 109 ± 3 c | 42.8 ± 0.1 e | 18.0 ± 1.0 b | 1.7 ± 0.1 e | 0.35 ± 0.01 a | 7.9 ± 0.1 a |
| 45 | 460 ± 10 b | 33.8 ± 0.2 e | 166 ± 3 b | 47.7 ± 0.1 b | 21.7 ± 0.2 a | 11.4 ± 0.3 a | 0.20 ± 0.01 d | 7.1 ± 0.1 c |
| 55 | 510 ± 9 a | 37.4 ± 0.1 d | 187 ± 4 a | 45.2 ± 0.1 d | 19.7 ± 0.2 ab | 9.4 ± 0.2 b | 0.28 ± 0.01 b | 8.1 ± 0.0 a |
| Tonda di Giffoni | ||||||||
| T (°C) | T2,a (ms) | Aa (%) | T2,b (ms) | Ab (%) | T2,c (ms) | Ac (%) | T2,d (ms) | Ad (%) |
| 8 | 189 ± 4 e | 41.7 ± 0.0 a | 68 ± 2 e | 47.0 ± 0.5 c | 20.0 ± 2.0 b | 3.7 ± 0.4 f | 0.21 ± 0.01 ab | 7.7 ± 0.1 b |
| 17 | 270 ± 4 d | 25.1 ± 0.3 f | 98 ± 1 d | 48.0 ± 0.1 b | 23.8 ± 0.2 a | 22.0 ± 0.1 a | 0.14 ± 0.02 cd | 4.9 ± 0.5 d |
| 29 | 362 ± 9 c | 26.3 ± 0.2 e | 128 ± 2 c | 49.1 ± 0.4 a | 22.4 ± 0.2 a | 18.5 ± 0.4 b | 0.12 ± 0.02 d | 6.2 ± 0.6 c |
| 41 | 439 ± 8 b | 31.8 ± 0.3 d | 158 ± 3 b | 48.5 ± 0.3 ab | 22.0 ± 0.3 a | 13.2 ± 0.4 c | 0.17 ± 0.02 c | 6.5 ± 0.4 c |
| 45 | 448 ± 9 b | 34.8 ± 0.2 c | 162 ± 3 b | 46.4 ± 0.2 c | 20.8 ± 0.3 b | 10.9 ± 0.3 d | 0.19 ± 0.01 bc | 7.8 ± 0.2 b |
| 52 | 550 ± 10 a | 35.6 ± 0.3 b | 192 ± 4 a | 46.9 ± 0.1 c | 19.4 ± 0.3 b | 8.8 ± 0.2 e | 0.24 ± 0.01 a | 8.7 ± 0.0 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gianferri, R.; Sciubba, F.; Durazzo, A.; Gabrielli, P.; Lombardi-Boccia, G.; Giorgi, F.; Santini, A.; Engel, P.; Di Cocco, M.E.; Delfini, M.; et al. Time Domain NMR Approach in the Chemical and Physical Characterization of Hazelnuts (Corylus avellana L.). Foods 2023, 12, 1950. https://doi.org/10.3390/foods12101950
Gianferri R, Sciubba F, Durazzo A, Gabrielli P, Lombardi-Boccia G, Giorgi F, Santini A, Engel P, Di Cocco ME, Delfini M, et al. Time Domain NMR Approach in the Chemical and Physical Characterization of Hazelnuts (Corylus avellana L.). Foods. 2023; 12(10):1950. https://doi.org/10.3390/foods12101950
Chicago/Turabian StyleGianferri, Raffaella, Fabio Sciubba, Alessandra Durazzo, Paolo Gabrielli, Ginevra Lombardi-Boccia, Francesca Giorgi, Antonello Santini, Petra Engel, Maria Enrica Di Cocco, Maurizio Delfini, and et al. 2023. "Time Domain NMR Approach in the Chemical and Physical Characterization of Hazelnuts (Corylus avellana L.)" Foods 12, no. 10: 1950. https://doi.org/10.3390/foods12101950