
Production of Bacterial Cellulose in the Medium with Yeasts Pre-Fermented Coconut Water or with Addition of Selected Amino Acids
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Media
2.2. Fermentation
2.3. Determination of Amino Acids
2.4. Determination of Growth
2.5. Determination of Gluconic Acid Content
2.6. Analyses of Morphology, Fourier-Transform Infrared (FTIR) Spectrum, X-ray Diffraction (XRD), and Thermogravimetry (TG)
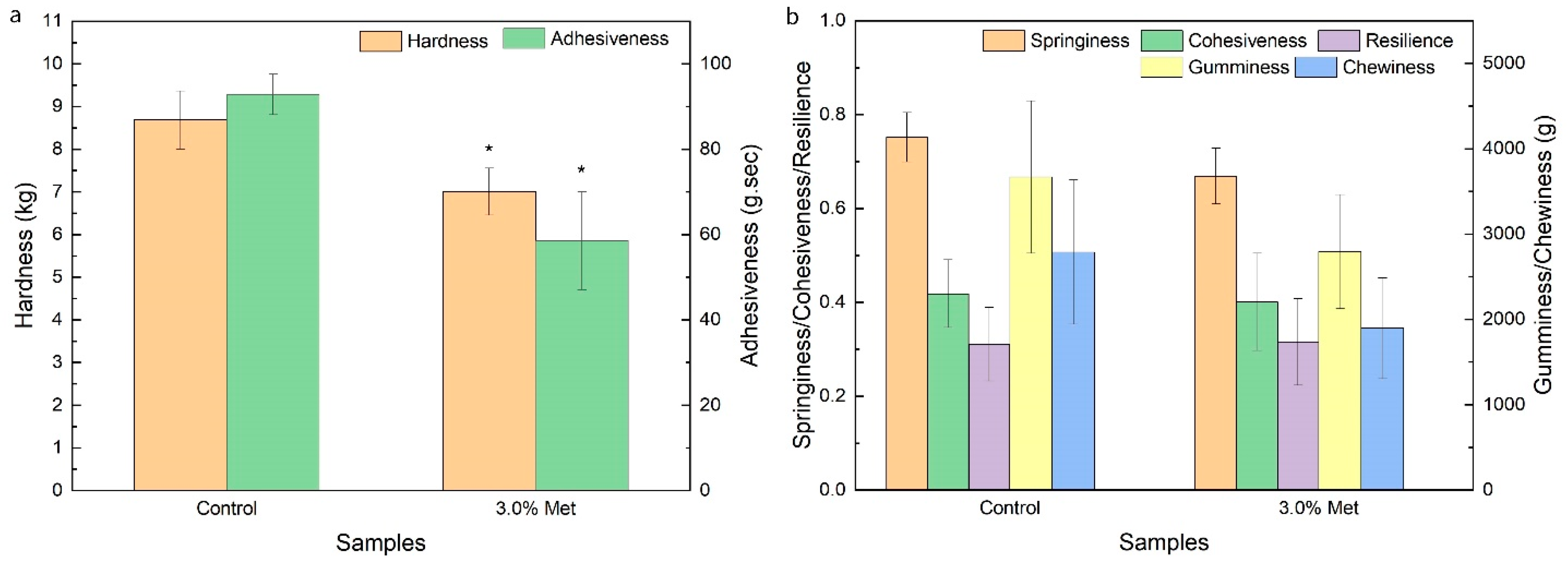
2.7. Analysis of Texture Characteristics
2.8. Statistical Analysis
3. Results
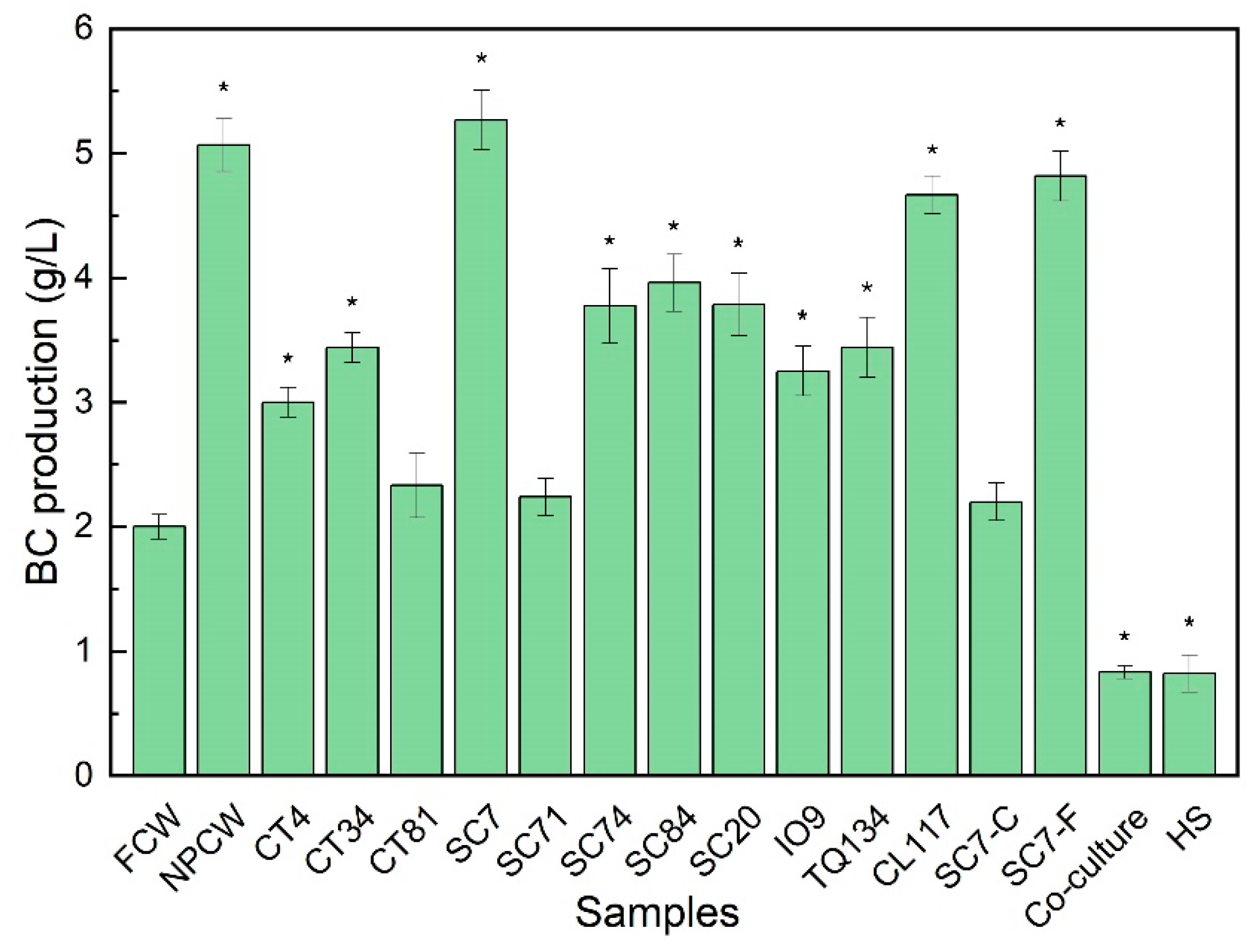
3.1. Production of BC from K. nataicola in the YPCW Media
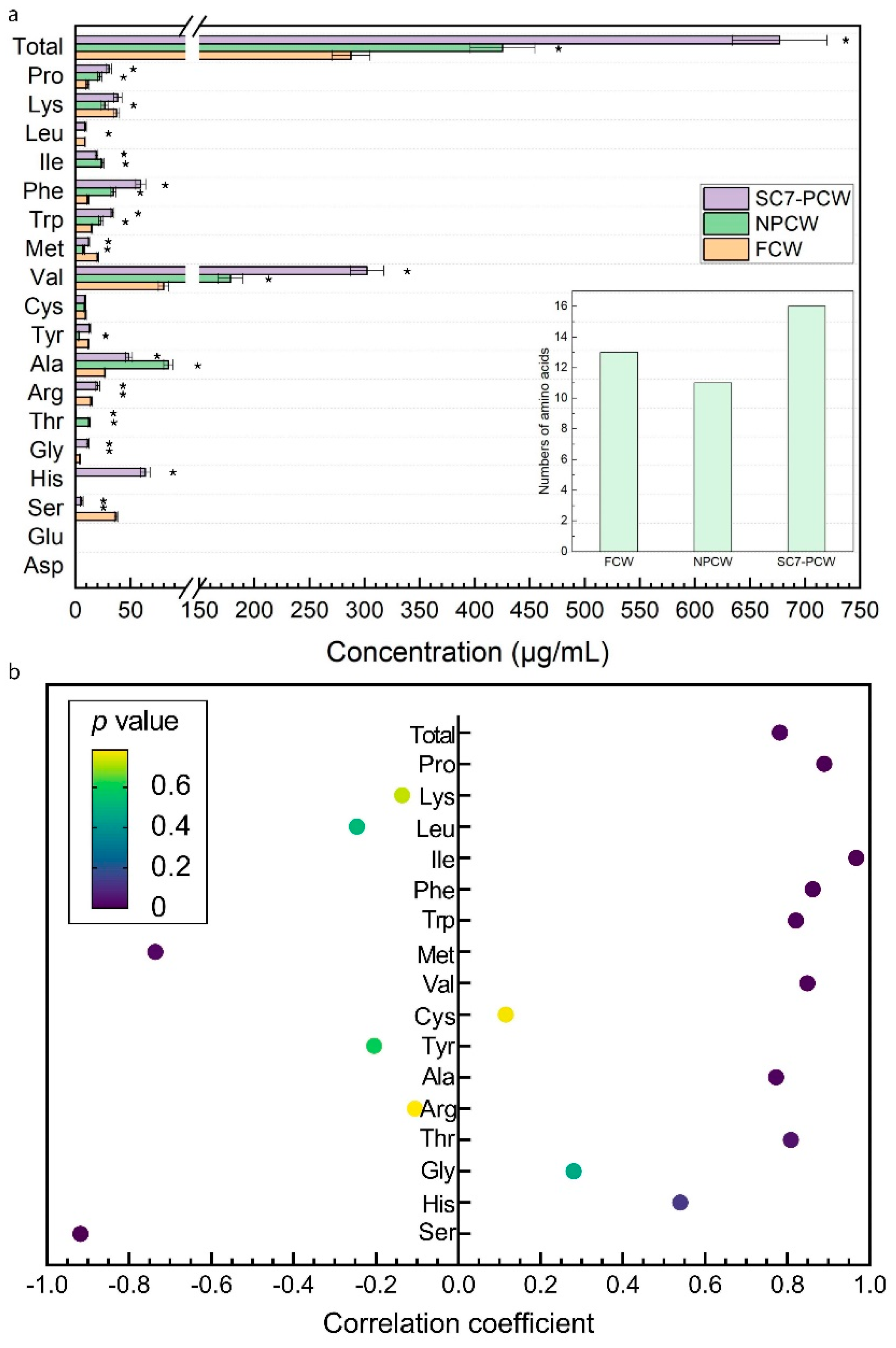
3.2. Amino Acids in FCW, NPCW, and SC7-PCW
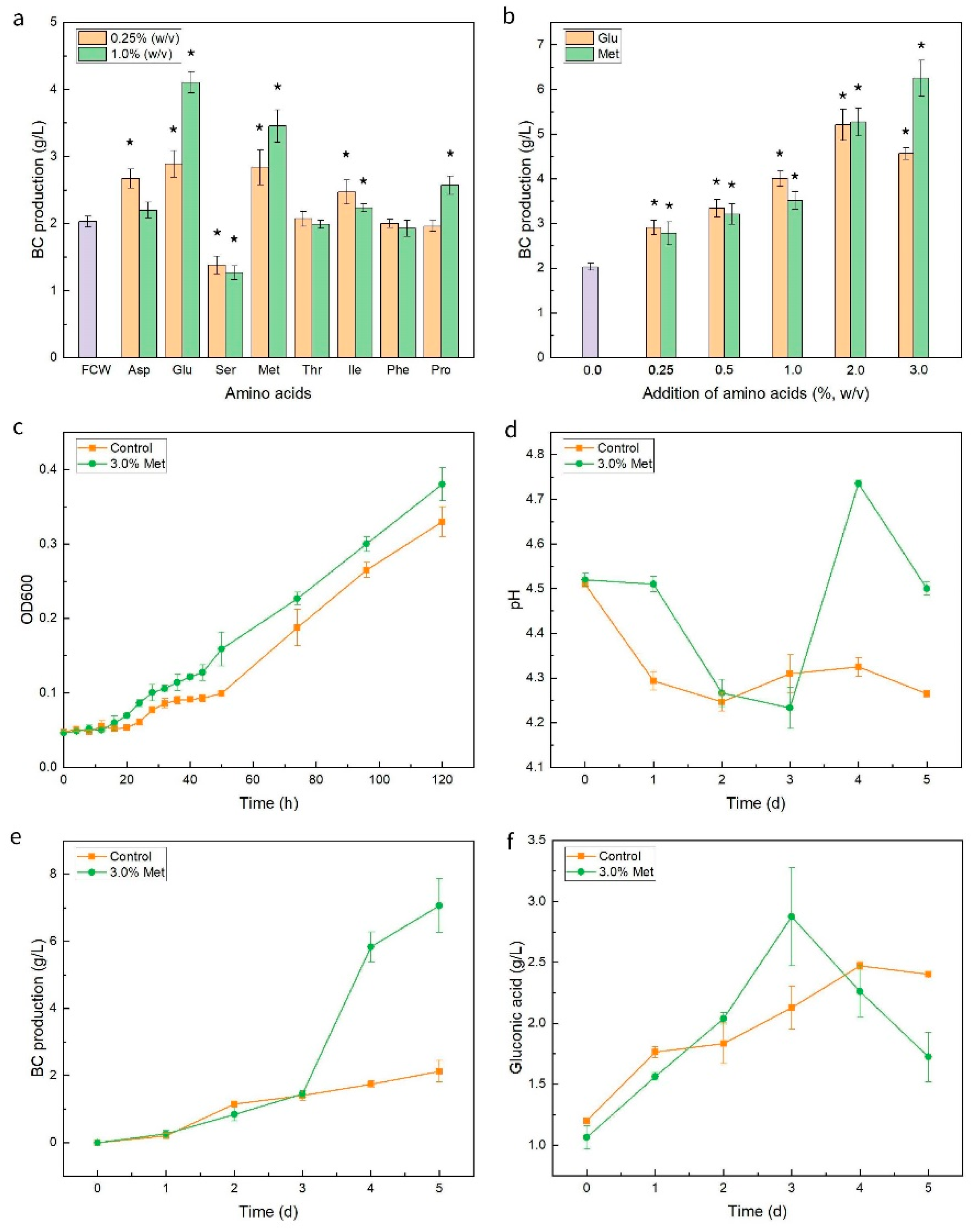
3.3. BC Production from K. nataicola with the Addition of Selected Amino Acids
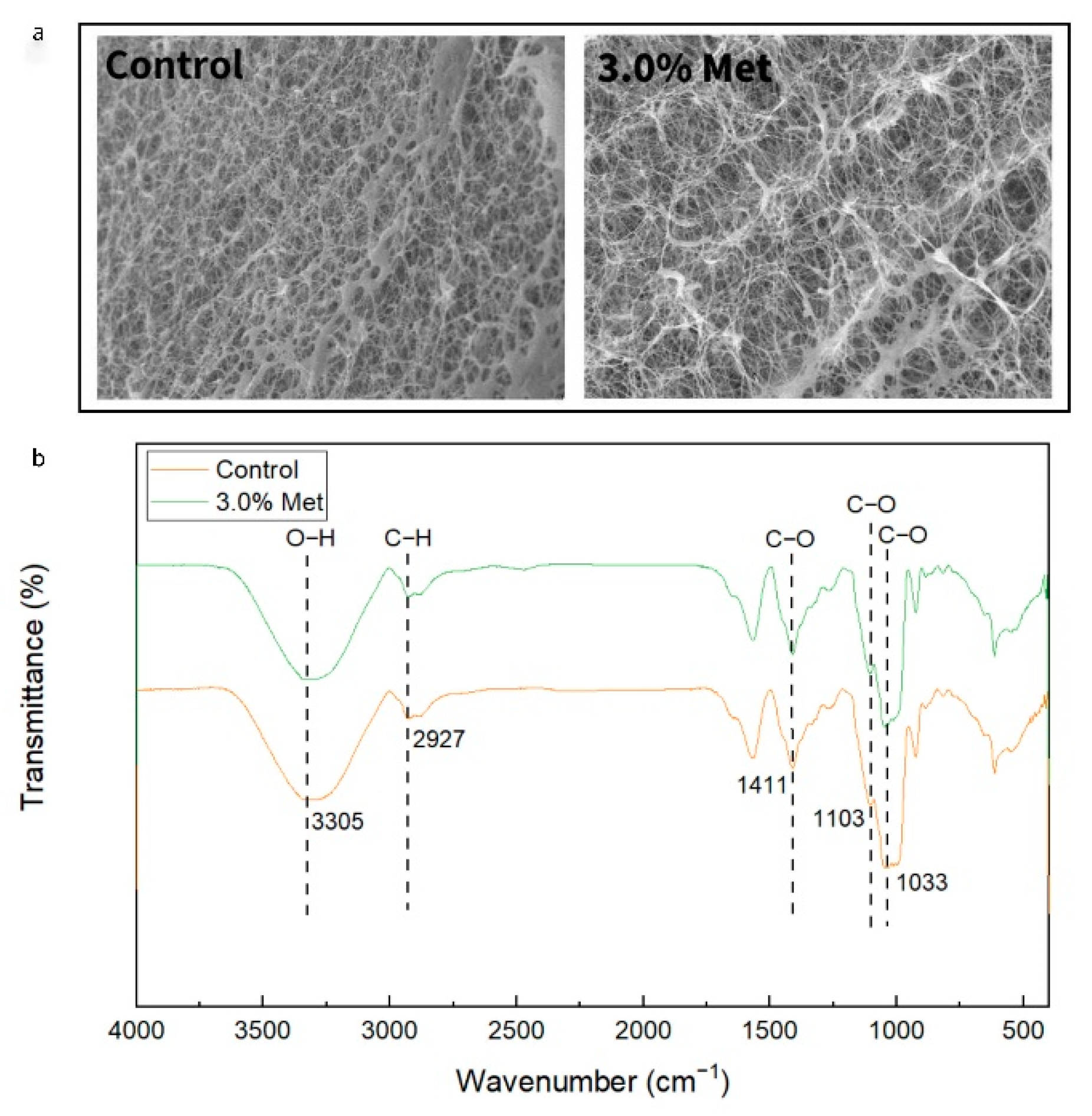
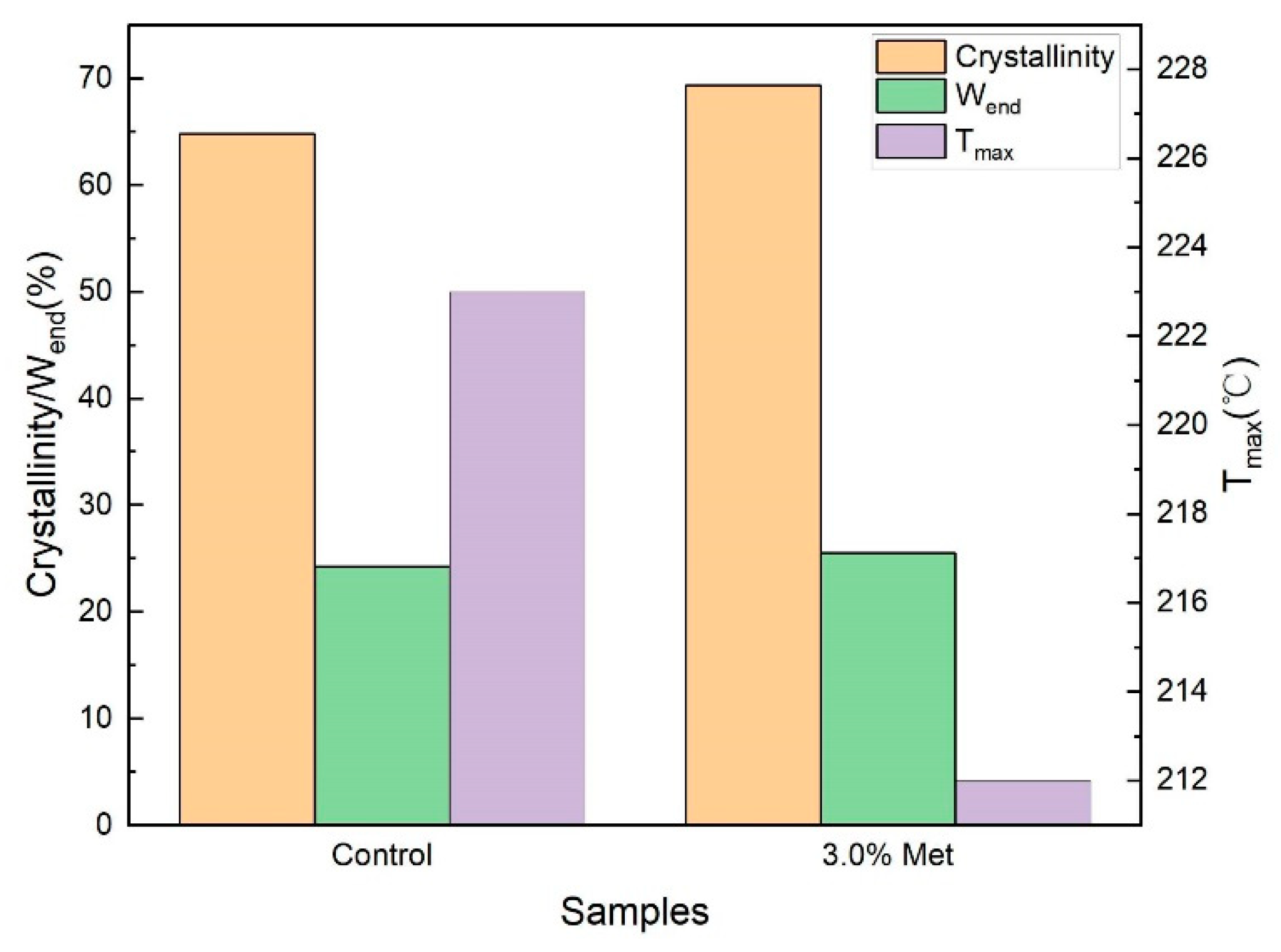
3.4. Characterization of BC Membranes Obtained with the Addition of 3.0% Methionine
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, J.; Yang, Y.; Deng, J.; Wang, Y.; Hu, Q.; Li, C.; Liu, S. Dynamic profile of the microbiota during coconut water pre-fermentation for nata de coco production. LWT-Food Sci. Technol. 2017, 81, 87–93. [Google Scholar] [CrossRef]
- Ul-Islam, M.; Ullah, M.W.; Khan, S.; Shah, N.; Park, J.K. Strategies for cost-effective and enhanced production of bacterial cellulose. Int. J. Biol. Macromol. 2017, 102, 1166–1173. [Google Scholar] [CrossRef] [PubMed]
- Ul-Islam, M.; Ullah, M.W.; Khan, S.; Park, J.K. Production of bacterial cellulose from alternative cheap and waste resources: A step for cost reduction with positive environmental aspects. Korean J. Chem. Eng. 2020, 37, 925–937. [Google Scholar] [CrossRef]
- Deng, J.; Liu, S.X.; Yang, Y.C.; Zhang, C.; He, X.L.; Bi, J.C.; Chen, H.M.; Li, C.F. Study on improvement of bacterial cellulose synthesis by coconut water pre-fermentation. Guangdong Agric. Sci. 2015, 19, 84–88. [Google Scholar]
- Wang, L.; Hong, H.; Zhang, C.; Huang, Z.; Guo, H. Transcriptome analysis of Komagataeibacter europaeus CGMCC 20445 responses to different acidity levels during acetic acid fermentation. Pol. J. Microbiol. 2021, 70, 305–313. [Google Scholar] [CrossRef]
- Yang, Y.C.; Li, C.F.; Deng, J.; Liu, L.X.; Hu, Q.S.; Bi, J.C. Changes of chemical composition and microbial community during naturally fermented coconut water used in nata de coco production. Food Res. Dev. 2015, 36, 168–171. [Google Scholar] [CrossRef]
- Maciel, N.O.P.; Piló, F.B.; Freitas, L.F.D.; Gomes, F.C.O.; Johann, S.; Nardi, R.M.D.; Lachance, M.; Rosa, C.A. The diversity and antifungal susceptibility of the yeasts isolated from coconut water and reconstituted fruit juices in Brazil. Int. J. Food Microbiol. 2013, 160, 201–205. [Google Scholar] [CrossRef]
- Amarasekara, A.S.; Wang, D.; Grady, T.L. A comparison of kombucha SCOBY bacterial cellulose purification methods. SN Appl. Sci. 2020, 2, 240. [Google Scholar] [CrossRef] [Green Version]
- Basak, A.K. Drying Characteristics of bacterial cellulose produced from fermentation of black tea by symbiotic colony of yeast and bacteria. Int. J. Sci. Res. 2013, 4, 1144–1147. [Google Scholar]
- Jayabalan, R.; Malbaša, R.V.; Lončar, E.S.; Vitas, J.S.; Sathishkumar, M. A review on Kombucha tea—Microbiology, composition, fermentation, beneficial effects, toxicity, and tea fungus. Compr. Rev. Food Sci. Food Saf. 2014, 13, 538–550. [Google Scholar] [CrossRef]
- Villarreal-Soto, S.A.; Beaufort, S.; Bouajila, J.; Souchard, J.; Taillandier, P. Understanding Kombucha tea fermentation: A review. J. Food Sci. 2018, 83, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Ou, H.; Yang, W.; Li, F.; Jia, S. Production of bacterial cellulose and beverage simultaneously by multi-strains-mixed-fermentation. Sci. Technol. Food Ind. 2008, 7, 188–191. [Google Scholar] [CrossRef]
- Ko, Y.H.; Lee, H.J.; Oh, H.J. Co-culture of a cellulose-producing acetic acid bacterium and a beer yeast. Life Sci. J. 2016, 13, 6. [Google Scholar] [CrossRef]
- Grahovac, J.; Grahovac, M.; Dodić, J.; Bajić, B.; Balaž, J. Optimization of cultivation medium for enhanced production of antifungal metabolites by Streptomyces hygroscopicus. Crop Prot. 2014, 65, 143–152. [Google Scholar] [CrossRef]
- Singh, V.; Haque, S.; Niwas, R.; Srivastava, A.; Pasupuleti, M.; Tripathi, C. Strategies for fermentation medium optimization: An in-depth review. Front. Microbiol. 2017, 7, 2087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, R.C.; Vanatta, P.R.; Fresco, J.R. Metabolic regulation of aminoacyl-tRNA synthetase biosynthesis in bakers’ yeast. J. Biol. Chem. 1977, 252, 878–882. [Google Scholar] [CrossRef]
- Nurfarahin, A.H.; Mohamed, M.S.; Phang, L.Y. Culture medium development for microbial-derived surfactants production—An overview. Molecules 1977, 23, 1049. [Google Scholar] [CrossRef] [Green Version]
- Gomes, R.J.; Ida, E.I.; Spinosa, W.A. Nutritional supplementation with amino acids on bacterial cellulose production by Komagataeibacter intermedius: Effect analysis and application of response surface methodology. Appl. Biochem. Biotechnol. 2022, 194, 5017–5036. [Google Scholar] [CrossRef]
- Wang, L.; Yang, X.; Li, Z.; Lin, X.; Hu, X.; Liu, S.; Li, C. Sensory characteristics of two kinds of alcoholic beverages produced with spent coffee grounds extract based on electronic senses and HS-SPME-GC-MS analyses. Fermentation 2021, 7, 254. [Google Scholar] [CrossRef]
- Wang, L.; Yang, X.; Jiang, H.; Song, Z.; Lin, X.; Hu, X.; Li, C. Protein kinases Elm1 and Sak1 of Saccharomyces cerevisiae exerted different functions under high-glucose and heat shock stresses. Appl. Microbiol. Biotechnol. 2022, 106, 2029–2042. [Google Scholar] [CrossRef]
- Zhang, L.; Hong, Q.; Yu, C.; Wang, R.; Li, C.; Liu, S. Acetobacter sp. improves the undesirable odors of fermented noni (Morinda citrifolia L.) juice. Food Chem. 2023, 401, 134126. [Google Scholar] [CrossRef]
- Qiao, W.; Qiao, Y.; Gao, G.; Liao, Z.; Wu, Z.; Saris, P.E.J.; Xu, H.; Qiao, M. A novel co-cultivation strategy to generate low-crystallinity bacterial cellulose and increase nisin yields. Int. J. Biol. Macromol. 2022, 202, 338–396. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Chen, S.; Wang, R.; Li, C.; Wang, L. Fabrication, characterization and biological properties of pectin and/or chitosan-based films incorporated with noni (Morinda citrifolia) fruit extract. Food Hydrocoll. 2023, 134, 108025. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhu, Y.; Xia, C. The nutrient value of edible yeast. Food Sci. Technol. 2004, 10, 94–96. [Google Scholar] [CrossRef]
- Zhu, H.; Li, Y.; Wang, J.; Lin, W.; Chen, Y. Enhancement of bacterial cellulose production in Bacillus amyloliquefaciens. IOP Conf. Ser. Mater. Sci. Eng. 2019, 493, 012036. [Google Scholar] [CrossRef]
- Zhang, L.P.; Lu, H.M.; Dai, R.; Dai, L.X.; Jiang, X.L. Study on the function of ethanol and organic acid to Acetobacter xylinum synthetic bacterial cellulose. Sci. Technol. Food Ind. 2014, 35, 161–169. [Google Scholar] [CrossRef]
- Hyun, J.Y.; Mahanty, B.; Kim, C.G. Utilization of makgeolli sludge filtrate (MSF) as low-cost substrate for bacterial cellulose production by Gluconacetobacter xylinus. Appl. Biochem. Biotech. 2014, 172, 3748–3760. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Chen, H.; Wang, Y.; Shao, L.; Liu, S.; Li, C.; Liu, S. The effect of ethanol on bacterial cellulose synthesis by different bacterial species and the sensitivity and tolerance of high-yield bacterial strains to ethanol. J. Trop. Biol. 2018, 9, 198–202. [Google Scholar] [CrossRef]
- Cheng, Y.F.; Du, Z.L.; Zhu, H.; Guo, X.; He, X. Protective effects of arginine on Saccharomyces cerevisiae against ethanol stress. Sci. Rep. 2016, 6, 31311. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Jiang, M.; Liu, Z.; Chen, Q.; Pei, F. Change of organic acid and free amino acid during Saccharomyces cerevisiae QY-1 fermentation. China Brew. 2019, 38, 125–128. [Google Scholar] [CrossRef]
- Xie, T.; Fang, H.Y.; Zhu, G.J. Kinetic analysis of stimulatory effects of amino acids on glycerol formation. Chin. J. Process Eng. 2006, 6, 623. [Google Scholar] [CrossRef] [Green Version]
- Wen, S.; Zhang, T.; Tan, T. Utilization of amino acids to enhance glutathione production in Saccharomyces cerevisiae. Enzym. Microb. Technol. 2004, 35, 501–507. [Google Scholar] [CrossRef]
- Son, H.J.; Kim, H.G.; Kim, K.K.; Kim, H.S.; Kim, Y.G.; Lee, S.J. Increased production of bacterial cellulose by Acetobacter sp. V6 in synthetic media under shaking culture conditions. Bioresour. Technol. 2003, 86, 215–219. [Google Scholar] [CrossRef]
- Heo, M.S.; Son, H.J. Development of an optimized, simple chemically defined medium for bacterial cellulose production by Acetobacter sp. A9 in shaking cultures. Biotechnol. Appl. Biochem. 2002, 36, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Parveen, N.; Cornell, K.A. Methylthioadenosine/S-adenosylhomocysteine nucleosidase, a critical enzyme for bacterial metabolism. Mol. Microbiol. 2011, 79, 7–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuoka, M.; Tsuchida, T.; Matsushita, K.; Adachi, O.; Yoshinaga, F. A synthetic medium for bacterial cellulose production by Acetobacter xylinum subsp. sucrofermentans. Biosci. Biotechnol. Biochem. 1996, 60, 575–579. [Google Scholar] [CrossRef]
- Seto, A.; Saito, Y.; Matsushige, M.; Kobayashi, H.; Sasaki, Y.; Tonouchi, N.; Tsuchida, T.; Yoshinaga, F.; Ueda, K.; Beppu, T. Effective cellulose production by a coculture of Gluconacetobacter xylinus and Lactobacillus mali. Appl. Microbiol. Biotechnol. 2006, 73, 915–921. [Google Scholar] [CrossRef]
- Pierucci, S.; Kleme, J.J.; Galdino, C.J.; D’Lamare, A.; de Amorim, J.D.P.; Nascimento, H.A.; Henrique, M.; Costa, A.F.S.; Sarubbo, L. The future of sustainable fashion: Bacterial cellulose biotextile naturally dyed. Chem. Eng. Trans. 2021, 86, 1333–1338. [Google Scholar] [CrossRef]
- Shrivastav, P.; Pramanik, S.; Vaidya, G.; Abdelgawad, M.A.; Ghoneim, M.M.; Singh, A.; Abualsoud, B.M.; Amaral, L.S.; Abourehab, M.A. Bacterial cellulose as a potential biopolymer in biomedical applications: A state-of-the-art review. J. Mater. Chem. B 2022, 10, 3199–3241. [Google Scholar] [CrossRef]
- Wahid, F.; Huang, L.H.; Zhao, X.Q.; Li, W.C.; Wang, Y.Y.; Jia, S.R.; Zhong, C. Bacterial cellulose and its potential for biomedical applications. Biotechnol. Adv. 2021, 53, 107856. [Google Scholar] [CrossRef]
- Yuan, J.X.; Wang, T.; Huang, X.N.; Wei, W. Research progress on the application of bacterial cellulose on papermaking. Pap. Pap. Mak. 2016, 35, 42–46. [Google Scholar] [CrossRef]
- Pillai, M.M.; Tran, H.N.; Sathishkumar, G.; Manimekalai, K.; Yoon, J.; Lim, D.Y.; Noh, I.; Bhattacharyya, A. Symbiotic culture of nanocellulose pellicle: A potential matrix for 3D bioprinting. Mater. Sci. Eng. C 2021, 119, 111552. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.C.; Ullah, N.; Wang, X.J.; Sun, X.C.; Li, C.Y.; Bai, Y.; Chen, L.; Li, Z.X. Characterization of bacterial cellulose by Gluconacetobacter hansenii CGMCC 3917. J. Food Sci. 2015, 80, 2217. [Google Scholar] [CrossRef]
- Lin, D.; Liu, Z.; Shen, R.; Chen, S.; Yang, X. Bacterial cellulose in food industry: Current research and future prospects. Int. J. Biol. Macromol. 2020, 158, 1007–1019. [Google Scholar] [CrossRef] [PubMed]
- Jacek, P.; Silva, F.; Dourado, F.; Bielecki, S.A.; Gama, M. Optimization and characterization of bacterial nanocellulose produced by Komagataeibacter rhaeticus K3. Carbohydr. Polym. Technol. Appl. 2020, 2, 100022. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, X.; Song, Z.; Jiang, H.; Hao, Y.; Hu, X.; Liu, S.; Li, C. Production of Bacterial Cellulose in the Medium with Yeasts Pre-Fermented Coconut Water or with Addition of Selected Amino Acids. Foods 2022, 11, 3627. https://doi.org/10.3390/foods11223627
Lin X, Song Z, Jiang H, Hao Y, Hu X, Liu S, Li C. Production of Bacterial Cellulose in the Medium with Yeasts Pre-Fermented Coconut Water or with Addition of Selected Amino Acids. Foods. 2022; 11(22):3627. https://doi.org/10.3390/foods11223627
Chicago/Turabian StyleLin, Xue, Zeming Song, Huanyuan Jiang, Yaofei Hao, Xiaoping Hu, Sixin Liu, and Congfa Li. 2022. "Production of Bacterial Cellulose in the Medium with Yeasts Pre-Fermented Coconut Water or with Addition of Selected Amino Acids" Foods 11, no. 22: 3627. https://doi.org/10.3390/foods11223627